
Fermentation of Microalgal Biomass for Innovative Food Production

 , and
, and
Abstract
:1. Introduction
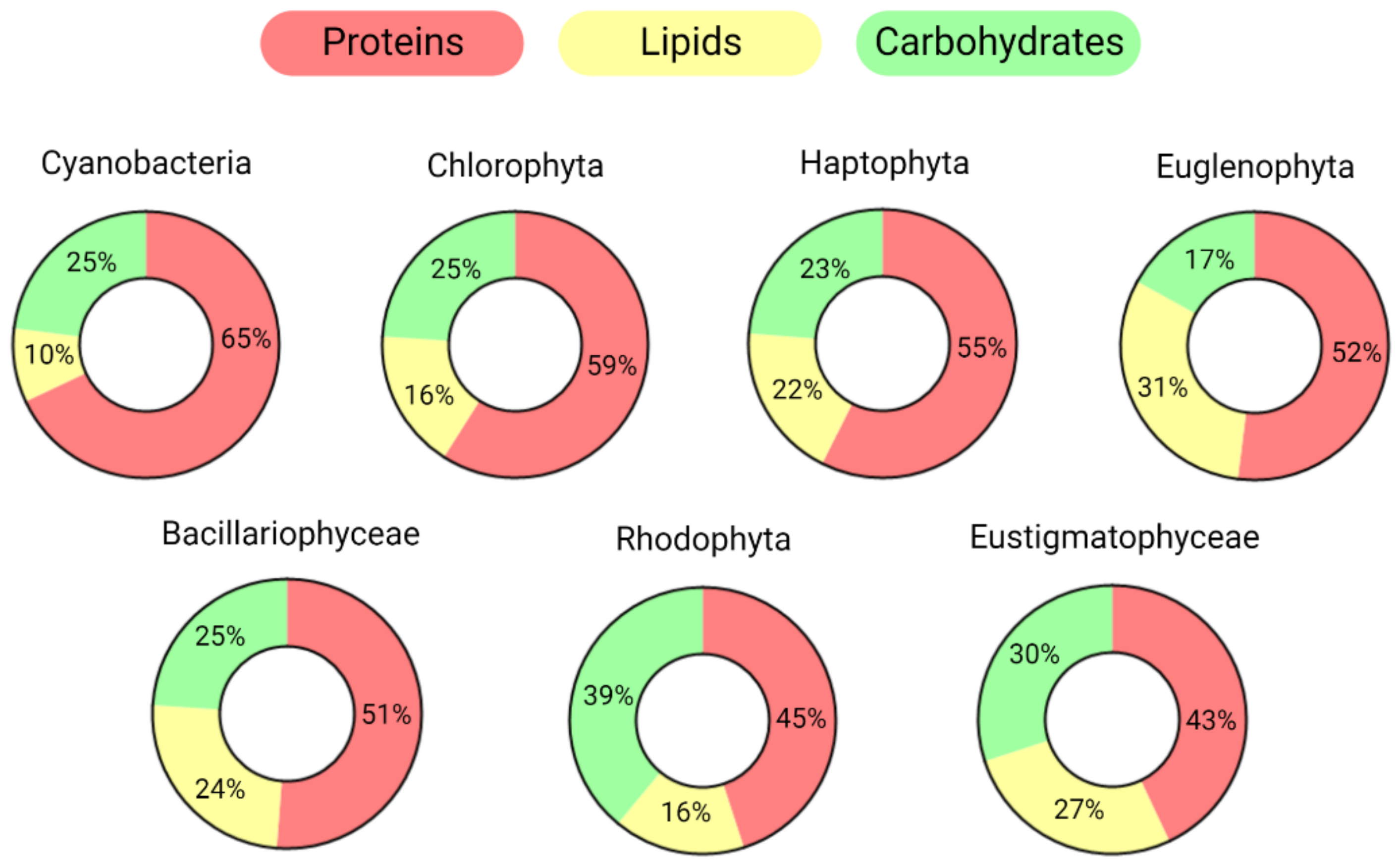
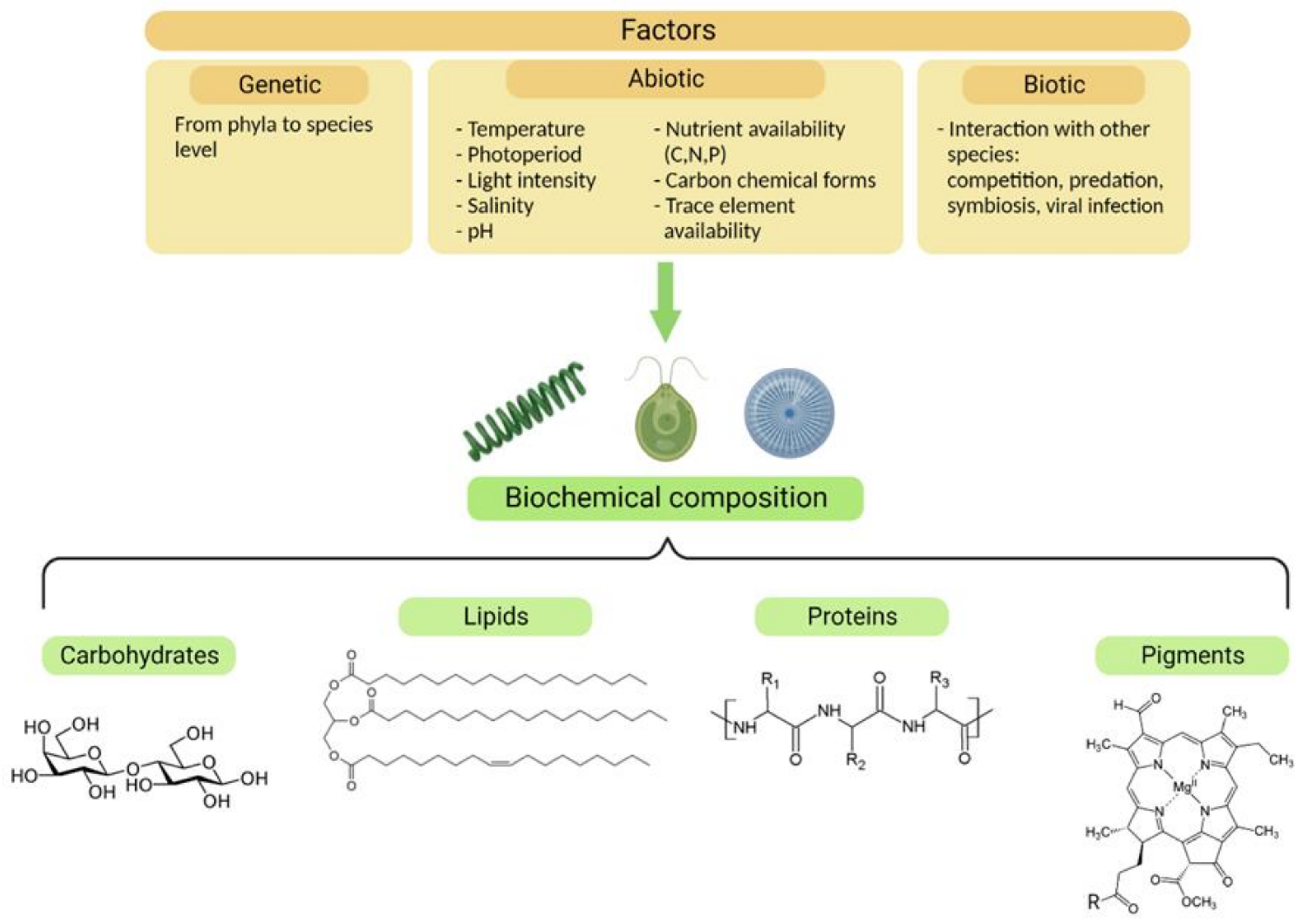
1.1. Microalgae’s Properties and Health Benefits

{kind=link}
{kind=link}
{kind=link}
| Microalgae | Product | Application | Effect on Human Health | Reference |
|---|---|---|---|---|
| Arthrospira platensis (Spirulina) | Biomass | Nutritional supplements and food ingredient | High protein content and rich in essential amino-acids. Rich in Fe, mineral elements, and vitamins | [33,34] |
| Phycocyanin | Nutritional supplements | Antioxidant, anti-inflammatory | [35] | |
| EPS | Medical applications | Anti-thrombotic and anti-tumoral | [6] | |
| Aphanizomenon flos-aquae | Biomass | Nutritional supplements | High protein content and essential fatty acids (Ω3) | [36] |
| Chlorella vulgaris | Biomass | Nutritional supplements and food ingredient | High protein and β-glucan content. Anti-inflammatory and anti-oxidant. | [37] |
| Dunaliella salina | β-carotene | Nutritional supplements and food ingredient | Antioxidant, pro-vitamin A, anti-allergic, anti-inflammatory | [38] |
| Dunaliella tertioletca | Vitamin A, B, E | Nutritional supplements | Maintenance of effective vision, protection against anemia and support of brain function, anti-oxidant | [39] |
| Haematococcus pluvialis | Astaxanthin | Nutritional supplements or supplements in aquaculture | Antioxidant, anti-inflammatory | [40] |
| Isochrysis galbana | EPA and DHA (Ω3) | Nutritional supplements or supplements in aquaculture | Anti-inflammatory, cardiovascular benefits, improves nervous system, atherosclerosis protection | [41,42,43] |
| Cryptothecodininum cohnii | DHA (Ω3) | Nutritional supplements or supplements in aquaculture | Cardiovascular benefits and improves nervous system | |
| Schizochytriumlimacinum | DHA (Ω3) | Nutritional supplements or supplements in aquaculture | Cardiovascular benefits and improves nervous system | |
| Nannochloropsis oceanica | EPA (Ω3) | Nutritional supplements or supplements in aquaculture | Cardiovascular benefits and protection against atherosclerosis, anti-inflammatory | |
| Porphyridium cruentum | ARA (Ω6) | Nutritional supplements or supplements in aquaculture | Improve functional development in infants | [44] |
| Porphyridium purpureum | EPS | Medical application | Anti-thrombotic and anti-tumoral | [6] |
| Phaeodactylum tricornutum | Fucoxanthin | Medical application | Anti-tumoral and beneficial effect against obesity | [45] |
| Euglena gracilis | Paramylon | Medical application | Anti-inflammatory and anti-tumoral | [27] |
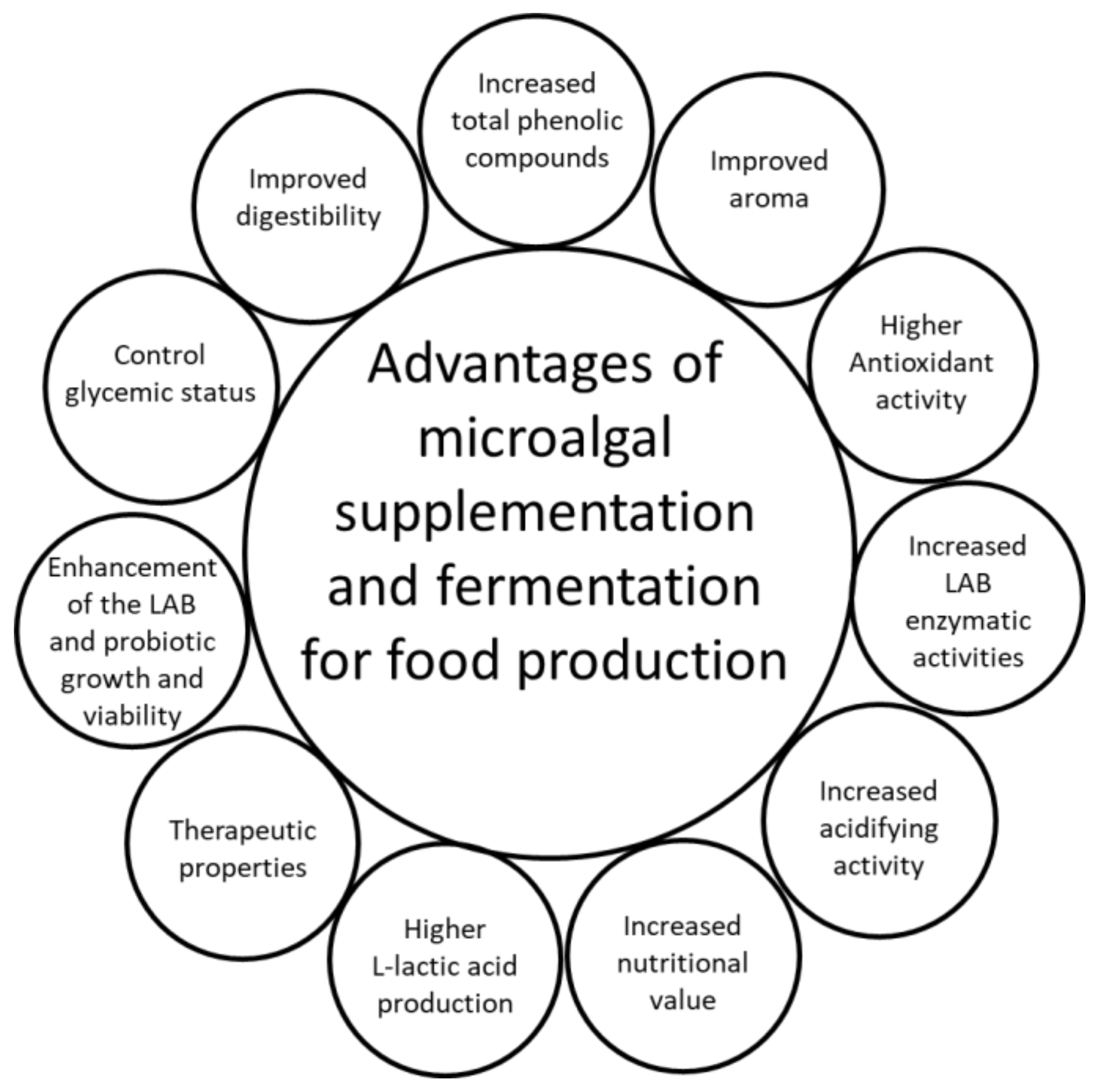
1.2. Advantages of Fermentation
2. Methodology
2.1. Search Procedure
2.2. Database Generation
3. Results and Discussion
3.1. Literature Review of Microalgal Fermentation for Novel Food Production
| Application | Microalga Species | Formulation | Concentration | Microbial Inoculum/Starter Cultures | Fermentation Conditions | Storage Conditions | Food Product | Reference |
|---|---|---|---|---|---|---|---|---|
| Microalgae as microbial growth promoter | Arthrospira platensis | Filtrate | 1:1 filtrate added to synthetic media | Lactococcus lactis subsp. lactis C2, Lactococcus casei YK3, Lactobacillus delbruekii subsp. bulgaricus YL1, Streptococcus salivarius subsp. thermophilus TH4, Lactobacillus spp. JL2 | 37 °C, 24 h | n.d. | n.s. | [50] |
| Arthrospira platensis | Extract | 3 mg/mL in fermented milk | Streptococcus thermophilus TH4, Lactococcus lactis subsp. lactis C2, Lactobacillus delbruekii subsp. bulgaricus YL1, Lactobacillus acidophilus LO1 | 37 °C, 20 h | n.d. | Fermented milk | [51] | |
| Arthrospira platensis | Powder | 3g/L in milk | Lactobacillus acidophilus, Bifidobacterium spp., Streptococcus thermophilus | 40 °C, 6 h | 4 °C, 42 days, and 15 °C, 18 days | Fermented ABT milk | [52] | |
| Arthrospira platensis; Chlorella vulgaris | Spray-dried biomass | 3 g/L in milk | Lactiplantibacillus plantarum, Enterococcus faecium | 30 °C or 37 °C | n.d. | Fermented milk | [53] | |
| Arthrospira platensis | Powder | 1, 5, 10 mg/mL | Lactobacillus acidophilus MTCC447, Streptococcus thermophilus MTCC1938, Lacticaseibacillus casei MTCC1423 | 37 °C, 10 h | n.d. | n.s. | [54] | |
| Arthrospira platensis | Powder | 0.5, 1.0% (w/v) in milk | Bifidobacterium animalis subsp. lactis Bb12, Lactobacillus acidophilus La-5 | 42 °C, 6 h | 5 ± 1 °C, 15 days | Fermented milks | [55] | |
| Microalgae supplementation to fermented foods | Arthrospira platensis | Powder | 0.5, 1.0% (w/v) in milk | Streptococcus thermophylus, Lactobacillus delbrueckii subsp. bulgaricus, Lactobacillus acidophilus | 40 °C and 42 °C, until the pH 4.7 was reached | 4 °C, 30 days | Yogurt and acidophilus milk | [56] |
| Arthrospira platensis; Chlorella vulgaris | Powder | 0.25, 0.5, 1.0% (w/v) in milk | Lactobacillus acidophilus LA-5, Bifidobacterium lactis BB-12, Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus | 40 °C until the pH 4.7 was reached | 5 °C, 28 days | Probiotic yogurt | [57] | |
| Arthrospira platensis | Powder | 0.3, 0.5, 0.8% in milk | Lactococcus lactis subsp. lactis, Lactococcus lactis subsp. cremoris, Lactobacillus acidophilus LA-5 | 30 °C, 48 h | 4 °C, 45 days | Probiotic feta cheese | [58] | |
| Arthrospira platensis | Powder | 0.25, 0.5, 0.75, 1.0% (w/v) in milk | Lactobacillus bulgaricus, Streptococcus thermophilus | 42 °C, 4 h | 7, 14, 21, 28 days of refrigerated storage | Yogurt | [59] | |
| Arthrospira platensis | Grains | 0.5, 1.0, 1.5% | Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus | n.r. | n.d. | Kareish cheese | [60] | |
| Arthrospira platensis | Dry biomass | 1.0% (w/v) in soy milk | Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus | 37 °C, 24 h | 4 °C | Soy yogurt | [61] | |
| Arthrospira platensis | Dry biomass | 0.5, 1.0, 1.5% in soy milk | Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus | 37 °C, 16 h | 4 °C | Soy yogurt | [62] | |
| Arthrospira platensis | Powder | 5 mg/mL in milk | Streptococcus salivarius subsp. thermophilus, Lactobacillus delbrueckii subsp. bulgaricus, Bifidobacterium spp. | 37 °C, 48 h | n.d. | Labneh | [63] | |
| Arthrospira platensis | Fresh vs oven-dried | 0.1, 0.3, 0.5% (w/v) in milk | Streptococcus thermophilus, Lactobacillus bulgaricus | 42 °C, 4 h | 4 °C, 24 h | Yogurt | [64] | |
| Arthrospira platensis | Powder | 0.5, 1.0, 1.5% (w/w) | Lacticaseibacillus casei, Lactococcus lactis subsp. lactis, Lactococcus lactis subsp. cremoris | 40 °C until the pH 5.2 was reached | 4 °C, 60 days | Feta-type cheese | [65] | |
| Arthrospira platensis SP6/CFTRI | Fresh wet biomass (spirulina milk emulsion) | 10% (w/w) | Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus, Lactobacillus acidophilus | 42 °C until the pH 4.6–4.7 was reached | 6–8 °C | Probiotic yogurt | [66] | |
| Arthrospira platensis | Powder | 0.25, 0.50, 1.0% (w/v) in milk | Streptococcus thermophilus, Lactobacillus delbrueckii subsp. bulgaricus, Lactobacillus acidophilus, Bifidobacterium lactis | 40 °C until the pH 4.4 was reached (about 4 h) | 4 ± 1 °C, 21 days | Ayran | [67,68] | |
| Arthrospira platensis | Powder | 1.0% (w/w) | Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus | 42 °C until the pH 4.6 was reached | 4 °C | Low-fat yogurt | [69] | |
| Arthrospira platensis F&M-C256 | Lyophilized biomass | 10% (w/v) in vegetal soybean drink or water | Lactiplantibacillus plantarum ATCC 8014 | 37 °C, 72 h, 100 rpm stirring | n.d. | Vegetal soybean drink | [70] | |
| Arthrospira platensis | Dried biomass | n.s. | Lacticaseibacillus paracasei | 40 °C until the pH 4.7 was reached | 4 °C | Probiotic yogurt | [71] | |
| Arthrospira platensis | Algae biomass in water (5% w/v) | 0.25, 0.50% (w/v) in skim milk powder (SSM) and in commercial soy-based beverages (SBB) | Lactococcus lactis subsp. lactis, Lactococcus lactis subsp. cremoris, Lactococcus casei, Lactobacillus delbruekii subsp. bulgaricus, Streptococcus thermophilus, Lacticaseibacillus rhamnosus, Lactobacillus helveticus, Lactobacillus delbruekii subsp. lactis, Weissella spp., Leuconostoc spp. | 37 °C, 48 h | n.d. | Milk and soy fermented beverages | [72] | |
| Arthrospira spp. | Dry biomass | 1.0, 2.0% (w/v) in milk | Milk kefir grains | 25–37 °C, 24 h | n.d. | Milk kefir | [73] | |
| Arthrospira platensis | Powder | 0.25, 0.50% in almond milk and soy milk | Lactobacilli and lactococci plant-based kefir culture | 42 °C until a pH 4.5 ± 0.02 was reached | 4 °C, 21 days | Vegan kefir (soy milk kefir and almond milk kefir) | [74] | |
| Arthrospira platensis | Powder | 0.25, 0.5, 1.0 g/kg | Yogurt culture | 20 °C, 24 h and then 12 °C, 48 h | n.d. | Greek soft cheese | [75] | |
| Chlorella vulgaris | Powder | 1.5% (w/v) in soya drink | Levilactobacillus brevis ŁOCK 0944 | 30 °C, 4 h and then matured at 18 °C, 20 h | n.d. | Soya drink | [76] | |
| Arthrospira spp. | Lyophilized biomass | 1.6 g, 2.4 g in 100 mL of distilled water | Water kefir grains | 25 °C, 48 h | n.d. | Water kefir | [77] | |
| Microalgae as the sole substrate for fermentation | Pavlova lutheri | Powder | 1:15 (w/v) | Hansenula polymorpha | 37 °C, 12 days | n.d. | n.s. | [78] |
| Arthrospira platensis | Powder | 2.0% (w/v) | Lactiplantibacillus plantarum B7, Lactiplantibacillus plantarum C8-1, Lactiplantibacillus plantarum 121, Lactobacillus acidophilus NCFM, Bacillus subtilis 168 | 37 °C, 24 h | n.d. | n.s. | [79] | |
| Arthrospira maxima | Powder | 10% (w/v) | Lactiplantibacillus plantarum HY-08 | 37 °C, 4 days | n.d. | n.s. | [80] | |
| Arthrospira maxima | Powder | 10% (w/v) | Lactiplantibacillus plantarum HY-08 | 37 °C | n.d. | n.s. | [81] | |
| Arthrospira platensis | Wet biomass | 5 g in 30 mL of distilled water | Lactiplantibacillus plantarum | 37 °C, 72 h in a shaker | n.d. | Fermented nutraceutical product | [47] | |
| Arthrospira platensis F&M-C256 | Lyophilized biomass | 10% (w/v) | Lactiplantibacillus plantarum ATCC 8014 | 37 °C, 72 h, 100 rpm stirring | n.d. | Probiotic-based products | [82] | |
| Chlorella vulgaris | Powder | 0.1, 1.5% (w/v) | Levilactobacillus brevis ŁOCK 0944, Levilactobacillus brevis ŁOCK 0980, Levilactobacillus brevis ŁOCK 0992, Levilactobacillus brevis MG451814 | 30 °C, 24 h | n.d. | n.s. | [83] | |
| Arthrospira platensis | Powder | 2.0% (w/v) | Lactiplantibacillus plantarum DY-1, Lacticaseibacillus casei KDB-LC, Lactobacillus acidophilus KDB-03, Lactobacillus acidophilus KDB-08, Bacillus subtilis ND, Bacillus spp., Bacillus amyloliquefaciens LXZ | 37 °C, 72 h | n.d. | n.s. | [84] | |
| Arthrospira platensis | Dehydrated biomass | n.s. | Lacticaseibacillus casei 2240, Lacticaseibacillus rhamnosus GG | 37 °C, 48 h | n.d. | New fermented food supplements | [85] | |
| Nostochopsis lobatus, Nostoc commune, Nostoc flagelliforme, Nostoc verrucosum, Arthrospira platensis, Dunaliella tertiolecta, Chlorogonium spp., Porphyridium purpureum, Pleurochrysis carterae, Euglena spp. | Powder | 10% (w/v) | Lactococcus lactis subsp. lactis, Lactiplantibacillus plantarum | 37 °C, 72 h | n.d. | n.s. | [86] | |
| Arthrospira platensis | Fresh | 10 g with 10 mL of physiological solution | Lactiplantibacillus plantarum | 30°C, 72 h | n.d. | n.s. | [87] | |
| Arthrospira platensis | Powder | 4.0% (w/v) | Debaryomyces hansenii, Kluyveromyces marxianus, Saccharomyces cerevisiae | 28 °C, 48 h, 130 rpm stirring | n.d. | n.s. | [88] | |
| Arthrospira platensis | Dried powder | 4.0% (w/v) | Lactobacillus helveticus B-4526, Lactiplantibacillus plantarum B531, Lacticaseibacillus rhamnosus B-442, Lacticaseibacillus casei B-1922, Bacillus subtilis B-3384, Bacillus subtilis B-3387, Bacillus licheniformis NRS-1264 | LAB strains at 37°C, 48 h, 100 rpm stirring; Bacillus strains at 28°C, 48 h, 130 rpm stirring | n.d. | n.s. | [89] |
3.2. Effect of the Addition of Microalgae on Growth and Viability of LAB and Probiotics
3.3. Development and Characterization of New Fermented Food Products Supplemented with Microalgae
3.4. Fermented Microalgae as Innovative Functional Foods or Ingredients
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rockström, J.; Edenhofer, O.; Gaertner, J.; DeClerck, F. Planet-Proofing the Global Food System. Nat. Food 2020, 1, 3–5. [Google Scholar] [CrossRef] [Green Version]
- UN DESA. World Population Prospects 2022: Summary of Results 2022; UN DESA: New York, NY, USA, 2022. [Google Scholar]
- Koyande, A.K.; Chew, K.W.; Rambabu, K.; Tao, Y.; Chu, D.T.; Show, P.L. Microalgae: A Potential Alternative to Health Supplementation for Humans. Food Sci. Hum. Wellness 2019, 8, 16–24. [Google Scholar] [CrossRef]
- Fabris, M.; Abbriano, R.M.; Pernice, M.; Sutherland, D.L.; Commault, A.S.; Hall, C.C.; Labeeuw, L.; McCauley, J.I.; Kuzhiuparambil, U.; Ray, P.; et al. Emerging Technologies in Algal Biotechnology: Toward the Establishment of a Sustainable, Algae-Based Bioeconomy. Front. Plant Sci. 2020, 11, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirbas, A. Progress and Recent Trends in Biodiesel Fuels. Energy Convers. Manag. 2009, 50, 14–34. [Google Scholar] [CrossRef]
- Pierre, G.; Delattre, C.; Dubessay, P.; Jubeau, S.; Vialleix, C.; Cadoret, J.P.; Probert, I.; Michaud, P. What Is in Store for EPS Microalgae in the Next Decade? Molecules 2019, 24, 4296. [Google Scholar] [CrossRef] [Green Version]
- Keeling, P.J. Diversity and Evolutionary History of Plastids and Their Hosts. Am. J. Bot. 2004, 91, 1481–1493. [Google Scholar] [CrossRef] [Green Version]
- Perez-Garcia, O.; Escalante, F.M.E.; de-Bashan, L.E.; Bashan, Y. Heterotrophic Cultures of Microalgae: Metabolism and Potential Products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef]
- Enzing, C.; Ploeg, M.; Barbosa, M.; Sijtsma, L. Microalgae-Based Products for the Food and Feed Sector: An Outlook for Europe; Publications Office of the European Union: Luxembourg, 2014; ISBN 978-92-79-34037-6. [Google Scholar]
- Varshney, P.; Mikulic, P.; Vonshak, A.; Beardall, J.; Wangikar, P.P. Extremophilic Micro-Algae and Their Potential Contribution in Biotechnology. Bioresour. Technol. 2015, 184, 363–372. [Google Scholar] [CrossRef]
- Finkel, Z.V.; Follows, M.J.; Liefer, J.D.; Brown, C.M.; Benner, I.; Irwin, A.J. Phylogenetic Diversity in the Macromolecular Composition of Microalgae. PLoS ONE 2016, 11, e0155977. [Google Scholar] [CrossRef] [Green Version]
- Christaki, E.; Florou-Paneri, P.; Bonos, E. Microalgae: A Novel Ingredient in Nutrition. Int. J. Food Sci. Nutr. 2011, 62, 794–799. [Google Scholar] [CrossRef]
- Niccolai, A.; Chini Zittelli, G.; Rodolfi, L.; Biondi, N.; Tredici, M.R. Microalgae of Interest as Food Source: Biochemical Composition and Digestibility. Algal Res. 2019, 42, 101617. [Google Scholar] [CrossRef]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for High-Value Products Towards Human Health and Nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef] [Green Version]
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a Future Food Source. Biotechnol. Adv. 2020, 41, 107536. [Google Scholar] [CrossRef]
- Molino, A.; Iovine, A.; Casella, P.; Mehariya, S.; Chianese, S.; Cerbone, A.; Rimauro, J.; Musmarra, D. Microalgae Characterization for Consolidated and New Application in Human Food, Animal Feed and Nutraceuticals. Int. J. Environ. Res. Public Health 2018, 15, 2436. [Google Scholar] [CrossRef] [Green Version]
- Procházková, G.; Brányiková, I.; Zachleder, V.; Brányik, T. Effect of Nutrient Supply Status on Biomass Composition of Eukaryotic Green Microalgae. J. Appl. Phycol. 2014, 26, 1359–1377. [Google Scholar] [CrossRef]
- Giordano, M.; Olivieri, C.; Ratti, S.; Norici, A.; Raven, J.A.; Knoll, A.H. A tale of two eras: Phytoplankton composition influenced by oceanic paleochemistry. Geobiology 2018, 16, 498–506. [Google Scholar] [CrossRef]
- Giordano, M.; Palmucci, M.; Norici, A. Taxonomy and growth conditions concur to determine the energetic suitability of fatty acid complements in algae. J. Appl. Phycol. 2015, 27, 1401–1413. [Google Scholar] [CrossRef]
- Ratti, S.; Knoll, A.H.; Giordano, M. Grazers and Phytoplankton Growth in the Oceans: An Experimental and Evolutionary Perspective. PLoS ONE 2013, 8, e77349. [Google Scholar] [CrossRef]
- Petrucciani, A.; Chaerle, P.; Norici, A. Diatoms Versus Copepods: Could Frustule Traits Have a Role in Avoiding Predation? Front. Mar. Sci. 2022, 8, 804960. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Monteiro, C.M.; Malcata, F.X. Simultaneous Effect of Irradiance and Temperature on Biochemical Composition of the Microalga Pavlova Lutheri. J. Appl. Phycol. 2009, 21, 543–552. [Google Scholar] [CrossRef]
- Klin, M.; Pniewski, F.; Latała, A. Growth Phase-Dependent Biochemical Composition of Green Microalgae: Theoretical Considerations for Biogas Production. Bioresour. Technol. 2020, 303, 122875. [Google Scholar] [CrossRef] [PubMed]
- Hockin, N.L.; Mock, T.; Mulholland, F.; Kopriva, S.; Malin, G. The Response of Diatom Central Carbon Metabolism to Nitrogen Starvation Is Different from That of Green Algae and Higher Plants. Plant Physiol. 2012, 158, 299–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q. Environmental Effects on Cell Composition. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons, Ltd.: Oxford, UK, 2013; pp. 114–122. ISBN 978-1-118-56716-6. [Google Scholar]
- Borowitzka, M.A. Microalgae in Medicine and Human Health. In Microalgae in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2018; pp. 195–210. ISBN 978-0-12-811405-6. [Google Scholar]
- Barsanti, L.; Birindelli, L.; Gualtieri, P. Paramylon and Other Bioactive Molecules in Micro and Macroalgae. Int. J. Mol. Sci. 2022, 23, 8301. [Google Scholar] [CrossRef] [PubMed]
- Araújo, R.; Peteiro, C. Algae as Food and Food Supplements in Europe; EUR 30779 EN; Publications Office of the European Union: Luxembourg, 2021; ISBN 978-92-76-40548-1. [Google Scholar]
- Grosshagauer, S.; Kraemer, K.; Somoza, V. The True Value of Spirulina. J. Agric. Food Chem. 2020, 68, 4109–4115. [Google Scholar] [CrossRef]
- Martelli, F.; Cirlini, M.; Lazzi, C.; Neviani, E.; Bernini, V. Edible Seaweeds and Spirulina Extracts for Food Application: In Vitro and In Situ Evaluation of Antimicrobial Activity towards Foodborne Pathogenic Bacteria. Foods 2020, 9, 1442. [Google Scholar] [CrossRef]
- Bito, T.; Okumura, E.; Fujishima, M.; Watanabe, F. Potential of Chlorella as a Dietary Supplement to Promote Human Health. Nutrients 2020, 12, 2524. [Google Scholar] [CrossRef]
- Gilroy, D.J.; Kauffman, K.W.; Hall, R.A.; Huang, X.; ChU, F.S. Assessing Potential Health Risks from Microcystin Toxins in Blue-Green Algae Dietary Supplements. Environ. Health Perspect. 2000, 1081, 5. [Google Scholar] [CrossRef]
- Ovando, C.A.; de Carvalho, J.C.; Vinícius de Melo Pereira, G.; Jacques, P.; Soccol, V.T.; Soccol, C.R. Functional Properties and Health Benefits of Bioactive Peptides Derived from Spirulina: A Review. Food Rev. Int. 2018, 34, 34–51. [Google Scholar] [CrossRef]
- Capelli, B.; Cysewski, G.R. Potential Health Benefits of Spirulina Microalgae: A Review of the Existing Literature. Nutrafoods 2010, 9, 19–26. [Google Scholar] [CrossRef]
- Kuddus, M.; Singh, P.; Thomas, G.; Al-Hazimi, A. Recent Developments in Production and Biotechnological Applications of C-Phycocyanin. BioMed Res. Int. 2013, 2013, 1–9. [Google Scholar] [CrossRef]
- Lyon-Colbert, A.; Su, S.; Cude, C. A Systematic Literature Review for Evidence of Aphanizomenon Flos-Aquae Toxigenicity in Recreational Waters and Toxicity of Dietary Supplements: 2000–2017. Toxins 2018, 10, 254. [Google Scholar] [CrossRef] [Green Version]
- Rani, K.; Sandal, N.; Sahoo, P. A Comprehensive Review on Chlorella-Its Composition, Health Benefits, Market and Regulatory Scenario. Pharma Innov. 2018, 7, 584–589. [Google Scholar]
- Murthy, K.N.C.; Vanitha, A.; Rajesha, J.; Swamy, M.M.; Sowmya, P.R.; Ravishankar, G.A. In Vivo Antioxidant Activity of Carotenoids from Dunaliella Salina—A Green Microalga. Life Sci. 2005, 76, 1381–1390. [Google Scholar] [CrossRef]
- Abalde, J.; Fabregas, J. β-Carotene, Vitamin C and Vitamin E Content of the Marine Microalga Dunaliella Tertiolecta Cultured with Different Nitrogen Sources. Bioresour. Technol. 1991, 38, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Mularczyk, M.; Michalak, I.; Marycz, K. Astaxanthin and Other Nutrients from Haematococcus Pluvialis—Multifunctional Applications. Mar. Drugs 2020, 18, 459. [Google Scholar] [CrossRef]
- Zhou, L.; Li, K.; Duan, X.; Hill, D.; Barrow, C.; Dunshea, F.; Martin, G.; Suleria, H. Bioactive Compounds in Microalgae and Their Potential Health Benefits. Food Biosci. 2022, 49, 101932. [Google Scholar] [CrossRef]
- Liu, Y.; Ren, X.; Fan, C.; Wu, W.; Zhang, W.; Wang, Y. Health Benefits, Food Applications, and Sustainability of Microalgae-Derived N-3 PUFA. Foods 2022, 11, 1883. [Google Scholar] [CrossRef]
- Khozin-Goldberg, I.; Iskandarov, U.; Cohen, Z. LC-PUFA from Photosynthetic Microalgae: Occurrence, Biosynthesis, and Prospects in Biotechnology. Appl. Microbiol. Biotechnol. 2011, 91, 905–915. [Google Scholar] [CrossRef]
- Shanab, S.M.M.; Hafez, R.M.; Fouad, A.S. A Review on Algae and Plants as Potential Source of Arachidonic Acid. J. Adv. Res. 2018, 11, 3–13. [Google Scholar] [CrossRef]
- Eilers, U.; Bikoulis, A.; Breitenbach, J.; Büchel, C.; Sandmann, G. Limitations in the Biosynthesis of Fucoxanthin as Targets for Genetic Engineering in Phaeodactylum Tricornutum. J. Appl. Phycol. 2016, 28, 123–129. [Google Scholar] [CrossRef]
- Pérez-Alva, A.; MacIntosh, A.J.; Baigts-Allende, D.K.; García-Torres, R.; Ramírez-Rodrigues, M.M. Fermentation of Algae to Enhance Their Bioactive Activity: A Review. Algal. Res. 2022, 64, 102684. [Google Scholar] [CrossRef]
- De Marco Castro, E.; Shannon, E.; Abu-Ghannam, N. Effect of Fermentation on Enhancing the Nutraceutical Properties of Arthrospira platensis (spirulina). Fermentation 2019, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Peng, K.; Koubaa, M.; Bals, O.; Vorobiev, E. Recent Insights in the Impact of Emerging Technologies on Lactic Acid Bacteria: A Review. Food Res. Int. 2020, 137, 109544. [Google Scholar] [CrossRef]
- Parada, J. Lactic Acid Bacteria Growth Promoters from Spirulina platensis. Int. J. Food Microbiol. 1998, 45, 225–228. [Google Scholar] [CrossRef]
- De Caire, G.Z.; Parada, J.L.; Zaccaro, M.C.; de Cano, M.M.S. Effect of Spirulina platensis Biomass on the Growth of Lactic Acid Bacteria in Milk. World J. Microbiol. Biotechnol. 2000, 16, 563–565. [Google Scholar] [CrossRef]
- Varga, L.; Szigeti, J.; Kovács, R.; Földes, T.; Buti, S. Influence of a Spirulina platensis Biomass on the Microflora of Fermented ABT Milks During Storage (R1). J. Dairy Sci. 2002, 85, 1031–1038. [Google Scholar] [CrossRef]
- Gyenis, B.; Szigeti, J.; Molnár, N.; Varga, L. Use of Dried Microalgal Biomasses to Stimulate Acid Production and Growth of Lactobacillus plantarum and Enterococcus faecium in Milk. Acta Agrar. Kaposváriensis 2005, 9, 53–59. [Google Scholar]
- Bhowmik, D.; Dubey, J.; Mehra, S. Probiotic Efficiency of Spirulina platensis–Stimulating Growth of Lactic Acid Bacteria. World J. Dairy Food Sci. 2009, 4, 160–163. [Google Scholar]
- Mocanu, G.D.; Botez, E.; Nistor, O.V.; Georgeta, D.; Vlăsceanu, G. Influence of Spirulina platensis Biomass over Some Starter Culture of Lactic Bacteria. J. Agroaliment. Process. Technol. 2013, 19, 474–479. [Google Scholar]
- Guldas, M.; Irkin, R. Influence of Spirulina platensis Powder on the Microflora of Yoghurt and Acidophilus Milk. Mljekarstvo 2010, 60, 237–243. [Google Scholar]
- Beheshtipour, H.; Mortazavian, A.M.; Mohammadi, R.; Sohrabvandi, S.; Khosravi-Darani, K. Supplementation of Spirulina platensis and Chlorella vulgaris Algae into Probiotic Fermented Milks: Algae Addition into Probiotic Fermented Milks. Compr. Rev. Food Sci. Food Saf. 2013, 12, 144–154. [Google Scholar] [CrossRef]
- Mazinani, S.; Fadaei, V.; Khosravi-Darani, K. Impact of Spirulina platensis on Physicochemical Properties and Viability of Lactobacillus acidophilus of Probiotic UF Feta Cheese: Microalgal Incorporation Probiotic UF Feta Cheese. J. Food Process. Preserv. 2016, 40, 1318–1324. [Google Scholar] [CrossRef]
- Barkallah, M.; Dammak, M.; Louati, I.; Hentati, F.; Hadrich, B.; Mechichi, T.; Ayadi, M.A.; Fendri, I.; Attia, H.; Abdelkafi, S. Effect of Spirulina platensis Fortification on Physicochemical, Textural, Antioxidant and Sensory Properties of Yogurt during Fermentation and Storage. LWT 2017, 84, 323–330. [Google Scholar] [CrossRef]
- Darwish, A.M.I. Physicochemical Properties, Bioactive Compounds and Antioxidant Activity of Kareish Cheese Fortified with Spirulina platensis. World J. Dairy Food Sci. 2017, 12, 71–78. [Google Scholar]
- Sengupta, S.; Bhowal, J. Optimization of Ingredient and Processing Parameter for the Production of Spirulina platensis Incorporated Soy Yogurt Using Response Surface Methodology. J. Microb. Biotechnol. Food Sci. 2017, 6, 1081–1085. [Google Scholar] [CrossRef]
- Sengupta, S.; Koley, H.; Dutta, S.; Bhowal, J. Hypocholesterolemic Effect of Spirulina platensis (SP) Fortified Functional Soy Yogurts on Diet-Induced Hypercholesterolemia. J. Funct. Foods 2018, 48, 54–64. [Google Scholar] [CrossRef]
- Abbas, M.H.; Farahat, E.S.A.K.; Zaky, W.M.; Mohamed, A.G. Applicability of Using Edible Algae (Spirulina platensis) to Prepare High Protein Quality Labenah. J. Biol. Sci. 2019, 19, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Bchir, B.; Felfoul, I.; Bouaziz, M.A.; Gharred, T.; Yaich, H.; Noumi, E.; Snoussi, M.; Bejaoui, H.; Kenzali, Y.; Blecker, C.; et al. Investigation of Physicochemical, Nutritional, Textural, and Sensory Properties of Yoghurt Fortified with Fresh and Dried spirulina (Arthrospira platensis). Int. Food Res. J. 2019, 26, 1565–1576. [Google Scholar]
- Golmakani, M.T.; Soleimanian-Zad, S.; Alavi, N.; Nazari, E.; Eskandari, M.H. Effect of Spirulina (Arthrospira platensis) Powder on Probiotic Bacteriologically Acidified Feta-Type Cheese. J. Appl. Phycol. 2019, 31, 1085–1094. [Google Scholar] [CrossRef]
- Patel, P.; Jethani, H.; Radha, C.; Vijayendra, S.V.N.; Mudliar, S.N.; Sarada, R.; Chauhan, V.S. Development of a Carotenoid Enriched Probiotic Yogurt from Fresh Biomass of Spirulina and Its Characterization. J. Food Sci. Technol. 2019, 56, 3721–3731. [Google Scholar] [CrossRef]
- Çelekli, A.; Alslibi, Z.A.; Bozkurt, H. üseyin. Influence of Incorporated Spirulina platensis on the Growth of Microflora and Physicochemical Properties of Ayran as a Functional Food. Algal Res. 2019, 44, 101710. [Google Scholar] [CrossRef]
- Çelekli, A.; Alslibi, Z.A.; Bozkurt, H. Boosting Effects of Spirulina platensis, Whey Protein, and Probiotics on the Growth of Microflora and the Nutritional Value of Ayran. Eng. Rep. 2020, 2, e12235. [Google Scholar] [CrossRef]
- Atallah, A.A.; Morsy, O.M.; Gemiel, D.G. Characterization of Functional Low-Fat Yogurt Enriched with Whey Protein Concentrate, Ca-Caseinate and Spirulina. Int. J. Food Prop. 2020, 23, 1678–1691. [Google Scholar] [CrossRef]
- Niccolai, A.; Bažec, K.; Rodolfi, L.; Biondi, N.; Zlatić, E.; Jamnik, P.; Tredici, M.R. Lactic Acid Fermentation of Arthrospira platensis (spirulina) in a Vegetal Soybean Drink for Developing New Functional Lactose-Free Beverages. Front. Microbiol. 2020, 11, 560684. [Google Scholar] [CrossRef]
- Alizadeh Khaledabad, M.; Ghasempour, Z.; Moghaddas Kia, E.; Rezazad Bari, M.; Zarrin, R. Probiotic Yoghurt Functionalised with Microalgae and Zedo Gum: Chemical, Microbiological, Rheological and Sensory Characteristics. Int. J. Dairy Technol. 2020, 73, 67–75. [Google Scholar] [CrossRef]
- Martelli, F.; Alinovi, M.; Bernini, V.; Gatti, M.; Bancalari, E. Arthrospira platensis as Natural Fermentation Booster for Milk and Soy Fermented Beverages. Foods 2020, 9, 350. [Google Scholar] [CrossRef]
- Laela, N.; Legowo, A.M.; Fulyani, F. The Effect of Kefir-Spirulina on Glycemic Status and Antioxidant Activity in Hyperglycemia Rats. Potr. S. J. F. Sci. 2021, 15, 101–110. [Google Scholar] [CrossRef]
- Sözeri Atik, D.; Gürbüz, B.; Bölük, E.; Palabıyık, İ. Development of Vegan Kefir Fortified with Spirulina platensis. Food Biosci. 2021, 42, 101050. [Google Scholar] [CrossRef]
- Bosnea, L.; Terpou, A.; Pappa, E.; Kondyli, E.; Mataragas, M.; Markou, G.; Katsaros, G. Incorporation of Spirulina platensis on Traditional Greek Soft Cheese with Respect to Its Nutritional and Sensory Perspectives. Proceedings 2021, 70, 99. [Google Scholar] [CrossRef]
- Ścieszka, S.; Gorzkiewicz, M.; Klewicka, E. Innovative Fermented Soya Drink with the Microalgae Chlorella vulgaris and the Probiotic Strain Levilactobacillus brevis ŁOCK 0944. LWT 2021, 151, 112131. [Google Scholar] [CrossRef]
- Nascimento, R.Q.; Deamici, K.M.; Tavares, P.P.L.G.; de Andrade, R.B.; Guimarães, L.C.; Costa, J.A.V.; Magalhães-Guedes, K.T.; Druzian, J.I.; de Souza, C.O. Improving Water Kefir Nutritional Quality via Addition of Viable Spirulina Biomass. Bioresour. Technol. Rep. 2022, 17, 100914. [Google Scholar] [CrossRef]
- Qian, Z.J.; Jung, W.K.; Kang, K.H.; Ryu, B.; Kim, S.K.; Je, J.Y.; Heo, S.J.; Oh, C.; Kang, D.H.; Park, W.S.; et al. In Vitro Antioxidant Activities of The Fermented Marine Microalga Pavlova lutheri (Haptophyta) With the Yeast Hansenula polymorpha. J. Phycol. 2012, 48, 475–482. [Google Scholar] [CrossRef]
- Bao, J.; Zhang, X.; Zheng, J.H.; Ren, D.F.; Lu, J. Mixed Fermentation of Spirulina platensis with Lactobacillus plantarum and Bacillus subtilis by Random-Centroid Optimization. Food Chem. 2018, 264, 64–72. [Google Scholar] [CrossRef]
- Choi, W.; Kang, D.; Heo, S.J.; Lee, H. Enhancement of the Neuroprotective Effect of Fermented Spirulina maxima Associated with Antioxidant Activities by Ultrasonic Extraction. Appl. Sci. 2018, 8, 2469. [Google Scholar] [CrossRef] [Green Version]
- Choi, W.Y.; Kang, D.H.; Lee, H.Y. Effect of Fermented Spirulina maxima Extract on Cognitive-Enhancing Activities in Mice with Scopolamine-Induced Dementia. Evid. Based Complement. Altern. Med. 2018, 7218504. [Google Scholar] [CrossRef] [Green Version]
- Niccolai, A.; Shannon, E.; Abu-Ghannam, N.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Lactic acid fermentation of Arthrospira platensis (Spirulina) biomass for probiotic-based products. J. Appl. Phycol. 2019, 31, 1077–1083. [Google Scholar] [CrossRef]
- Ścieszka, S.; Klewicka, E. Influence of the Microalga Chlorella vulgaris on the Growth and Metabolic Activity of Lactobacillus spp. Bacteria. Foods 2020, 9, 959. [Google Scholar] [CrossRef]
- Yu, J.; Ma, D.; Qu, S.; Liu, Y.; Xia, H.; Bian, F.; Zhang, Y.; Huang, C.; Wu, R.; Wu, J.; et al. Effects of Different Probiotic Combinations on the Components and Bioactivity of Spirulina. J. Basic Microbiol. 2020, 60, 543–557. [Google Scholar] [CrossRef]
- Martelli, F.; Cirlini, M.; Lazzi, C.; Neviani, E.; Bernini, V. Solid-State Fermentation of Arthrospira platensis to Implement New Food Products: Evaluation of Stabilization Treatments and Bacterial Growth on the Volatile Fraction. Foods 2020, 10, 67. [Google Scholar] [CrossRef]
- Kaga, Y.; Kuda, T.; Taniguchi, M.; Yamaguchi, Y.; Takenaka, H.; Takahashi, H.; Kimura, B. The Effects of Fermentation with Lactic Acid Bacteria on the Antioxidant and Anti-Glycation Properties of Edible Cyanobacteria and Microalgae. LWT 2021, 135, 110029. [Google Scholar] [CrossRef]
- Jamnik, P.; Mahnič, N.; Mrak, A.; Pogačnik, L.; Jeršek, B.; Niccolai, A.; Masten Rutar, J.; Ogrinc, N.; Dušak, L.; Ferjančič, B.; et al. Fermented Biomass of Arthrospira platensis as a Potential Food Ingredient. Antioxidants 2022, 11, 216. [Google Scholar] [CrossRef] [PubMed]
- Sahin, B.; Hosoglu, M.I.; Guneser, O.; Karagul-Yuceer, Y. Fermented Spirulina Products with Saccharomyces and Non- Saccharomyces Yeasts: Special Reference to Their Microbial, Physico-Chemical and Sensory Characterizations. Food Biosci. 2022, 47, 101691. [Google Scholar] [CrossRef]
- Kurt, H.; Hosoglu, M.I.; Guneser, O.; Karagul-Yuceer, Y. Influence of Different Bacteria Species in Chemical Composition and Sensory Properties of Fermented Spirulina. Food Chem. 2023, 400, 133994. [Google Scholar] [CrossRef]
- Uchida, M.; Miyoshi, T. Algal Fermentation—The Seed for a New Fermentation Industry of Foods and Related Products. Jpn. Agric. Res. Q. 2013, 47, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Kavimandan, A. Incorporation of Spirulina platensis into Probiotic Fermented Dairy Products. Int. J. Dairy Sci. 2014, 10, 1–11. [Google Scholar] [CrossRef]
- Hernández, H.; Nunes, M.C.; Prista, C.; Raymundo, A. Innovative and Healthier Dairy Products through the Addition of Microalgae: A Review. Foods 2022, 11, 755. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Osimani, A.; Milanović, V.; Aquilanti, L.; De Filippis, F.; Stellato, G.; Di Mauro, S.; Turchetti, B.; Buzzini, P.; Ercolini, D.; et al. Bacteria and Yeast Microbiota in Milk Kefir Grains from Different Italian Regions. Food Microbiol. 2015, 49, 123–133. [Google Scholar] [CrossRef]
- Garofalo, C.; Ferrocino, I.; Reale, A.; Sabbatini, R.; Milanović, V.; Alkić-Subašić, M.; Boscaino, F.; Aquilanti, L.; Pasquini, M.; Trombetta, M.F.; et al. Study of Kefir Drinks Produced by Backslopping Method Using Kefir Grains from Bosnia and Herzegovina: Microbial Dynamics and Volatilome Profile. Food Res. Int. 2020, 137, 109369. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The Promising Future of Microalgae: Current Status, Challenges, and Optimization of a Sustainable and Renewable Industry for Biofuels, Feed, and Other Products. Microb. Cell Factories 2018, 17, 36. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garofalo, C.; Norici, A.; Mollo, L.; Osimani, A.; Aquilanti, L. Fermentation of Microalgal Biomass for Innovative Food Production. Microorganisms 2022, 10, 2069. https://doi.org/10.3390/microorganisms10102069
Garofalo C, Norici A, Mollo L, Osimani A, Aquilanti L. Fermentation of Microalgal Biomass for Innovative Food Production. Microorganisms. 2022; 10(10):2069. https://doi.org/10.3390/microorganisms10102069
Chicago/Turabian StyleGarofalo, Cristiana, Alessandra Norici, Lorenzo Mollo, Andrea Osimani, and Lucia Aquilanti. 2022. "Fermentation of Microalgal Biomass for Innovative Food Production" Microorganisms 10, no. 10: 2069. https://doi.org/10.3390/microorganisms10102069