
Evaluation of Safety and Probiotic Potential of Enterococcus faecalis MG5206 and Enterococcus faecium MG5232 Isolated from Kimchi, a Korean Fermented Cabbage


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Whole-Genome Sequencing Analysis (WGS)
2.3. In Vitro Safety Test of the Strains MG5206 and MG5232
2.3.1. Antibiotic Susceptibility
2.3.2. Hyaluronidase Activity
2.3.3. Gelatinase Activity
2.3.4. Hemolytic Activity
2.4. Acute Toxicity Study
2.5. In Vitro Characterization of the Strains MG5206 and MG5232
2.5.1. Autoaggregation
2.5.2. Bacterial Adhesion to Solvents
2.5.3. Survivability in Simulated Gastrointestinal Condition
2.5.4. Enzyme Production and Carbohydrate Fermentation
3. Results and Discussion
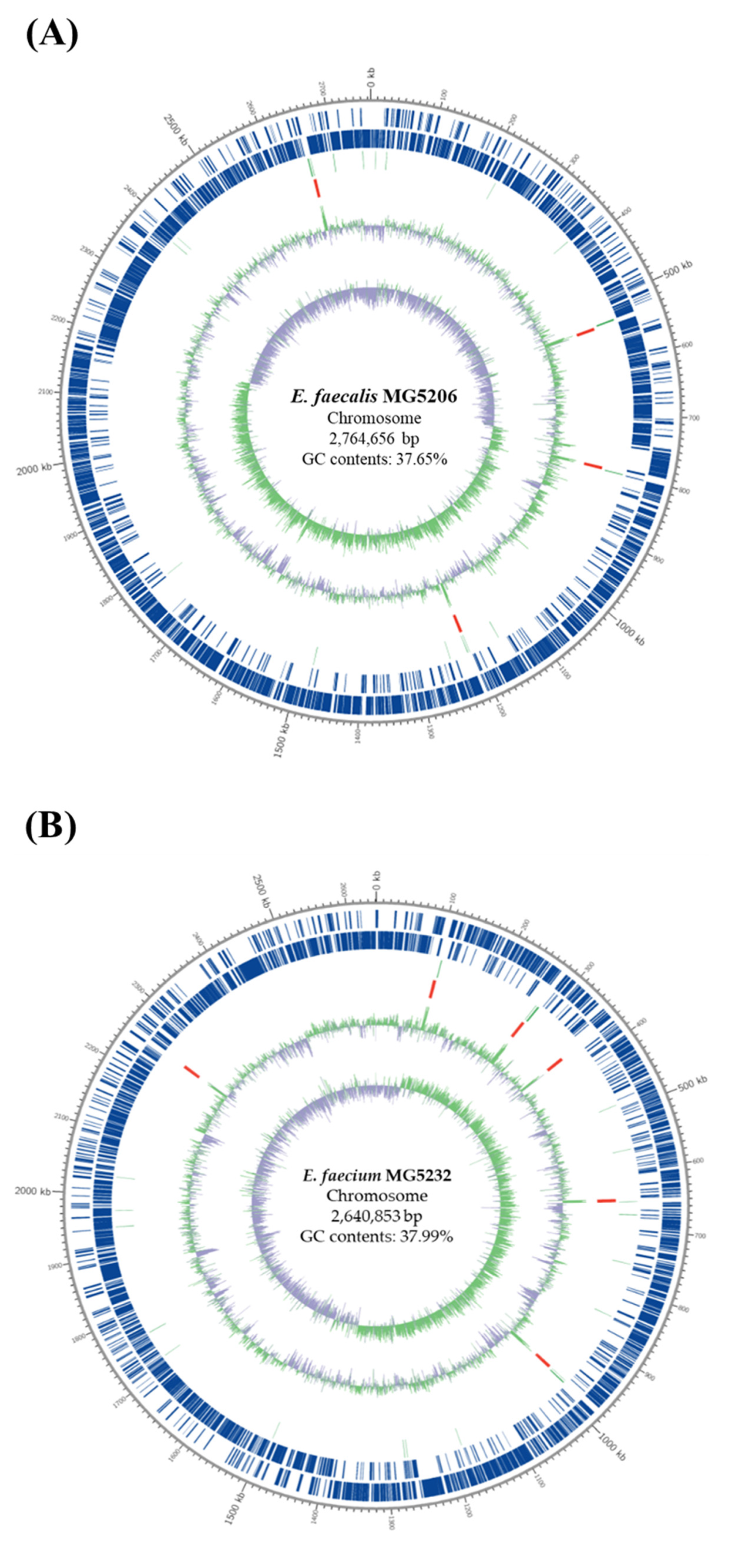
3.1. Whole-Genome Sequencing (WGS) Analysis of MG5206 and MG5232
3.2. Identification of Phenotypic Characteristics of the Strains MG5206 and MG5232
3.2.1. Antibiotic Resistance
3.2.2. Hyaluronidase and Gelatinase Activity
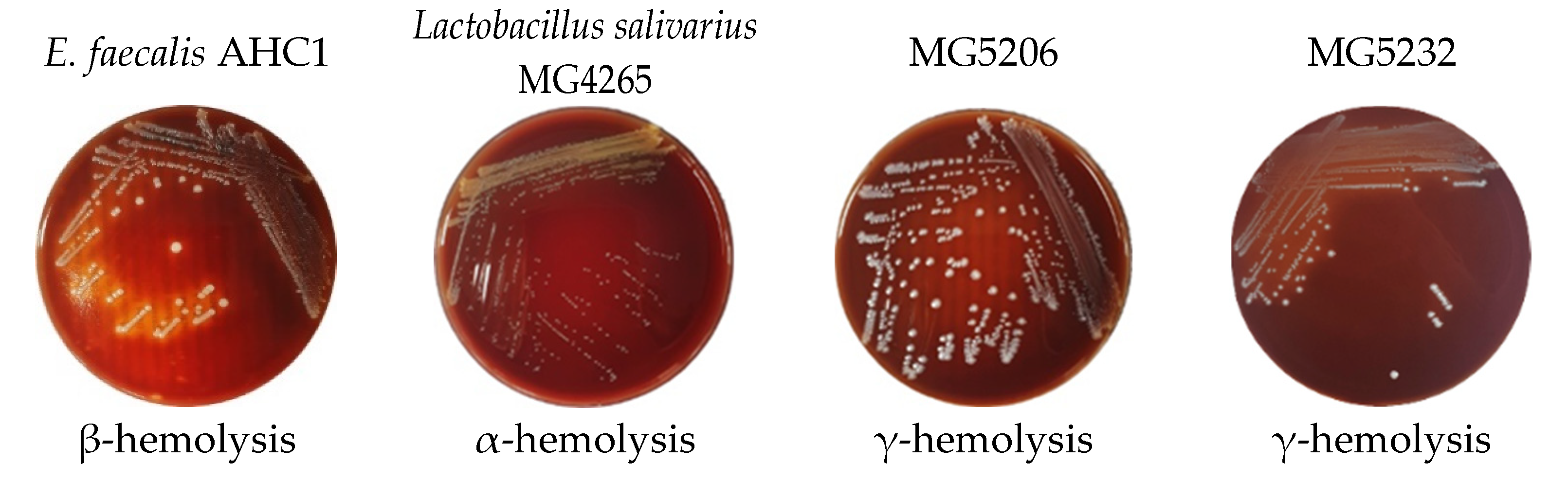
3.2.3. Hemolytic Activity
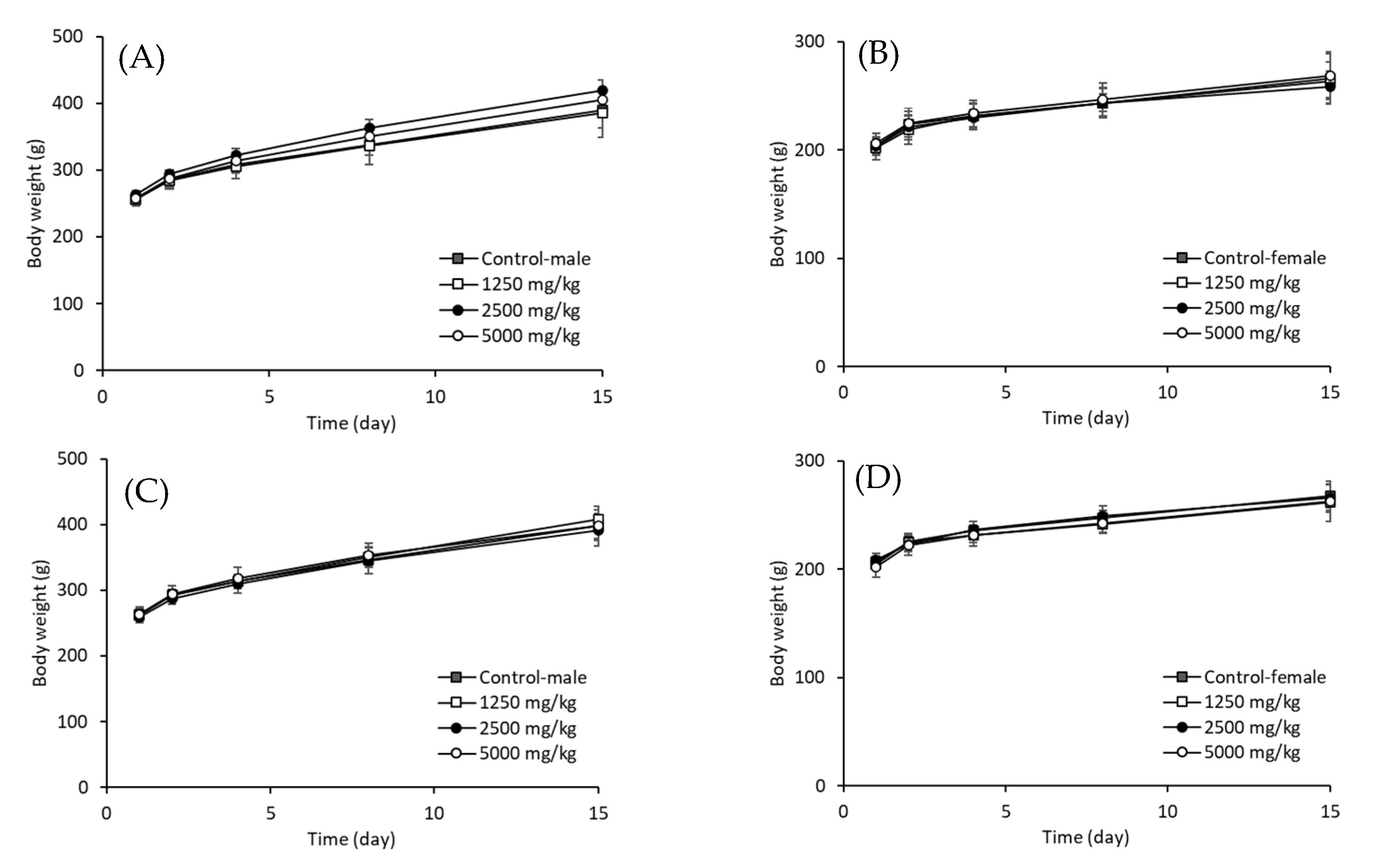
3.3. Acute Toxicity
3.4. Probiotic Properties of the Strains MG5206 and MG5232
3.4.1. Autoaggregation Ability
3.4.2. Hydrophobicity
3.4.3. Survival under Conditions Simulating the Human Gastrointestinal Tract
3.4.4. Characterization of Enzyme Production and Carbohydrate Fermentation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Klein, G. Taxonomy, ecology and antibiotic resistance of enterococci from food and the gastro-intestinal tract. Int. J. Food Microbiol. 2003, 88, 123–131. [Google Scholar] [CrossRef]
- Hugas, M.; Garriga, M.; Aymerich, M. Functionality of enterococci in meat products. Int. J. Food Microbiol. 2003, 88, 223–233. [Google Scholar] [CrossRef]
- Ben Omar, N.; Castro, A.; Lucas, R.; Abriouel, H.; Yousif, N.M.; Franz, C.M.; Holzapfel, W.H.; Rubén, P.-P.; Cañamero, M.M.M.; Galvez, A. Functional and Safety Aspects of Enterococci Isolated from Different Spanish Foods. Syst. Appl. Microbiol. 2004, 27, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Remschmidt, C.; Schröder, C.; Behnke, M.; Gastmeier, P.; Geffers, C.; Kramer, T.S. Continuous increase of vancomycin resistance in enterococci causing nosocomial infections in Germany- 10 years of surveillance. Antimicrob. Resist. Infect. Control. 2018, 7, 54. [Google Scholar] [CrossRef]
- Franz, C.M.; Stiles, M.E.; Schleifer, K.H.; Holzapfel, W.H. Enterococci in foods-a conundrum for food safety. Int. J. Food Microbiol. 2003, 88, 105–122. [Google Scholar] [CrossRef]
- Dogru, A.K.; Gencay, Y.E.; Ayaz, N.D. Comparison of virulence gene profiles of Enterococcus faecium and Enterococcus faecalis chicken neck skin and faeces isolates. Kafkas Univ. Vet. Fak. Derg. 2010, 16, 129–133. [Google Scholar]
- Eaton, T.J.; Gasson, M.J. Molecular Screening of Enterococcus Virulence Determinants and Potential for Genetic Exchange between Food and Medical Isolates. Appl. Environ. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef] [Green Version]
- Baccouri, O.; Boukerb, A.M.; Farhat, L.B.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I.; et al. Probiotic Potential and Safety Evaluation of Enterococcus faecalis OB14 and OB15, Isolated From Traditional Tunisian Testouri Cheese and Rigouta, Using Physiological and Genomic Analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef]
- Graham, K.; Stack, H.; Rea, R. Safety, beneficial and technological properties of enterococci for use in functional food applications—A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 3836–3861. [Google Scholar] [CrossRef]
- Son, S.-H.; Jeon, H.-L.; Jeon, E.B.; Lee, N.-K.; Park, Y.-S.; Kang, D.-K.; Paik, H.-D. Potential probiotic Lactobacillus plantarum Ln4 from kimchi: Evaluation of β-galactosidase and antioxidant activities. LWT Food Sci. Technol. 2017, 85, 181–186. [Google Scholar] [CrossRef]
- Lee, K.W.; Shim, J.M.; Park, S.K.; Heo, H.J.; Kim, H.J.; Ham, K.S.; Kim, J.H. Isolation of lactic acid bacteria with probiotic potentials from kimchi, traditional Korean fermented vegetable. LWT Food Sci. Technol. 2016, 71, 130–137. [Google Scholar] [CrossRef]
- Lee, S.-J.; Jeon, H.-S.; Yoo, J.-Y.; Kim, J.-H. Some Important Metabolites Produced by Lactic Acid Bacteria Originated from Kimchi. Foods 2021, 10, 2148. [Google Scholar] [CrossRef] [PubMed]
- Rho, M.K.; Kim, Y.E.; Rho, H.I.; Kim, T.R.; Kim, Y.B.; Sung, W.K.; Kim, T.W.; Kim, D.O.; Kang, H. Enterococcus faecium FC-K derived for kimchi is a probiotic strain that show anti-allergic activity. J. Microbiol. Biotech. 2017, 27, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Valledor, S.J.D.; Dioso, C.M.; Bucheli, J.E.V.; Park, Y.J.; Suh, D.H.; Jung, E.S.; Kim, B.; Holzapfel, W.H.; Todorov, S.D. Characterization and safety evaluation of two beneficial, enterocin-producing Enterococcus faecium strains isolated from kimchi, a Korean fermented cabbage. Food Microbiol. 2022, 102, 103886. [Google Scholar] [CrossRef]
- Ahn, S.B.; Park, H.E.; Lee, S.M.; Kim, S.Y.; Lee, W.K. Characteristics and immune-modulatory effect of Enterococcus faecium JS1-8 isolated from Kimchi. J. Biomed. Res. 2014, 15, 182–188. [Google Scholar] [CrossRef]
- Kang, C.-H.; Han, S.H.; Kim, Y.; Jeong, Y.; Paek, N.-S. Antibacterial Activity and Probiotic Properties of Lactic Acid Bacteria Isolated from Traditional Fermented Foods. KSBB J. 2017, 32, 199–205. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Cepas, J.; Szklarczyk, D.; Forslund, K.; Cook, H.; Heller, D.; Walter, M.C.; Rattei, T.; Mende, D.R.; Sunagawa, S.; Kuhn, M. eggNOG 4.5: A hierarchical orthology framework with improved functional annotations for eukaryotic, prokaryotic and viral sequences. Nucleic Acids Res. 2016, 44, D286–D293. [Google Scholar] [CrossRef] [Green Version]
- Klare, I.; Konstabel, C.; Müller-Bertling, S.; Reissbrodt, R.; Huys, G.; Vancanneyt, M.; Swings, J.; Goossens, H.; Witte, W. Evaluation of New Broth Media for Microdilution Antibiotic Susceptibility Testing of Lactobacilli, Pediococci, Lactococci, and Bifidobacteria. Appl. Environ. Microbiol. 2005, 71, 8982–8986. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Additives and Products; Substances Used in Animal Feed (FEEDAP); Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.d.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; et al. Guidance on the characterization of microorgan-isms used as feed additives or as production organisms. EFSA J. 2018, 16, e05206. [Google Scholar]
- Hynes, W.L.; Ferretti, J.J. Assays for hyaluronidase activity. Methods Enzymol. 1994, 235, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.; Gibbs, P.; Teixeira, P. Virulence factors among enterococci isolated from traditional fermented meat products produced in the North of Portugal. Food Control 2010, 21, 651–656. [Google Scholar] [CrossRef]
- Buxton, R. Blood Agar Plates and Hemolysis Protocols. Available online: https://asm.org/Protocols/Blood-Agar-Plates-and-Hemolysis-Protocols (accessed on 1 January 2016).
- The Organization for Economic Co-Operation and Development (OECD) Guidelines No. 420 for Acute Oral Toxicity-Fixed Dose Procedure. Available online: https://www.oecd-ilibrary.org/environment/test-no-420-acute-oral-toxicity-fixed-dose-procedure_9789264070943-en (accessed on 8 February 2002).
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Maragkoudakis, P.A.; Zoumpopoulou, G.; Miaris, C.; Kalantzopoulos, G.; Pot, B.; Tsakalidou, E. Probiotic potential of Lactobacillus strains isolated from dairy products. Int. Dairy J. 2006, 16, 189–199. [Google Scholar] [CrossRef]
- Jett, B.D.; Huyche, M.M.; Gilmore, M.S. Virulence of Enterococci. Clin. Microbiol. Rev. 1994, 7, 462–478. [Google Scholar] [CrossRef] [PubMed]
- Amini, F.; Krimpour, H.A.; Ghaderi, M.; Vaziri, S.; Ferdowsi, S.; Azizi, M.; Amini, S. Prevalence of aminoglycoside resistance genes in Enterococcus strains in Kermanshanh. Iran. J. Med. Sci. 2018, 43, 487–493. [Google Scholar]
- Fyfe, C.; Grossman, T.H.; Kerstein, K.; Sutcliffe, J. Resistance to Macrolide Antibiotics in Public Health Pathogens. Cold Spring Harb. Perspect. Med. 2016, 6, a025395. [Google Scholar] [CrossRef] [Green Version]
- Portillo, A.; Ruiz-Larrea, F.; Zarazaga, M.; Alonso, A.; Martinez, J.L.; Torres, C. Macrolide Resistance Genes in Enterococcus spp. Antimicrob. Agents Chemother. 2000, 44, 967–971. [Google Scholar] [CrossRef] [Green Version]
- Van Tyne, D.; Martin, M.J.; Gilmore, M.S. Structure, Function, and Biology of the Enterococcus faecalis Cytolysin. Toxins 2013, 5, 895–911. [Google Scholar] [CrossRef] [Green Version]
- Thurlow, L.R.; Thomas, V.C.; Narayanan, S.; Olson, S.; Fleming, S.D.L.; Hancock, L.E. Gelatinase contributes to the pathogenesis of endocarditis caused by Enterococcus faecalis. Infect. Immun. 2010, 78, 4936–4943. [Google Scholar] [CrossRef] [Green Version]
- Rosa, R.D.; Creti, R.; Venditti, M. Relationship between biofilm formation, the enterococcal surface protein (Esp) and gelatinase in clinical isolates of Enterococcus faecalis and Enterococcus faecium. FEMS Microbiol. Lett. 2006, 256, 145–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.; Munita, J.M.; A Arias, C. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti Infect. Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.A.; Contreras, G.A.; Murray, B.E. Management of multidrug-resistant enterococcal infections. Clin. Microbiol. Infect. 2010, 16, 555–562. [Google Scholar] [CrossRef] [PubMed]
- van Reenen, C.A.; Dicks, L.M. Horizontal gene transfer amongst probiotic lactic acid bacteria and other intestinal microbiota: What are the possibilities? A review. Arch. Microbiol. 2011, 193, 157–168. [Google Scholar] [CrossRef] [PubMed]
- WHO. Report on Joint FAO/WHO Guidelines for the Evaluation of Probiotics in Food. Available online: http://fanus.com.ar/posgrado/10-09-25/fao%20probiotics.pdf (accessed on 3 August 2022).
- Patil, S.; Chaudhari, B. A simple, rapid and sensitive plate assay for detection of microbial hyaluronidase activity. J. Basic Microbiol. 2017, 57, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Liu, Z.D.; Xue, Y.; Wang, J.F.; Li, H.; Tang, Q.J.; Wang, Y.M.; Dong, P.; Xue, C.H. Ds-echinoside A, A new triterpene glycoside derived from sea cucumber, exhibits antimetastatic activity via the inhibition of NF-κB-dependent MMP-9 and VEGF expressions. J. Zhejiang Univ. Sci. B 2011, 12, 534–544. [Google Scholar] [CrossRef] [Green Version]
- Patil, S.; Bhadane, B.; Shirsath, L.; Patil, R.; Chaudhari, B. Steroidal fraction of Carissa carandas L. inhibits microbial hyaluronidase activity by mixed inhibition mechanism. Prep. Biochem. Biotechnol. 2019, 49, 298–306. [Google Scholar] [CrossRef]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–569. [Google Scholar] [CrossRef] [Green Version]
- Sava, I.G.; Heikens, E.; Huebner, J. Pathogenesis and immunity in enterococcal infections. Clin. Microbiol. Infect. 2010, 16, 533–540. [Google Scholar] [CrossRef]
- Franz, C.M.; Huch, M.; Abriouel, H.; Holzapfel, W.; Gálvez, A. Enterococci as probiotics and their implications in food safety. Int. J. Food Microbiol. 2011, 151, 125–140. [Google Scholar] [CrossRef] [Green Version]
- Deng, F.; Chen, Y.; Zhou, X.; Xiao, H.; Sun, T.; Deng, Y.; Wen, J. New Insights into the Virulence Traits and Antibiotic Resistance of Enterococci Isolated from Diverse Probiotic Products. Microorganisms 2021, 9, 726. [Google Scholar] [CrossRef] [PubMed]
- Sanlibaba, P.; Tezel, B.U.; Senturk, E. Antimicrobial Resistance of Enterococcus Species Isolated from Chicken in Turkey. Korean J. Food Sci. Anim. Resour. 2018, 38, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Dubin, K.; Pamer, E.G. Enterococci and Their Interactions with the Intestinal Microbiome. Microbiol. Spectr. 2017, 5, 5–6. [Google Scholar] [CrossRef]
- Manap, N.A.; Azlan, A.; Hamzah, H.; Fakurazi, S.; Kadir, N. Evaluation of acute toxicity induced by supercritical carbon dioxide extract of Canarium odontophyllum (CO) Miq. pulp oil in SPF Sprague Dawley rats. Malays. J. Med. Health Sci. 2019, 15, 113–119. [Google Scholar]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef] [PubMed]
- Krausova, G.; Hyrslova, I.; Hynstova, I. In Vitro Evaluation of Adhesion Capacity, Hydrophobicity, and Auto-Aggregation of Newly Isolated Potential Probiotic Strains. Fermentation 2019, 5, 100. [Google Scholar] [CrossRef] [Green Version]
- Mansour, N.M.; Elkhatib, W.; Aboshanab, K.; Bahr, M. Inhibition of Clostridium difficile in Mice Using a Mixture of Potential Probiotic Strains Enterococcus faecalis NM815, E. faecalis NM915, and E. faecium NM1015: Novel Candidates to Control C. difficile Infection (CDI). Probiotics Antimicrob. Proteins 2018, 10, 511–522. [Google Scholar] [CrossRef]
- Rojas, M.; Ascencio, F.; Conway, P.L. Purification and Characterization of a Surface Protein from Lactobacillus fermentum 104R That Binds to Porcine Small Intestinal Mucus and Gastric Mucin. Appl. Environ. Microbiol. 2002, 68, 2330–2336. [Google Scholar] [CrossRef] [Green Version]
- Schär-Zammaretti, P.; Ubbink, J. The Cell Wall of Lactic Acid Bacteria: Surface Constituents and Macromolecular Conformations. Biophys. J. 2003, 85, 4076–4092. [Google Scholar] [CrossRef] [Green Version]
- Chou, L.-S.; Weimer, B. Isolation and Characterization of Acid- and Bile-Tolerant Isolates from Strains of Lactobacillus acidophilus. J. Dairy Sci. 1999, 82, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Succi, M.; Tremonte, P.; Reale, A.; Sorrentino, E.; Grazia, L.; Pacifico, S.; Coppola, R. Bile salt and acid tolerance of Lactobacillus rhamnosus strains isolated from Parmigiano Reggiano cheese. FEMS Microbiol. Lett. 2005, 244, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, H.-L.; Lee, N.-K.; Yang, S.-J.; Kim, W.-S.; Paik, H.-D. Probiotic characterization of Bacillus subtilis P223 isolated from kimchi. Food Sci. Biotechnol. 2017, 26, 1641–1648. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | MG5206 | MG5232 |
|---|---|---|
| No. of contigs | 1 | 2 |
| Chromosome size (bp) | 2,764,656 | 2,640,853 |
| Plasmid size (bp) | - | 136,433 |
| G + C contents (%) | 37.65 | 37.92 |
| Total genes | 2635 | 2707 |
| rRNA genes | 12 | 18 |
| tRNA genes | 61 | 68 |
| Coding sequence (CDS) | 2562 | 2621 |
| Total genome size (bp) | 2,764,656 | 2,777,286 |
| No. of plasmid | 0 | 1 |
| EggNOG | Function | MG5206 | MG5232 | ||
|---|---|---|---|---|---|
| No. of CDS | Ration of CDS | No. of CDS | Ration of CDS | ||
| J | Translation, ribosomal structure, and biogenesis | 157 | 6.150 | 148 | 5.732 |
| A | RNA processing and modification | 0 | 0.000 | 0 | 0.000 |
| K | Transcription | 191 | 7.481 | 190 | 7.358 |
| L | Replication, recombination, and repair | 123 | 4.818 | 193 | 7.475 |
| B | Chromatin structure and dynamics | 0 | 0 | 0 | 0.000 |
| D | Cell cycle control, cell division, and chromosome partitioning | 20 | 0.783 | 20 | 0.775 |
| Y | Nuclear structure | 0 | 0 | 0 | 0.000 |
| V | Defense mechanisms | 59 | 2.311 | 53 | 2.053 |
| T | Signal transduction mechanisms | 62 | 2.429 | 65 | 2.517 |
| M | Cell wall/membrane/envelope biogenesis | 121 | 4.740 | 133 | 5.151 |
| C | Cell motility | 5 | 0.196 | 4 | 0.155 |
| Z | Cytoskeleton | 0 | 0.000 | 0 | 0.000 |
| W | Extracellular structures | 0 | 0.000 | 0 | 0.000 |
| U | Intracellular trafficking, secretion, and vesicular transport | 23 | 0.901 | 27 | 1.046 |
| O | Posttranslational modification, protein turnover, and chaperones | 56 | 2.194 | 59 | 2.285 |
| C | Energy production and conversion | 95 | 3.721 | 73 | 2.827 |
| G | Carbohydrate transport and metabolism | 218 | 8.539 | 286 | 11.077 |
| E | Amino acid transport and metabolism | 173 | 6.776 | 146 | 5.655 |
| F | Nucleotide transport and metabolism | 89 | 3.486 | 73 | 2.827 |
| H | Coenzyme transport and metabolism | 57 | 2.233 | 40 | 1.549 |
| I | Lipid transport and metabolism | 52 | 2.037 | 49 | 1.898 |
| P | Inorganic ion transport and metabolism | 131 | 5.131 | 109 | 4.222 |
| Q | Secondary metabolites biosynthesis, transport, and catabolism | 17 | 0.666 | 15 | 0.581 |
| R | General function prediction only | 231 | 9.048 | 268 | 10.380 |
| S | Function unknown | 673 | 26.361 | 631 | 24.438 |
| Total | 2553 | 100 | 2,582 | 100 | |
| Antibiotics | MIC (µL/mL) | Cut-Off Value (µL/mL) | |
|---|---|---|---|
| MG5206 | MG5232 | ||
| Ampicillin | 2.0 ± 0.0 | 2.0 ± 0.0 | 2.0 |
| Chloramphenicol | 8.0 ± 0.0 | 8.0 ± 0.0 | 16.0 |
| Clindamycin | 4.0 ± 0.0 | 2.0 ± 0.0 | 4.0 |
| Erythromycin | 2.0 ± 0.0 | 4.0 ± 0.0 | 4.0 |
| Gentamycin | 32.0 ± 0.0 | 8.0 ± 0.0 | 32.0 |
| Kanamycin | 256.0 ± 0.0 | 1024.0 ± 0.0 | 1024.0 |
| Streptomycin | 128.0 ± 0.0 | 64.0 ± 0.0 | 128.0 |
| Tetracycline | 2.0 ± 0.0 | 4.0 ± 0.0 | 4.0 |
| Tylosin | 4.0 ± 0.0 | 4.0 ± 0.0 | 4.0 |
| Vancomycin | 2.0 ± 0.0 | 2.0 ± 0.0 | 4.0 |
| Strains | Hyaluronidase | Gelatinase |
|---|---|---|
| MG5206 | (-) | (-) |
| MG5232 | (-) | (-) |
| Strains | Autoaggregation (%) | Adhesion to Solvents (%) | ||
|---|---|---|---|---|
| Xylene | Chloroform | Ethyl Acetate | ||
| MG5206 | 37.20 ± 3.40 | 65.31 ± 1.01 | 94.89 ± 1.73 | 11.90 ± 1.66 |
| MG5232 | 66.30 ± 1.30 | 12.69 ± 1.18 | 28.88 ± 1.95 | 54.60 ± 15.73 |
| Strains | Viable Counts (log CFU/mL) | |||
|---|---|---|---|---|
| Simulated Gastric Fluid 1 | Simulated Intestinal Fluid 2 | |||
| pH 3 | pH 4 | pH 7 | pH 8 | |
| MG5206 | 4.79 ± 0.01 | 7.84 ± 0.03 | 7.92 ± 0.01 | 7.90 ± 0.02 |
| MG5232 | 6.93 ± 0.04 | 7.62 ± 0.06 | 7.56 ± 0.04 | 7.58 ± 0.02 |
| Enzymes | Substrate | MG5206 | MG5232 |
|---|---|---|---|
| Alkaline phosphatase | 2-naphthyl phosphate | 0 | 0 |
| Esterase (C4) | 2-naphthyl butyrate | 3 | 3 |
| Esterase Lipase (C8) | 2-naphthyl caprylate | 2 | 2 |
| Lipase (C14) | 2-naphthyl myristate | 0 | 0 |
| Leucine arylamidase | L- leucyl-2-naphthylamide | 3 | 3 |
| Valine arylamidase | L-valyl-2-naphthylamide | 0 | 1 |
| Crystine arylamidase | L-cystyl-2-naphthylamide | 0 | 2 |
| Trypsin | N-benzoyl-DL-arginine-2-naphthylamide | 0 | 0 |
| α-chymotrypsin | N-glutaryl-phenylanine-2-naphthylamide | 1 | 0 |
| Acid phosphatase | 2-naphtyl phosphate | 2 | 2 |
| Naphthol-AS-BI-phosphohydrolase | Naphthol-AS-BI-phosphate | 3 | 2 |
| α-galactosidase | 6-Br-2-naphthyl-αD-galactopyranoside | 0 | 0 |
| β- galactosidase | 2-naphthyl-βD-galactopyranoside | 0 | 0 |
| β-glucuronidase | Naphthol-AS-BI-βD-glucuronide | 0 | 0 |
| α-glucosidase | 2-naphthyl-αD-glucopyranoside | 0 | 0 |
| β-glucosidase | 6-Br-2-naphthyl- βD-glucopyranoside | 0 | 0 |
| N-acetyl-β-glucosaminidase | 1-naphthyl-N-acetyl-βD-glucosaminide | 0 | 0 |
| α-mannosidase | 6-Br-2-naphthyl-αD-mannopyranoside | 0 | 0 |
| α-fucosidase | 2-naphthyl-αL-fucopyranoside | 0 | 0 |
| Substrate | MG5206 | MG5232 | Substrate | MG5206 | MG5232 |
|---|---|---|---|---|---|
| Glycerol | + 1 | + | Salicin | + | + |
| Erythritol | − 2 | − | D-cellobiose | + | + |
| D-arabinose | − | − | D-maltose | + | + |
| L-arabinose | + | + | D-lactose | + | + |
| D-ribose | + | + | D-melibiose | − | − |
| D-xylose | + | + | D-sucrose | + | + |
| L-xylose | − | − | D-trehalose | + | + |
| D-adonitol | − | − | Inulin | − | − |
| Methyl-β D-xylopyranoside | − | − | D-melezitose | + | − |
| D-galactose | + | + | D-raffinose | − | − |
| D-glucose | + | + | Starch | + | − |
| D-fructose | + | + | Glycogen | − | − |
| D-mannose | + | + | Xylitol | − | − |
| L-sorbose | − | − | Gentiobiose | + | + |
| L-rhamnose | + | − | D-turanose | − | − |
| Dulcitol | − | − | D-lyxose | − | − |
| Inositol | − | − | D-tagatose | + | − |
| D-mannitol | + | + | D-fucose | − | − |
| D-sorbitol | + | + | L-fucose | − | − |
| Methyl-α D-mannoside | + | + | D-arabitol | − | − |
| Methyl-α D-glucoside | − | − | L-arabitol | − | − |
| N-acetyl-glucosamine | + | + | Gluconate | + | − |
| Amygdalin | + | + | 2-keto-gluconate | − | − |
| Arbutin | + | + | 5-keto-gluconate | − | − |
| Esculin | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Choi, S.-I.; Jeong, Y.; Kang, C.-H. Evaluation of Safety and Probiotic Potential of Enterococcus faecalis MG5206 and Enterococcus faecium MG5232 Isolated from Kimchi, a Korean Fermented Cabbage. Microorganisms 2022, 10, 2070. https://doi.org/10.3390/microorganisms10102070
Kim Y, Choi S-I, Jeong Y, Kang C-H. Evaluation of Safety and Probiotic Potential of Enterococcus faecalis MG5206 and Enterococcus faecium MG5232 Isolated from Kimchi, a Korean Fermented Cabbage. Microorganisms. 2022; 10(10):2070. https://doi.org/10.3390/microorganisms10102070
Chicago/Turabian StyleKim, YongGyeong, Soo-Im Choi, Yulah Jeong, and Chang-Ho Kang. 2022. "Evaluation of Safety and Probiotic Potential of Enterococcus faecalis MG5206 and Enterococcus faecium MG5232 Isolated from Kimchi, a Korean Fermented Cabbage" Microorganisms 10, no. 10: 2070. https://doi.org/10.3390/microorganisms10102070