

Functional Characterization of Saccharomyces cerevisiae P5C Reductase, the Enzyme at the Converging Point of Proline and Arginine Metabolism


Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrates and Reagents
2.2. Purification of Yeast P5C Reductase
2.3. P5C Reductase Assay
2.4. In Silico Analyses
3. Results
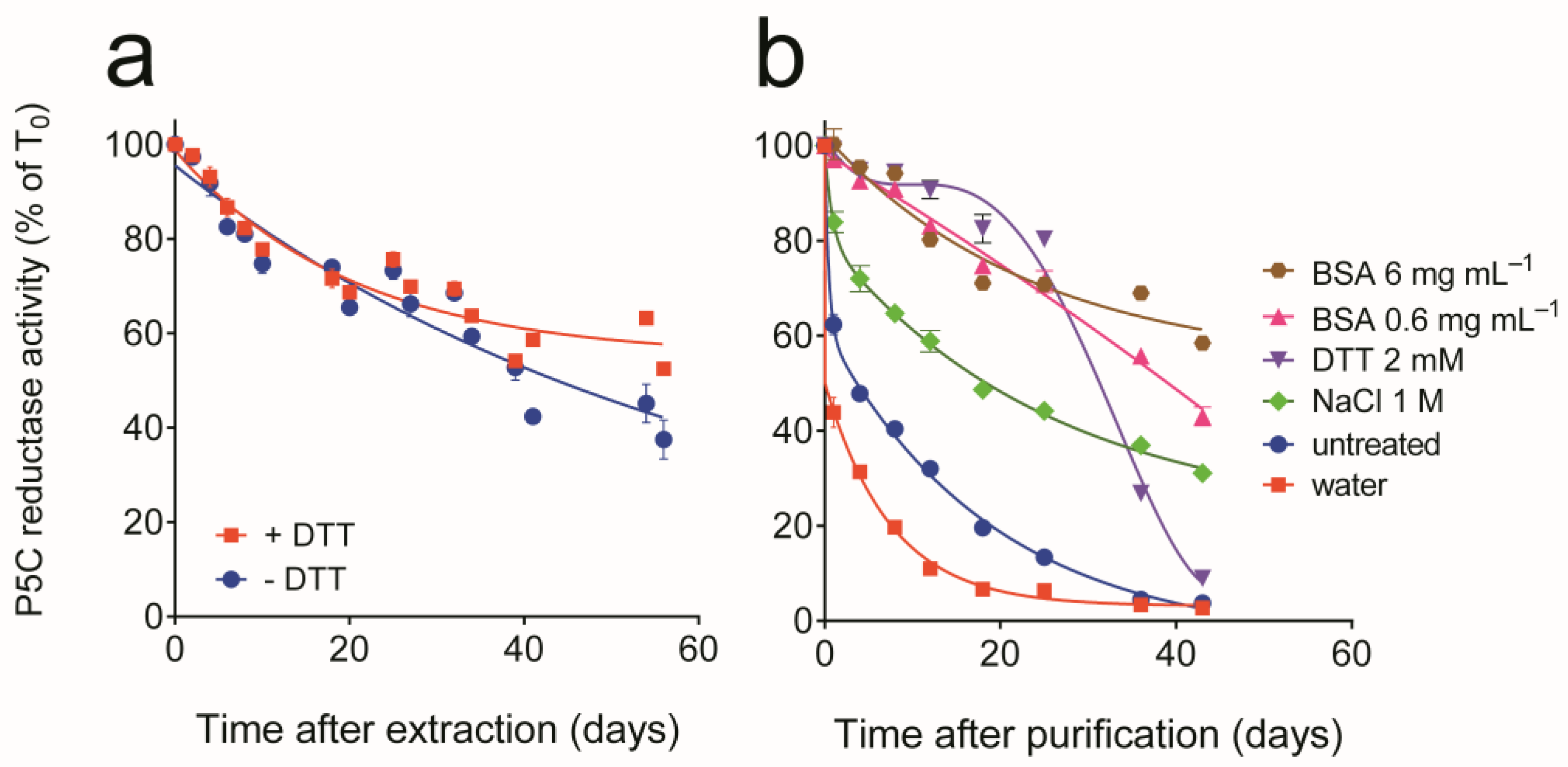
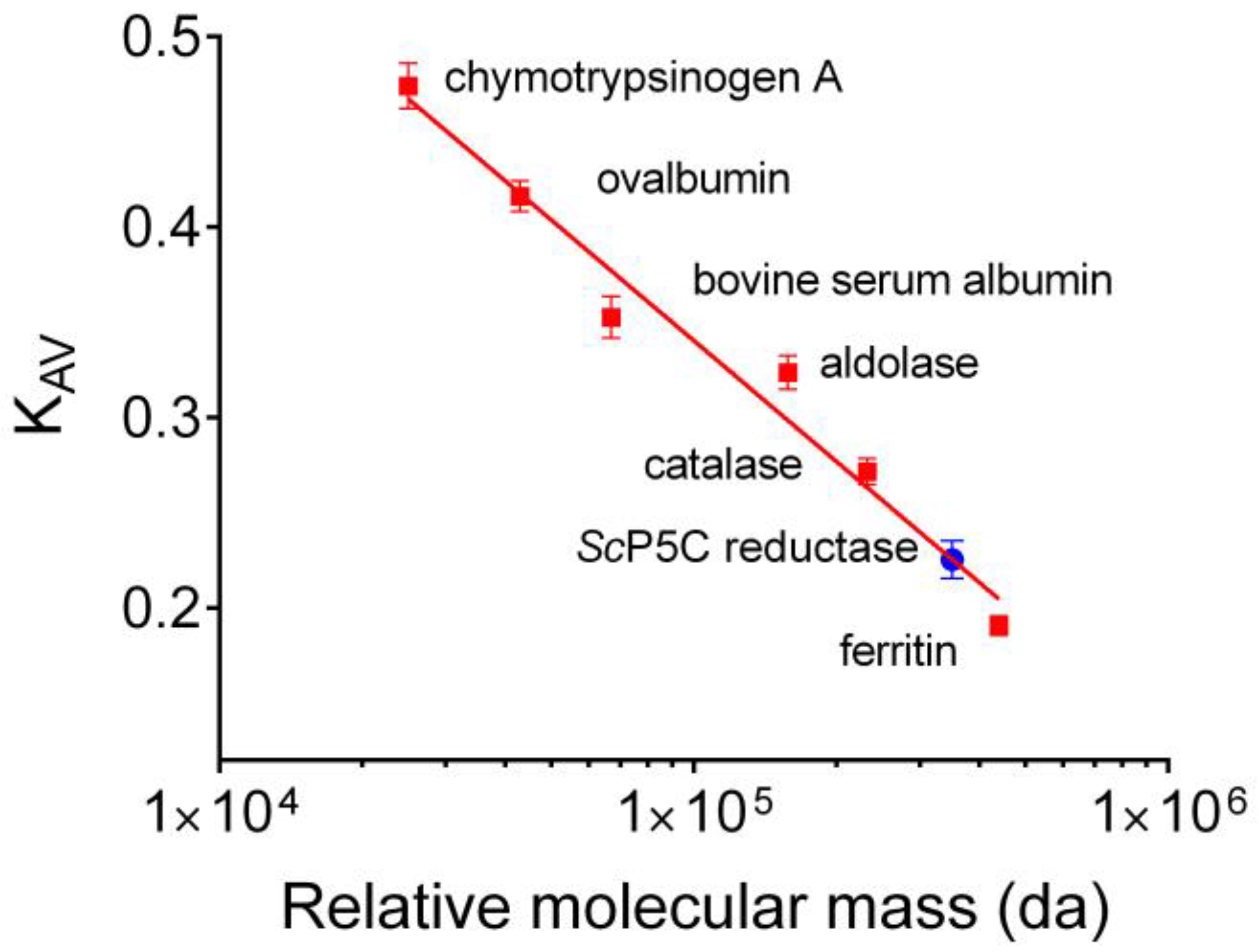
3.1. Purification of Yeast P5C Reductase
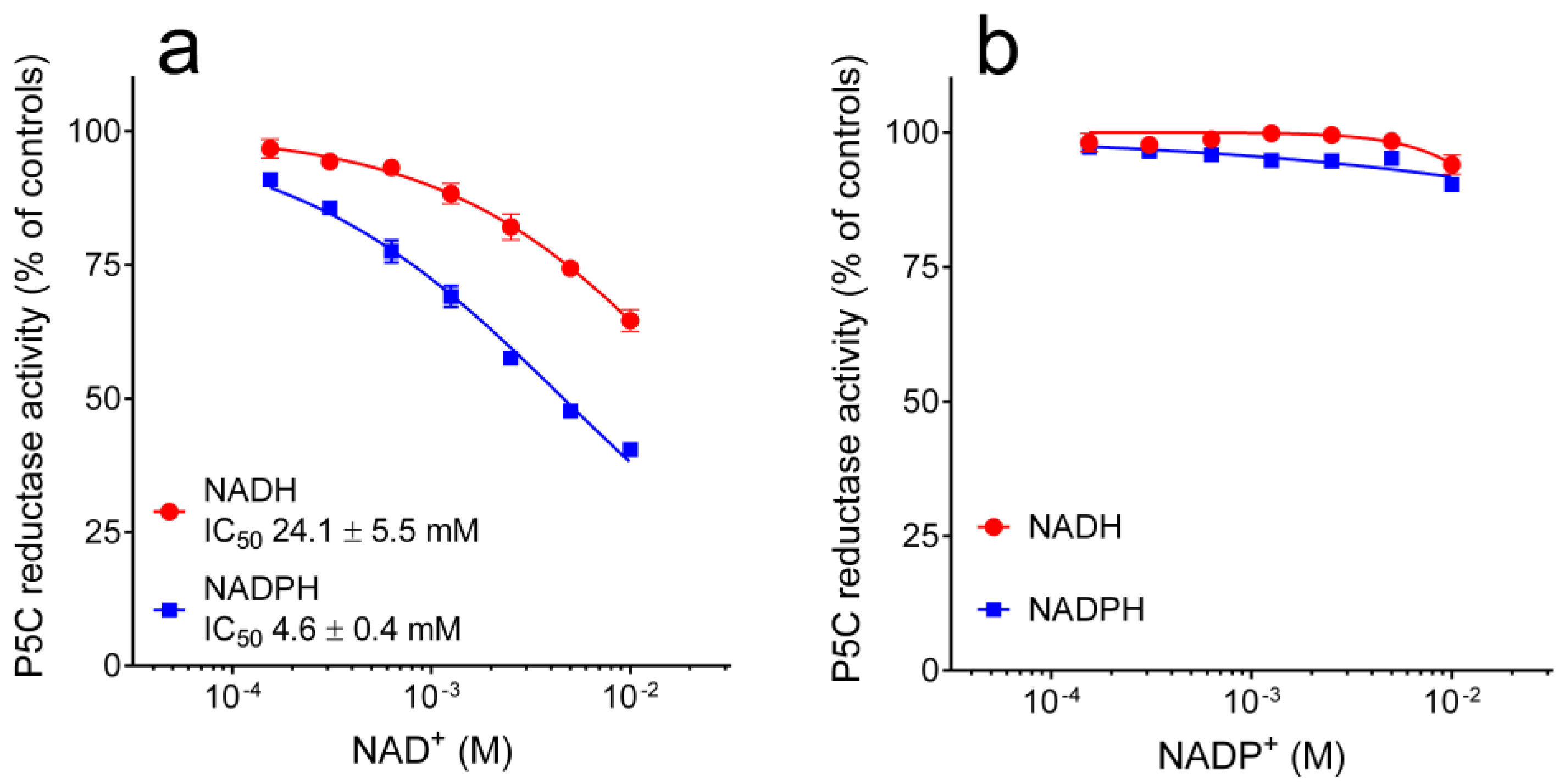
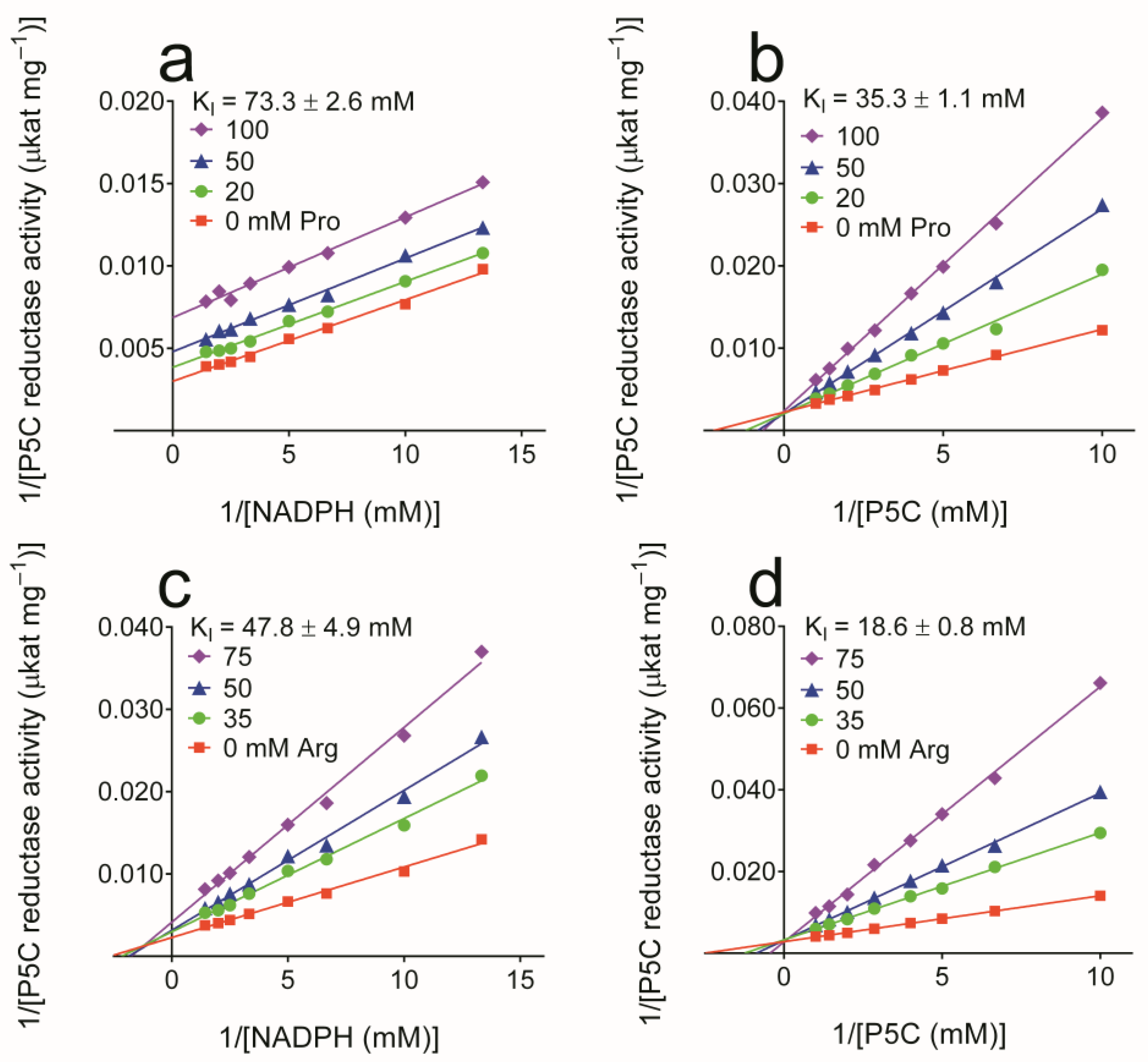
3.2. Properties of Yeast P5C Reductase
3.3. Structural Features of Yeast P5C Reductase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trovato, M.; Forlani, G.; Signorelli, S.; Funck, D. Proline Metabolism and Its Functions in Development and Stress Tolerance. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants: Recent Advances and Future Perspectives; Hossain, M.A., Kumar, V., Burritt, D.J., Fujita, M., Mäkelä, P.S.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 41–72. [Google Scholar]
- Forlani, G.; Trovato, M.; Funck, D.; Signorelli, S. Regulation of Proline Accumulation and Its Molecular and Physiological Functions in Stress Defence. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants: Recent Advances and Future Perspectives; Hossain, M.A., Kumar, V., Burritt, D.J., Fujita, M., Mäkelä, P.S.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 73–97. [Google Scholar]
- Zheng, Y.; Cabassa-Hourton, C.; Planchais, S.; Lebreton, S.; Savouré, A. The proline cycle as an eukaryotic redox valve. J. Exp. Bot. 2021, 72, 6856–6866. [Google Scholar] [CrossRef] [PubMed]
- Qamar, A.; Mysore, K.S.; Senthil-Kumar, M. Role of proline and pyrroline-5-carboxylate metabolism in plant defense against invading pathogens. Front. Plant Sci. 2015, 6, 503. [Google Scholar] [CrossRef] [Green Version]
- Huynh, T.Y.L.; Zareba, I.; Baszanowska, W.; Lewoniewska, S.; Palka, J. Understanding the role of key amino acids in regulation of proline dehydrogenase/proline oxidase (prodh/pox)-dependent apoptosis/autophagy as an approach to targeted cancer therapy. Mol. Cell. Biochem. 2020, 466, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Zarattini, M.; Forlani, G. Toward Unveiling the Mechanisms for transcriptional regulation of proline biosynthesis in the plant cell response to biotic and abiotic stress conditions. Front. Plant Sci. 2017, 8, 927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fichman, Y.; Gerdes, S.Y.; Kovács, H.; Szabados, L.; Zilberstein, A.; Csonka, L.N. Evolution of proline biosynthesis: Enzymology, bioinformatics, genetics, and transcriptional regulation. Biol. Rev. 2015, 90, 1065–1099. [Google Scholar] [CrossRef]
- Giberti, S.; Funck, D.; Forlani, G. Delta1-Pyrroline-5-carboxylate reductase from Arabidopsis thaliana: Stimulation or inhibition by chloride ions and feedback regulation by proline depend on whether NADPH or NADH acts as co-substrate. New Phytol. 2014, 202, 911–919. [Google Scholar] [CrossRef]
- Sabbioni, G.; Funck, D.; Forlani, G. Enzymology and regulation of δ1-pyrroline-5-carboxylate synthetase 2 from rice. Front. Plant Sci. 2021, 12, 672702. [Google Scholar] [CrossRef]
- Takagi, H. Proline as a stress protectant in yeast: Physiological functions, metabolic regulations, and biotechnological applications. Appl. Microbiol. Biotechnol. 2008, 81, 211–223. [Google Scholar] [CrossRef]
- Tomenchok, D.M.; Brandriss, M.C. Gene-enzyme relationships in the proline biosynthetic pathway of Saccharomyces cerevisiae. J. Bacteriol. 1987, 169, 5364–5372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandriss, M.C.; Magasanik, B. Proline: An essential intermediate in arginine degradation in Saccharomyces cerevisiae. J. Bacteriol. 1980, 143, 1403–1410. [Google Scholar] [CrossRef]
- Ter Schure, E.G.; van Riel, N.A.; Verrips, C.T. The role of ammonia metabolism in nitrogen catabolite repression in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2000, 24, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Sekine, T.; Kawaguchi, A.; Hamano, Y.; Takagi, H. Desensitization of feedback inhibition of the Saccharomyces cerevisiae gamma-glutamyl kinase enhances proline accumulation and freezing tolerance. Appl. Environ. Microbiol. 2007, 73, 4011–4019. [Google Scholar] [CrossRef] [Green Version]
- Brandriss, M.C.; Magasanik, B. Genetics and physiology of proline utilization in Saccharomyces cerevisiae: Enzyme induction by proline. J. Bacteriol. 1979, 140, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.L.; Brandriss, M.C. The regulator of the yeast proline utilization pathway is differentially phosphorylated in response to the quality of the nitrogen source. Mol. Cell. Biol. 2000, 20, 892–899. [Google Scholar] [CrossRef] [Green Version]
- Hinnebusch, A.G. Transcriptional and translational regulation of gene expression in the general control of amino-acid biosynthesis in Saccharomyces cerevisiae. Prog. Nucleic Acid Res. Mol. Biol. 1990, 38, 195–240. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, T.; Ishiguro, I. Δ1-Pyrroline-5-carboxylate reductase from Baker’s yeast. Purification, properties and its application in the assays of L-δ1-pyrroline-5-carboxylate and L-ornithine in tissue. Biochim. Biophys. Acta BBA-Enzymol. 1980, 613, 318–323. [Google Scholar] [CrossRef]
- Matsuzawa, T.; Ishiguro, I. Δ1-pyrroline-5-carboxylate reductase from baker’s yeast: Further purification by affinity chromatography with 5′ AMP-Sepharose 4B. Biochim. Biophys. Acta BBA-Enzymol. 1980, 616, 381–383. [Google Scholar] [CrossRef]
- Forlani, G.; Bertazzini, M.; Zarattini, M.; Funck, D.; Ruszkowski, M.; Nocek, B. Functional properties and structural characterization of rice δ1-pyrroline-5-carboxylate reductase. Front. Plant Sci. 2015, 6, 565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forlani, G.; Nocek, B.; Chakravarthy, S.; Joachimiak, A. Functional characterization of four putative δ1-pyrroline-5-carboxylate reductases from Bacillus subtilis. Front. Microbiol. 2017, 8, 1442. [Google Scholar] [CrossRef] [Green Version]
- Petrollino, D.; Forlani, G. Coenzyme preference of Streptococcus pyogenes δ1-pyrroline-5-carboxylate reductase: Evidence supporting NADPH as the physiological electron donor. Amino Acids 2012, 43, 493–497. [Google Scholar] [CrossRef]
- Forlani, G.; Nocek, B.; Ruszkowski, M. Peculiar substrate specificity of δ1-pyrroline-5-carboxylate reductase in the obligately fermentative bacterium Zymomonas mobilis. Mol. Biol. Rep. 2021, 48, 6205–6211. [Google Scholar] [CrossRef] [PubMed]
- Forlani, G.; Makarova, K.S.; Ruszkowski, M.; Bertazzini, M.; Nocek, B. Evolution of plant δ1-pyrroline-5-carboxylate reductases from phylogenetic and structural perspectives. Front. Plant Sci. 2015, 6, 567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, I.; Frank, L. Improved chemical synthesis and enzymatic assay of δ1-pyrroline-5-carboxylic acid. Anal. Biochem. 1975, 64, 85–97. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Ashkenazy, H.; Abadi, S.; Martz, E.; Chay, O.; Mayrose, I.; Pupko, T.; Ben-Tal, N. ConSurf 2016: An improved methodology to estimate and visualize evolutionary conservation in macromolecules. Nucleic Acids Res. 2016, 44, W344–W350. [Google Scholar] [CrossRef] [Green Version]
- Suzek, B.E.; Wang, Y.Q.; Huang, H.Z.; McGarvey, P.B.; Wu, C.H.; The UniProt Consortium. UniRef clusters: A comprehensive and scalable alternative for improving sequence similarity searches. Bioinformatics 2015, 31, 926–932. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef] [Green Version]
- Dolinsky, T.J.; Nielsen, J.E.; McCammon, J.A.; Baker, N.A. PDB2PQR: An automated pipeline for the setup of Poisson-Boltzmann electrostatics calculations. Nucleic Acids Res. 2004, 32, W665–W667. [Google Scholar] [CrossRef]
- Ramirez-Gaona, M.; Marcu, A.; Pon, A.; Guo, A.C.; Sajed, T.; Wishart, N.A.; Karu, N.; Djoumbou Feunang, Y.; Arndt, D.; Wishart, D.S. YMDB 2.0: A significantly expanded version of the yeast metabolome database. Nucleic Acids Res. 2017, 45, D440–D445. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Ruszkowski, M.; Nocek, B.; Forlani, G.; Dauter, Z. The structure of Medicago truncatula δ1-pyrroline-5-carboxylate reductase provides new insights into regulation of proline biosynthesis in plants. Front. Plant Sci. 2015, 6, 869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezl, V.A.; Knox, W.E. Properties and analysis of a stable derivative of pyrroline-5-carboxylic acid for use in metabolic studies. Anal. Biochem. 1976, 74, 430–440. [Google Scholar] [CrossRef]
- Giberti, S.; Bertazzini, M.; Liboni, M.; Berlicki, L.; Kafarski, P.; Forlani, G. Phytotoxicity of aminobisphosphonates targeting both δ1-pyrroline-5-carboxylate reductase and glutamine synthetase. Pest Manag. Sci. 2017, 73, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Deuschle, K.; Funck, D.; Forlani, G.; Stransky, H.; Biehl, A.; Leister, D.; van der Graaff, E.; Kunze, R.; Frommer, W.B. The role of δ1-pyrroline-5-carboxylate dehydrogenase in proline degradation. Plant Cell 2004, 16, 3413–3425. [Google Scholar] [CrossRef] [Green Version]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological implications of arginine metabolism in plants. Front. Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef] [Green Version]
- Zulkifli, M.; Neff, J.K.; Timbalia, S.A.; Garza, N.M.; Chen, Y.; Watrous, J.D.; Murgia, M.; Trivedi, P.P.; Anderson, S.K.; Tomar, D.; et al. Yeast homologs of human MCUR1 regulate mitochondrial proline metabolism. Nat. Commun. 2020, 11, 4866. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Total Activity (nkat) | Protein (mg) | Specific Activity (nkat (mg protein)−1) | Purification (Fold) | Yield (%) |
|---|---|---|---|---|---|
| Crude extract | 3234.8 | 85.6 | 37.79 | 1.0 | 100.0 |
| 60–80% ammonium sulfate fractionation | 1169.1 | 16.71 | 69.96 | 1.9 | 36.1 |
| Anion-exchange chromatography | 787.4 | 0.0386 | 20,398 | 540 | 24.3 |
| Gel permeation chromatography | 505.3 | 0.0053 | 95,344 | 2523 | 15.6 |
| Specific activity (NADH) 1 | 95.9 ± 2.7 μkat (mg protein)−1 |
| Specific activity (NADPH) 1 | 239.8 ± 6.8 μkat (mg protein)−1 |
| pH optimum 2 | 6.61 to 7.34 |
| KM (app) for L-P5C (NADH) 3 | 64.3 ± 5.5 μM |
| KM (app) for L-P5C (NADPH) 3 | 440.5 ± 21.4 μM |
| KM (app) for NADH 3 | 84.0 ± 7.1 μM |
| KM (app) for NADPH 3 | 133.4 ± 7.3 μM |
| Vmax (NADH) 3 | 107.9 ± 3.0 μkat (mg protein)−1 |
| Vmax (P5C, with NADH as the co-substrate) 3 | 111.9 ± 2.2 μkat (mg protein)−1 |
| Vmax (NADPH) 3 | 294.2 ± 5.7 μkat (mg protein)−1 |
| Vmax (P5C, with NADPH as the co-substrate) 3 | 349.1 ± 8.1 μkat (mg protein)−1 |
| Kcat (NADH) per monomer 4 | 3314 s−1 |
| Kcat (NADPH) per monomer 4 | 10,515 s−1 |
| Kcat/KM (NADH) | 3.95 × 107 M−1 s−1 |
| Kcat/KM (NADPH) | 7.88 × 107 M−1 s−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forlani, G.; Sabbioni, G.; Ruszkowski, M. Functional Characterization of Saccharomyces cerevisiae P5C Reductase, the Enzyme at the Converging Point of Proline and Arginine Metabolism. Microorganisms 2022, 10, 2077. https://doi.org/10.3390/microorganisms10102077
Forlani G, Sabbioni G, Ruszkowski M. Functional Characterization of Saccharomyces cerevisiae P5C Reductase, the Enzyme at the Converging Point of Proline and Arginine Metabolism. Microorganisms. 2022; 10(10):2077. https://doi.org/10.3390/microorganisms10102077
Chicago/Turabian StyleForlani, Giuseppe, Giuseppe Sabbioni, and Milosz Ruszkowski. 2022. "Functional Characterization of Saccharomyces cerevisiae P5C Reductase, the Enzyme at the Converging Point of Proline and Arginine Metabolism" Microorganisms 10, no. 10: 2077. https://doi.org/10.3390/microorganisms10102077