
Microbial Effectors: Key Determinants in Plant Health and Disease

 , ,
, ,
Abstract
:1. Introduction
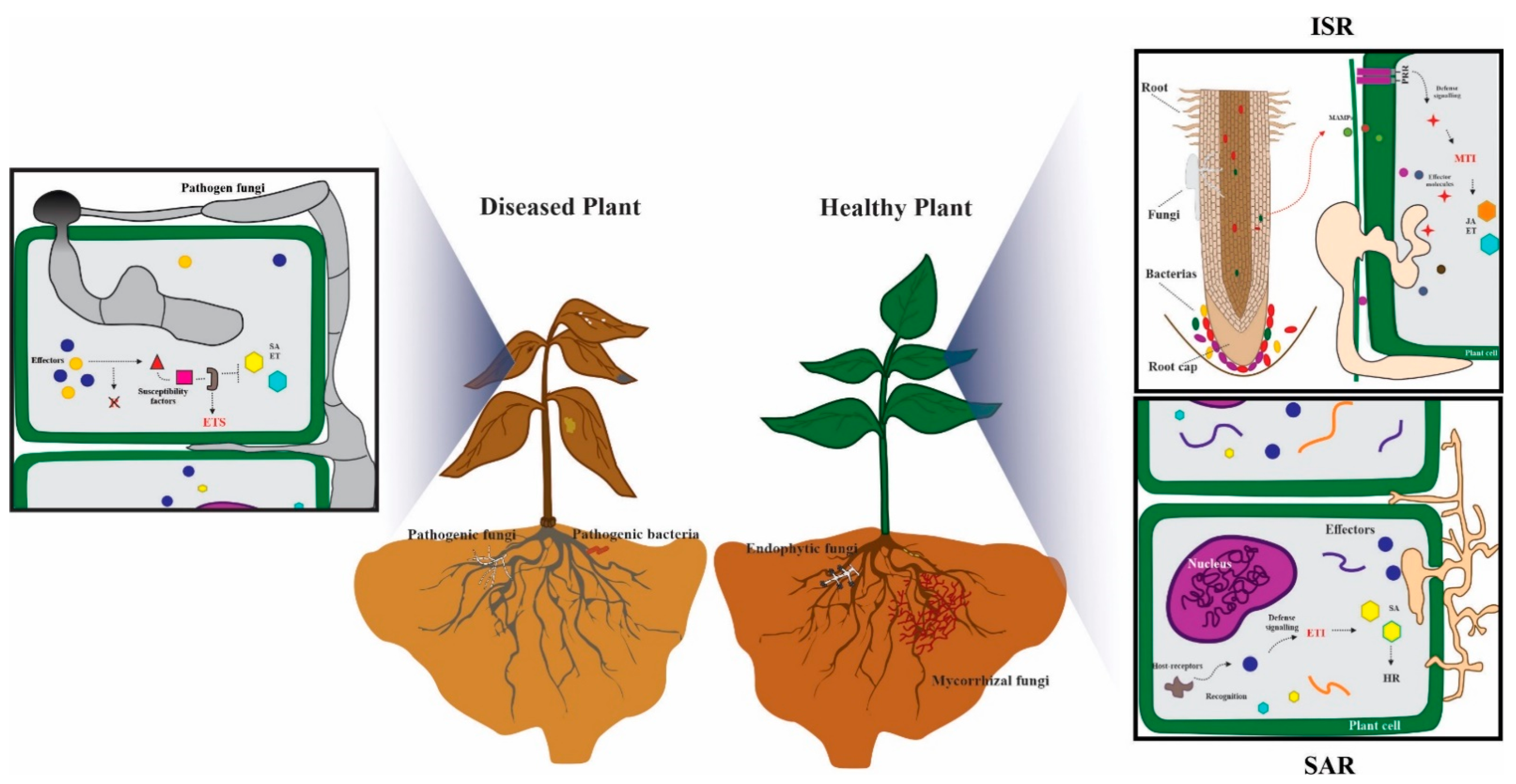
2. Effectors and Plant Defense
2.1. Effectors in Plant–Pathogen Interactions
2.2. Fungal and Oomycete Effectors
2.3. Bacterial Effectors
2.4. Effectors in Plant-Beneficial Microbe Interactions

{kind=link}
{kind=link}
| Beneficial Organism | Effector | Associated Plant | Function | References |
|---|---|---|---|---|
| Mycorrhizae | ||||
| Laccaria bicolor | MiSSP7 | Populus trichocarpa Populus tremula × Populus alba | Interacts with host plant JA signaling repressors to suppress JA-related host defense signaling | [158] |
| Laccaria bicolor | MiSSP7.6 | Populus tremula x Populus alba | Interacts with two host transcription factors: PtTrihelix1 and PtTrihelix2; involved in the establishment of Hartig net | [195] |
| Laccaria bicolor | MiSSP8 | Populus tremula x Populus alba | Involved in mantle formation and Hartig net development for the establishment of symbiosis with host | [196] |
| Glomus intraradices | SP7 | Medicago truncatula | Interacts with the host transcription factor ERF19 involved in ethylene-related defense signaling to suppress host defense | [157] |
| Rhizophagus irregularis | RiCRN1 | Medicago truncatula | Localizes to plant nucleus; involved in arbuscule development | [159] |
| Rhizophagus irregularis | RiSLM | Medicago truncatula | Binds chitin and protects against hydrolysis by chitinases. Interferes with host chitin-triggered immunity to suppress defense response | [197] |
| Pisolithus albus | PaMiSSP10b | Eucalyptus grandis | Interacts with an S-adenosyl methionine decarboxylase (AdoMetDC) in the polyamine pathway; alters polyamine biosynthesis to aid colonization | [198] |
| Rhizophagus irregularis | RiNLE1 | Medicago truncatula | Interacts with the host histone 2B protein (H2B) impairing its mono-ubiquitination which suppresses host defense-related gene expression | [160] |
| Endophytes | ||||
| Bradryhizobium elkanii USDA61 | Bel2-5 | Glycine max | Cysteine protease; involved in root nodulation | [199] |
| Rhizobium sp. NGR234 | NopM | Lablab purpureus | E3 ubiquitin ligase; promotes root nodulation | [200] |
| Rhizobium sp. NGR234 | NopE | Glycine max, Macroptilium atropurpureum and Vigna radiata | Calcium binding protein; regulates host root nodulation | [201] |
| Serendipita indica | FGB1 | Hordeum vulgare, Nicotiana benthamiana, Arabidopsis thaliana | β-glucan binding lectin; alters fungal cell wall composition and suppresses β-glucan-triggered plant immunity | [165] |
| Serendipita indica | Dld1 | Hordeum vulgare | Fungal metal ion homeostasis and micronutrient acquisition; antioxidant; enhances host root colonization | [202] |
| Trichoderma asperellum | TasXyn29.4 and TasXyn24.2 | Populus davidiana × P. alba var. pyramidalis | Xylanases; induced Me-JA accumulation. ISR against A. alternata, R. solani, and F. oxysporum | [203] |
| T. harzianum Th22 | Thph1 and Thph2 | Maize (Inbred line Huangzao 4) | Cellulases; triggered production of (ROS) and induced genes related to the jasmonate/ethylene signaling pathway. ISR against C. lunata. | [173] |
| T. atroviride IMI 206040 | Epl1 | Solanum lycopersicum | Ceratoplatanin family protein; induced the expression of a host peroxidase. ISR against A. solani and B. cinerea | [204] |
| T. virens Gv29-8 | Sm1 | Gossypium hirsutum | Ceratoplatanin family protein; triggered production of ROS and induces the expression of host defense-related genes. ISR against Colletotrichum sp. | [205] |
2.5. The New Age of Effector Identification and Characterization
3. Conclusions and Perspectives
- (a)
- Bottlenecks still exist in effector identification; effectors of plant-beneficial organisms as well as those pathogenic effectors which do not possess all the canonical effector characteristics (small size, high cysteine content, etc.) may not be well represented in in silico deduced effectoromes. Newer pipelines should take these limitations into consideration, looking beyond the common physicochemical protein characteristics of effectors currently used.
- (b)
- Effector identification is occurring at a rapid pace, but characterization is lagging relative to the large amount of effector candidates identified per organism. It is necessary to propose novel strategies and, if possible, establish standardized means of prioritizing candidates for further characterization.
- (c)
- More attention should be placed on the effectors of plant-beneficial organisms and their characterization. This can foster effector-based screening and selection of better strains of biological control organisms for their implementation in the agricultural sector. Furthermore, the isolation and application of novel effectors from pathogens, as well as plant-beneficial organisms, may prove viable in plant protection strategies.
Author Contributions
Funding
Conflicts of Interest
References
- Kamoun, S. A Catalogue of the Effector Secretome of Plant Pathogenic Oomycetes. Annu. Rev. Phytopathol. 2006, 44, 41–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogenhout, S.A.; Van der Hoorn, R.A.L.; Terauchi, R.; Kamoun, S. Emerging Concepts in Effector Biology of Plant-Associated Organisms. Mol. Plant-Microbe Interact. 2009, 22, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Xu, L.; Jia, Q.; Pan, R.; Oelmüller, R.; Zhang, W.; Wu, C. Arms Race: Diverse Effector Proteins with Conserved Motifs. Plant Signal. Behav. 2019, 14, 1557008. [Google Scholar] [CrossRef]
- Rocafort, M.; Fudal, I.; Mesarich, C.H. Apoplastic Effector Proteins of Plant-Associated Fungi and Oomycetes. Curr. Opin. Plant Biol. 2020, 56, 9–19. [Google Scholar] [CrossRef]
- Schreiber, K.J.; Chau-Ly, I.J.; Lewis, J.D. What the Wild Things Do: Mechanisms of Plant Host Manipulation by Bacterial Type III-Secreted Effector Proteins. Microorganisms 2021, 9, 1029. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; Collemare, J.; Mehrabi, R.; De Wit, P.J.G.M. Phytotoxic Secondary Metabolites and Peptides Produced by Plant Pathogenic Dothideomycete Fungi. FEMS Microbiol. Rev. 2013, 37, 67–93. [Google Scholar] [CrossRef] [Green Version]
- Ditengou, F.A.; Müller, A.; Rosenkranz, M.; Felten, J.; Lasok, H.; van Doorn, M.M.; Legué, V.; Palme, K.; Schnitzler, J.-P.; Polle, A. Volatile Signalling by Sesquiterpenes from Ectomycorrhizal Fungi Reprogrammes Root Architecture. Nat. Commun. 2015, 6, 6279. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.; Gupta, S.K.; Swapnil, P.; Zehra, A.; Dubey, M.K.; Upadhyay, R.S. Alternaria Toxins: Potential Virulence Factors and Genes Related to Pathogenesis. Front. Microbiol. 2017, 8, 1451. [Google Scholar] [CrossRef] [Green Version]
- Weiberg, A.; Wang, M.; Lin, F.-M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.-D.; Jin, H. Fungal Small RNAs Suppress Plant Immunity by Hijacking Host RNA Interference Pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Weiberg, A.; Dellota, E.; Yamane, D.; Jin, H. Botrytis Small RNA Bc-SiR37 Suppresses Plant Defense Genes by Cross-Kingdom RNAi. RNA Biol. 2017, 14, 421–428. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, C.; Wang, G.; Li, Y.; Wang, S. Insects Defend against Fungal Infection by Employing MicroRNAs to Silence Virulence-Related Genes. Proc. Natl. Acad. Sci. USA 2021, 118, e2023802118. [Google Scholar] [CrossRef] [PubMed]
- Selin, C.; de Kievit, T.R.; Belmonte, M.F.; Fernando, W.G.D. Elucidating the Role of Effectors in Plant-Fungal Interactions: Progress and Challenges. Front. Microbiol. 2016, 7, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Seto, D.; Subramaniam, R.; Desveaux, D. Oh, the Places They’ll Go! A Survey of Phytopathogen Effectors and Their Host Targets. Plant J. 2018, 93, 651–663. [Google Scholar] [CrossRef]
- Büttner, D. Behind the Lines-Actions of Bacterial Type III Effector Proteins in Plant Cells. FEMS Microbiol. Rev. 2016, 40, 894–937. [Google Scholar] [CrossRef] [Green Version]
- Uhse, S.; Djamei, A. Effectors of Plant-Colonizing Fungi and Beyond. PLoS Pathog. 2018, 14, e1006992. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Zhang, W.; Lin, M.; Teymournejad, O.; Budachetri, K.; Lakritz, J.; Rikihisa, Y. Iron Robbery by Intracellular Pathogen via Bacterial Effector–Induced Ferritinophagy. Proc. Natl. Acad. Sci. USA 2021, 118, e2026598118. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Si, J.; Han, Z.; Chen, D. Action Mechanisms of Effectors in Plant-Pathogen Interaction. Int. J. Mol. Sci. 2022, 23, 6758. [Google Scholar] [CrossRef]
- Figueroa, M.; Ortiz, D.; Henningsen, E.C. Tactics of Host Manipulation by Intracellular Effectors from Plant Pathogenic Fungi. Curr. Opin. Plant Biol. 2021, 62, 102054. [Google Scholar] [CrossRef]
- Rodriguez, P.A.; Escudero-Martinez, C.; Bos, J.I.B. An Aphid Effector Targets Trafficking Protein VPS52 in a Host-Specific Manner to Promote Virulence. Plant Physiol. 2017, 173, 1892–1903. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, X.; Fu, Y.; Crespo-Herrera, L.; Liu, H.; Wang, Q.; Zhang, Y.; Chen, J. Salivary Effector Sm9723 of Grain Aphid Sitobion miscanthi Suppresses Plant Defense and Is Essential for Aphid Survival on Wheat. Int. J. Mol. Sci. 2022, 23, 6909. [Google Scholar] [CrossRef]
- Leelarasamee, N.; Zhang, L.; Gleason, C. The Root-Knot Nematode Effector MiPFN3 Disrupts Plant Actin Filaments and Promotes Parasitism. PLoS Pathog. 2018, 14, e1006947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kud, J.; Wang, W.; Gross, R.; Fan, Y.; Huang, L.; Yuan, Y.; Gray, A.; Duarte, A.; Kuhl, J.C.; Caplan, A.; et al. The Potato Cyst Nematode Effector RHA1B Is a Ubiquitin Ligase and Uses Two Distinct Mechanisms to Suppress Plant Immune Signaling. PLoS Pathog. 2019, 15, e1007720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.-J.; Wu, X.-Q.; Li, H.-Y.; Zhao, Q.; Wang, Y.-C.; Ye, J.-R. An Effector, BxSapB1, Induces Cell Death and Contributes to Virulence in the Pine Wood Nematode Bursaphelenchus xylophilus. Mol. Plant-Microbe Interact. 2018, 32, 452–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, P.; Leary, A.Y.; Tumtas, Y.; Savage, Z.; Dagvadorj, B.; Duggan, C.; Yuen, E.L.; Sanguankiattichai, N.; Tan, E.; Khandare, V.; et al. An Oomycete Effector Subverts Host Vesicle Trafficking to Channel Starvation-Induced Autophagy to the Pathogen Interface. eLife 2021, 10, e65285. [Google Scholar] [CrossRef]
- Zhu, C.; Liu, J.-H.; Zhao, J.-H.; Liu, T.; Chen, Y.-Y.; Wang, C.-H.; Zhang, Z.-H.; Guo, H.-S.; Duan, C.-G. A Fungal Effector Suppresses the Nuclear Export of AGO1–MiRNA Complex to Promote Infection in Plants. Proc. Natl. Acad. Sci. USA 2022, 119, e2114583119. [Google Scholar] [CrossRef]
- He, Q.; McLellan, H.; Boevink, P.C.; Birch, P.R.J. All Roads Lead to Susceptibility: The Many Modes of Action of Fungal and Oomycete Intracellular Effectors. Plant Commun. 2020, 1, 100050. [Google Scholar] [CrossRef]
- Choi, S.; Jayaraman, J.; Segonzac, C.; Park, H.-J.; Park, H.; Han, S.-W.; Sohn, K.H. Pseudomonas syringae pv. actinidiae Type III Effectors Localized at Multiple Cellular Compartments Activate or Suppress Innate Immune Responses in Nicotiana Benthamiana. Front. Plant Sci. 2017, 8, 2157. [Google Scholar] [CrossRef]
- Rufián, J.S.; Rueda-Blanco, J.; López-Márquez, D.; Macho, A.P.; Beuzón, C.R.; Ruiz-Albert, J. The Bacterial Effector HopZ1a Acetylates MKK7 to Suppress Plant Immunity. New Phytol. 2021, 231, 1138–1156. [Google Scholar] [CrossRef]
- Zvereva, A.S.; Golyaev, V.; Turco, S.; Gubaeva, E.G.; Rajeswaran, R.; Schepetilnikov, M.V.; Srour, O.; Ryabova, L.A.; Boller, T.; Pooggin, M.M. Viral Protein Suppresses Oxidative Burst and Salicylic Acid-Dependent Autophagy and Facilitates Bacterial Growth on Virus-Infected Plants. New Phytol. 2016, 211, 1020–1034. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Qi, T.; Li, W.-X.; Tian, H.; Gao, H.; Wang, J.; Ge, J.; Yao, R.; Ren, C.; Wang, X.-B.; et al. Viral Effector Protein Manipulates Host Hormone Signaling to Attract Insect Vectors. Cell Res. 2017, 27, 402–415. [Google Scholar] [CrossRef]
- Plett, J.M.; Yin, H.; Mewalal, R.; Hu, R.; Li, T.; Ranjan, P.; Jawdy, S.; Paoli, H.C.D.; Butler, G.; Burch-Smith, T.M.; et al. Populus trichocarpa Encodes Small, Effector-like Secreted Proteins That Are Highly Induced during Mutualistic Symbiosis. Sci. Rep. 2017, 7, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toruño, T.Y.; Stergiopoulos, I.; Coaker, G. Plant-Pathogen Effectors: Cellular Probes Interfering with Plant Defenses in Spatial and Temporal Manners. Annu. Rev. Phytopathol. 2016, 54, 419–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, O.B.; dos Garruti, D.S.; Norões, N.P.; de Silva, S.O.; Weber, O.B.; dos Garruti, D.S.; Norões, N.P.; de Silva, S.O. Performance of Banana Genotypes with Resistance to Black Leaf Streak Disease in Northeastern Brazil. Pesqui. Agropecuária Bras. 2017, 52, 161–169. [Google Scholar] [CrossRef]
- Han, X.; Kahmann, R. Manipulation of Phytohormone Pathways by Effectors of Filamentous Plant Pathogens. Front. Plant Sci. 2019, 10, 822. [Google Scholar] [CrossRef] [Green Version]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal Effectors and Plant Susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.A.; McDowell, J.M. Recent Advances in Understanding of Fungal and Oomycete Effectors. Curr. Opin. Plant Biol. 2022, 68, 102228. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Kahmann, R. Cell Wall–Associated Effectors of Plant-Colonizing Fungi. Mycologia 2021, 113, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ding, Y.; Cai, B.; Qin, X.; Wu, J.; Yuan, M.; Wan, S.; Zhao, Y.; Xin, X.-F. Bacterial Effectors Manipulate Plant Abscisic Acid Signaling for Creation of an Aqueous Apoplast. Cell Host Microbe 2022, 30, 518–529.e6. [Google Scholar] [CrossRef]
- Robin, G.P.; Kleemann, J.; Neumann, U.; Cabre, L.; Dallery, J.-F.; Lapalu, N.; O’Connell, R.J. Subcellular Localization Screening of Colletotrichum higginsianum Effector Candidates Identifies Fungal Proteins Targeted to Plant Peroxisomes, Golgi Bodies, and Microtubules. Front. Plant Sci. 2018, 9, 562. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Xiao, K.; Pan, H.; Liu, J. Chloroplast: The Emerging Battlefield in Plant–Microbe Interactions. Front. Plant Sci. 2021, 12, 637853. [Google Scholar] [CrossRef]
- Fabro, G. Oomycete Intracellular Effectors: Specialised Weapons Targeting Strategic Plant Processes. New Phytol. 2022, 233, 1074–1082. [Google Scholar] [CrossRef] [PubMed]
- van Schie, C.C.N.; Takken, F.L.W. Susceptibility Genes 101: How to Be a Good Host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef] [PubMed]
- Shao, D.; Smith, D.L.; Kabbage, M.; Roth, M.G. Effectors of Plant Necrotrophic Fungi. Front. Plant Sci. 2021, 12, 687713. [Google Scholar] [CrossRef] [PubMed]
- Murphy, F.; He, Q.; Armstrong, M.; Giuliani, L.M.; Boevink, P.C.; Zhang, W.; Tian, Z.; Birch, P.R.J.; Gilroy, E.M. The Potato MAP3K StVIK Is Required for the Phytophthora infestans RXLR Effector Pi17316 to Promote Disease. Plant Physiol. 2018, 177, 398–410. [Google Scholar] [CrossRef] [Green Version]
- Lan, X.; Liu, Y.; Song, S.; Yin, L.; Xiang, J.; Qu, J.; Lu, J. Plasmopara viticola Effector PvRXLR131 Suppresses Plant Immunity by Targeting Plant Receptor-like Kinase Inhibitor BKI1. Mol. Plant Pathol. 2019, 20, 765–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C. The Intricate Dance between Ustilago Effector and Maize Defense. Plant Cell 2022, 34, 2586–2587. [Google Scholar] [CrossRef]
- Ma, L.; Djavaheri, M.; Wang, H.; Larkan, N.J.; Haddadi, P.; Beynon, E.; Gropp, G.; Borhan, M.H. Leptosphaeria maculans Effector Protein AvrLm1 Modulates Plant Immunity by Enhancing MAP Kinase 9 Phosphorylation. iScience 2018, 3, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Huai, B.; Lu, Y.; Cai, K.; Guo, J.; Zhu, X.; Kang, Z.; Guo, J. A Stripe Rust Effector Pst18363 Targets and Stabilises TaNUDX23 That Promotes Stripe Rust Disease. New Phytol. 2020, 225, 880–895. [Google Scholar] [CrossRef]
- Vleeshouwers, V.G.A.A.; Rietman, H.; Krenek, P.; Champouret, N.; Young, C.; Oh, S.-K.; Wang, M.; Bouwmeester, K.; Vosman, B.; Visser, R.G.F.; et al. Effector Genomics Accelerates Discovery and Functional Profiling of Potato Disease Resistance and Phytophthora infestans Avirulence Genes. PLoS ONE 2008, 3, e2875. [Google Scholar] [CrossRef] [PubMed]
- Rietman, H.; Bijsterbosch, G.; Cano, L.M.; Lee, H.-R.; Vossen, J.H.; Jacobsen, E.; Visser, R.G.F.; Kamoun, S.; Vleeshouwers, V.G.A.A. Qualitative and Quantitative Late Blight Resistance in the Potato Cultivar Sarpo Mira Is Determined by the Perception of Five Distinct RXLR Effectors. Mol. Plant-Microbe Interact. 2012, 25, 910–919. [Google Scholar] [CrossRef]
- Timilsina, S.; Abrahamian, P.; Potnis, N.; Minsavage, G.V.; White, F.F.; Staskawicz, B.J.; Jones, J.B.; Vallad, G.E.; Goss, E.M. Analysis of Sequenced Genomes of Xanthomonas perforans Identifies Candidate Targets for Resistance Breeding in Tomato. Phytopathology 2016, 106, 1097–1104. [Google Scholar] [CrossRef] [Green Version]
- Giesbers, A.K.J.; Pelgrom, A.J.E.; Visser, R.G.F.; Niks, R.E.; Van den Ackerveken, G.; Jeuken, M.J.W. Effector-Mediated Discovery of a Novel Resistance Gene against Bremia lactucae in a Nonhost Lettuce Species. New Phytol. 2017, 216, 915–926. [Google Scholar] [CrossRef] [Green Version]
- Vleeshouwers, V.G.A.A.; Oliver, R.P. Effectors as Tools in Disease Resistance Breeding against Biotrophic, Hemibiotrophic, and Necrotrophic Plant Pathogens. Mol. Plant-Microbe Interact. 2014, 27, 196–206. [Google Scholar] [CrossRef] [Green Version]
- See, P.T.; Iagallo, E.M.; Oliver, R.P.; Moffat, C.S. Heterologous Expression of the Pyrenophora tritici-repentis Effector Proteins ToxA and ToxB, and the Prevalence of Effector Sensitivity in Australian Cereal Crops. Front. Microbiol. 2019, 10, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gust, A.A.; Brunner, F.; Nürnberger, T. Biotechnological Concepts for Improving Plant Innate Immunity. Curr. Opin. Biotechnol. 2010, 21, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Pavan, S.; Jacobsen, E.; Visser, R.G.F.; Bai, Y. Loss of Susceptibility as a Novel Breeding Strategy for Durable and Broad-Spectrum Resistance. Mol. Breed. 2010, 25, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Ruiz, H.; Szurek, B.; Van den Ackerveken, G. Stop Helping Pathogens: Engineering Plant Susceptibility Genes for Durable Resistance. Curr. Opin. Biotechnol. 2021, 70, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xu, X.; Gong, Q.; Li, Z.; Li, Y.; Wang, S.; Yang, Y.; Ma, W.; Liu, L.; Zhu, B.; et al. Engineering Broad-Spectrum Bacterial Blight Resistance by Simultaneously Disrupting Variable TALE-Binding Elements of Multiple Susceptibility Genes in Rice. Mol. Plant 2019, 12, 1434–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iida, Y.; van ‘t Hof, P.; Beenen, H.; Mesarich, C.; Kubota, M.; Stergiopoulos, I.; Mehrabi, R.; Notsu, A.; Fujiwara, K.; Bahkali, A.; et al. Novel Mutations Detected in Avirulence Genes Overcoming Tomato Cf Resistance Genes in Isolates of a Japanese Population of Cladosporium fulvum. PLoS ONE 2015, 10, e0123271. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Peng, G.; Kutcher, H.R.; Balesdent, M.-H.; Delourme, R.; Fernando, W.G.D. Breakdown of Rlm3 Resistance in the Brassica napus–Leptosphaeria maculans Pathosystem in Western Canada. Eur. J. Plant Pathol. 2016, 145, 659–674. [Google Scholar] [CrossRef]
- Yan, H.; Nelson, B. Adaptation of Phytophthora sojae to Rps Resistance Genes over the Past Two Decades in North Dakota. Plant Health Prog. 2019, 20, 88–93. [Google Scholar] [CrossRef]
- Ivanov, A.A.; Ukladov, E.O.; Golubeva, T.S. Phytophthora infestans: An Overview of Methods and Attempts to Combat Late Blight. J. Fungi 2021, 7, 1071. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodds, P.N.; Rathjen, J.P. Plant Immunity: Towards an Integrated View of Plant–Pathogen Interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Boyd, L.A.; Ridout, C.; O’Sullivan, D.M.; Leach, J.E.; Leung, H. Plant–Pathogen Interactions: Disease Resistance in Modern Agriculture. Trends Genet. 2013, 29, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Ngou, B.P.M.; Ahn, H.-K.; Ding, P.; Jones, J.D.G. Mutual Potentiation of Plant Immunity by Cell-Surface and Intracellular Receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef]
- Alhoraibi, H.; Bigeard, J.; Rayapuram, N.; Colcombet, J.; Hirt, H. Plant Immunity: The MTI-ETI Model and Beyond. Curr. Issues Mol. Biol. 2019, 30, 39–58. [Google Scholar] [CrossRef]
- Hemetsberger, C.; Mueller, A.N.; Matei, A.; Herrberger, C.; Hensel, G.; Kumlehn, J.; Mishra, B.; Sharma, R.; Thines, M.; Hückelhoven, R.; et al. The Fungal Core Effector Pep1 Is Conserved across Smuts of Dicots and Monocots. New Phytol. 2015, 206, 1116–1126. [Google Scholar] [CrossRef] [PubMed]
- Irieda, H.; Inoue, Y.; Mori, M.; Yamada, K.; Oshikawa, Y.; Saitoh, H.; Uemura, A.; Terauchi, R.; Kitakura, S.; Kosaka, A.; et al. Conserved Fungal Effector Suppresses PAMP-Triggered Immunity by Targeting Plant Immune Kinases. Proc. Natl. Acad. Sci. USA 2019, 116, 496–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flor, H.H. Inheritance of Pathogenicity in Melampsora lini. Phytopathology 1942, 32, 653–669. [Google Scholar]
- Eitas, T.K.; Dangl, J.L. NB-LRR Proteins: Pairs, Pieces, Perception, Partners and Pathways. Curr. Opin. Plant Biol. 2010, 13, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Liu, J.; Guo, Q.; Pan, L.; Chai, S.; Cheng, Y.; Ruan, M.; Ye, Q.; Wang, R.; Yao, Z.; et al. Genomic Organization and Comparative Phylogenic Analysis of NBS-LRR Resistance Gene Family in Solanum pimpinellifolium and Arabidopsis thaliana. Evol. Bioinform. 2020, 16, 1176934320911055. [Google Scholar] [CrossRef] [Green Version]
- Win, J.; Chaparro-Garcia, A.; Belhaj, K.; Saunders, D.G.O.; Yoshida, K.; Dong, S.; Schornack, S.; Zipfel, C.; Robatzek, S.; Hogenhout, S.A.; et al. Effector Biology of Plant-Associated Organisms: Concepts and Perspectives. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 235–247. [Google Scholar] [CrossRef]
- Bent, A.F.; Mackey, D. Elicitors, Effectors, and R Genes: The New Paradigm and a Lifetime Supply of Questions. Annu. Rev. Phytopathol. 2007, 45, 399–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-Microbe Interactions: Shaping the Evolution of the Plant Immune Response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.-M.; He, S.Y.; Xin, X.-F. Pattern-Recognition Receptors Are Required for NLR-Mediated Plant Immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Derevnina, L.; Petre, B.; Kellner, R.; Dagdas, Y.F.; Sarowar, M.N.; Giannakopoulou, A.; De la Concepcion, J.C.; Chaparro-Garcia, A.; Pennington, H.G.; van West, P.; et al. Emerging Oomycete Threats to Plants and Animals. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150459. [Google Scholar] [CrossRef] [Green Version]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The Still Underestimated Problem of Fungal Diseases Worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodds, P.N.; Rafiqi, M.; Gan, P.H.P.; Hardham, A.R.; Jones, D.A.; Ellis, J.G. Effectors of Biotrophic Fungi and Oomycetes: Pathogenicity Factors and Triggers of Host Resistance. New Phytol. 2009, 183, 993–1000. [Google Scholar] [CrossRef]
- Tintor, N.; Nieuweboer, G.A.M.; Bakker, I.A.W.; Takken, F.L.W. The Intracellularly Acting Effector Foa3 Suppresses Defense Responses When Infiltrated into the Apoplast. Front. Plant Sci. 2022, 13, 1628. [Google Scholar] [CrossRef] [PubMed]
- Saado, I.; Chia, K.-S.; Betz, R.; Alcântara, A.; Pettkó-Szandtner, A.; Navarrete, F.; D’Auria, J.C.; Kolomiets, M.V.; Melzer, M.; Feussner, I.; et al. Effector-Mediated Relocalization of a Maize Lipoxygenase Protein Triggers Susceptibility to Ustilago maydis. Plant Cell 2022, 34, 2785–2805. [Google Scholar] [CrossRef] [PubMed]
- An, B.; Wang, W.; Guo, Y.; Wang, Q.; Luo, H.; He, C. BAS2 Is Required for Conidiation and Pathogenicity of Colletotrichum gloeosporioides from Hevea brasiliensis. Int. J. Mol. Sci. 2018, 19, 1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukada, F.; Rössel, N.; Münch, K.; Glatter, T.; Kahmann, R. A Small Ustilago maydis Effector Acts as a Novel Adhesin for Hyphal Aggregation in Plant Tumors. New Phytol. 2021, 231, 416–431. [Google Scholar] [CrossRef]
- Rodriguez-Moreno, L.; Ebert, M.K.; Bolton, M.D.; Thomma, B.P.H.J. Tools of the Crook- Infection Strategies of Fungal Plant Pathogens. Plant J. Cell Mol. Biol. 2018, 93, 664–674. [Google Scholar] [CrossRef] [Green Version]
- Carreón-Anguiano, K.G.; Islas-Flores, I.; Vega-Arreguín, J.; Sáenz-Carbonell, L.; Canto-Canché, B. EffHunter: A Tool for Prediction of Effector Protein Candidates in Fungal Proteomic Databases. Biomolecules 2020, 10, 712. [Google Scholar] [CrossRef]
- Duplessis, S.; Cuomo, C.A.; Lin, Y.-C.; Aerts, A.; Tisserant, E.; Veneault-Fourrey, C.; Joly, D.L.; Hacquard, S.; Amselem, J.; Cantarel, B.L.; et al. Obligate Biotrophy Features Unraveled by the Genomic Analysis of Rust Fungi. Proc. Natl. Acad. Sci. USA 2011, 108, 9166–9171. [Google Scholar] [CrossRef] [Green Version]
- Nemri, A.; Saunders, D.G.O.; Anderson, C.; Upadhyaya, N.M.; Win, J.; Lawrence, G.J.; Jones, D.A.; Kamoun, S.; Ellis, J.G.; Dodds, P.N. The Genome Sequence and Effector Complement of the Flax Rust Pathogen Melampsora Lini. Front. Plant Sci. 2014, 5, 98. [Google Scholar] [CrossRef]
- Faris, J.D.; Friesen, T.L. Plant Genes Hijacked by Necrotrophic Fungal Pathogens. Curr. Opin. Plant Biol. 2020, 56, 74–80. [Google Scholar] [CrossRef]
- Tada, Y.; Kusaka, K.; Betsuyaku, S.; Shinogi, T.; Sakamoto, M.; Ohura, Y.; Hata, S.; Mori, T.; Tosa, Y.; Mayama, S. Victorin Triggers Programmed Cell Death and the Defense Response via Interaction with a Cell Surface Mediator. Plant Cell Physiol. 2005, 46, 1787–1798. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Gao, Y.; Kim, Y.M.; Faris, J.D.; Shelver, W.L.; de Wit, P.J.G.M.; Xu, S.S.; Friesen, T.L. SnTox1, a Parastagonospora nodorum Necrotrophic Effector, Is a Dual-Function Protein That Facilitates Infection While Protecting from Wheat-Produced Chitinases. New Phytol. 2016, 211, 1052–1064. [Google Scholar] [CrossRef]
- van Esse, H.P.; Bolton, M.D.; Stergiopoulos, I.; de Wit, P.J.G.M.; Thomma, B.P.H.J. The Chitin-Binding Cladosporium fulvum Effector Protein Avr4 Is a Virulence Factor. Mol. Plant. Microbe Interact. 2007, 20, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; van den Burg, H.A.; Ökmen, B.; Beenen, H.G.; van Liere, S.; Kema, G.H.J.; de Wit, P.J.G.M. Tomato Cf Resistance Proteins Mediate Recognition of Cognate Homologous Effectors from Fungi Pathogenic on Dicots and Monocots. Proc. Natl. Acad. Sci. USA 2010, 107, 7610–7615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerink, N.; Brandwagt, B.F.; de Wit, P.J.G.M.; Joosten, M.H.A.J. Cladosporium fulvum Circumvents the Second Functional Resistance Gene Homologue at the Cf-4 Locus (Hcr9-4E) by Secretion of a Stable Avr4E Isoform. Mol. Microbiol. 2004, 54, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Bolton, M.D.; Esse, H.P.V.; Vossen, J.H.; Jonge, R.D.; Stergiopoulos, I.; Stulemeijer, I.J.E.; Berg, G.C.M.V.D.; Borrás-Hidalgo, O.; Dekker, H.L.; Koster, C.G.D.; et al. The Novel Cladosporium fulvum Lysin Motif Effector Ecp6 Is a Virulence Factor with Orthologues in Other Fungal Species. Mol. Microbiol. 2008, 69, 119–136. [Google Scholar] [CrossRef]
- de Jonge, R.; van Esse, H.P.; Kombrink, A.; Shinya, T.; Desaki, Y.; Bours, R.; van der Krol, S.; Shibuya, N.; Joosten, M.H.A.J.; Thomma, B.P.H.J. Conserved Fungal LysM Effector Ecp6 Prevents Chitin-Triggered Immunity in Plants. Science 2010, 329, 953–955. [Google Scholar] [CrossRef]
- Ve, T.; Williams, S.J.; Catanzariti, A.-M.; Rafiqi, M.; Rahman, M.; Ellis, J.G.; Hardham, A.R.; Jones, D.A.; Anderson, P.A.; Dodds, P.N.; et al. Structures of the Flax-Rust Effector AvrM Reveal Insights into the Molecular Basis of Plant-Cell Entry and Effector-Triggered Immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 17594–17599. [Google Scholar] [CrossRef] [Green Version]
- Cesari, S.; Thilliez, G.; Ribot, C.; Chalvon, V.; Michel, C.; Jauneau, A.; Rivas, S.; Alaux, L.; Kanzaki, H.; Okuyama, Y.; et al. The Rice Resistance Protein Pair RGA4/RGA5 Recognizes the Magnaporthe oryzae Effectors AVR-Pia and AVR1-CO39 by Direct Binding. Plant Cell 2013, 25, 1463–1481. [Google Scholar] [CrossRef] [Green Version]
- Kanzaki, H.; Yoshida, K.; Saitoh, H.; Fujisaki, K.; Hirabuchi, A.; Alaux, L.; Fournier, E.; Tharreau, D.; Terauchi, R. Arms Race Co-Evolution of Magnaporthe oryzae AVR-Pik and Rice Pik Genes Driven by Their Physical Interactions. Plant J. 2012, 72, 894–907. [Google Scholar] [CrossRef]
- Inoue, Y.; Vy, T.T.P.; Yoshida, K.; Asano, H.; Mitsuoka, C.; Asuke, S.; Anh, V.L.; Cumagun, C.J.R.; Chuma, I.; Terauchi, R.; et al. Evolution of the Wheat Blast Fungus through Functional Losses in a Host Specificity Determinant. Science 2017, 357, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Olave-Achury, A.; Heal, R.; Pais, M.; Witek, K.; Ahn, H.-K.; Zhao, H.; Bhanvadia, S.; Karki, H.S.; Song, T.; et al. A Potato Late Blight Resistance Gene Protects against Multiple Phytophthora Species by Recognizing a Broadly Conserved RXLR-WY Effector. Mol. Plant 2022, 15, 1457–1469. [Google Scholar] [CrossRef]
- Henares, B.M.; Debler, J.W.; Farfan-Caceres, L.M.; Grime, C.R.; Syme, R.A.; Blake, S.N.; Herdina; Davidson, J.A.; Oliver, R.P.; Singh, K.B.; et al. The Novel Avirulence Effector AlAvr1 from Ascochyta lentis Mediates Host Cultivar Specificity of Ascochyta Blight in Lentil. Mol. Plant Pathol. 2022, 23, 984–996. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Leonard, A.; Cahill, J.; Lv, M.; Li, Y.; Thatcher, S.; Li, X.; Zhao, X.; Du, W.; Li, Z.; et al. The RppC-AvrRppC NLR-Effector Interaction Mediates the Resistance to Southern Corn Rust in Maize. Mol. Plant 2022, 15, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Mukaihara, T. The Type III Effector RipB from Ralstonia solanacearum RS1000 Acts as a Major Avirulence Factor in Nicotiana Benthamiana and Other Nicotiana Species. Mol. Plant Pathol. 2019, 20, 1237–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.; Moon, H.; Choi, S.; Yoon, H.; Prokchorchik, M.; Jayaraman, J.; Sujeevan, R.; Kang, Y.M.; McCann, H.C.; Segonzac, C.; et al. Ralstonia solanacearum Type III Effector RipJ Triggers Bacterial Wilt Resistance in Solanum pimpinellifolium. Mol. Plant-Microbe Interact. 2021, 34, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Pandey, A.; Yoon, H.; Choi, S.; Jeon, H.; Prokchorchik, M.; Jung, G.; Witek, K.; Valls, M.; McCann, H.C.; et al. Identification of RipAZ1 as an Avirulence Determinant of Ralstonia solanacearum in Solanum americanum. Mol. Plant Pathol. 2021, 22, 317–333. [Google Scholar] [CrossRef]
- Kang, H.; Nguyen, Q.-M.; Iswanto, A.B.B.; Hong, J.C.; Bhattacharjee, S.; Gassmann, W.; Kim, S.H. Nuclear Localization of HopA1Pss61 Is Required for Effector-Triggered Immunity. Plants 2021, 10, 888. [Google Scholar] [CrossRef]
- He, Q.; Naqvi, S.; McLellan, H.; Boevink, P.C.; Champouret, N.; Hein, I.; Birch, P.R.J. Plant Pathogen Effector Utilizes Host Susceptibility Factor NRL1 to Degrade the Immune Regulator SWAP70. Proc. Natl. Acad. Sci. USA 2018, 115, E7834–E7843. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; McLellan, H.; Naqvi, S.; He, Q.; Boevink, P.C.; Armstrong, M.; Giuliani, L.M.; Zhang, W.; Tian, Z.; Zhan, J.; et al. Potato NPH3/RPT2-Like Protein StNRL1, Targeted by a Phytophthora infestans RXLR Effector, Is a Susceptibility Factor. Plant Physiol. 2016, 171, 645–657. [Google Scholar] [CrossRef] [Green Version]
- Boevink, P.C.; Wang, X.; McLellan, H.; He, Q.; Naqvi, S.; Armstrong, M.R.; Zhang, W.; Hein, I.; Gilroy, E.M.; Tian, Z.; et al. A Phytophthora infestans RXLR Effector Targets Plant PP1c Isoforms That Promote Late Blight Disease. Nat. Commun. 2016, 7, 10311. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, H.; Jing, M.; Zhu, J.; Guo, B.; Wang, Y.; Lin, Y.; Chen, H.; Kong, L.; Ma, Z.; et al. A Phytophthora Effector Recruits a Host Cytoplasmic Transacetylase into Nuclear Speckles to Enhance Plant Susceptibility. eLife 2018, 7, e40039. [Google Scholar] [CrossRef]
- Gilroy, E.M.; Breen, S.; Whisson, S.C.; Squires, J.; Hein, I.; Kaczmarek, M.; Turnbull, D.; Boevink, P.C.; Lokossou, A.; Cano, L.M.; et al. Presence/Absence, Differential Expression and Sequence Polymorphisms between PiAVR2 and PiAVR2-like in Phytophthora infestans Determine Virulence on R2 Plants. New Phytol. 2011, 191, 763–776. [Google Scholar] [CrossRef]
- Turnbull, D.; Wang, H.; Breen, S.; Malec, M.; Naqvi, S.; Yang, L.; Welsh, L.; Hemsley, P.; Zhendong, T.; Brunner, F.; et al. AVR2 Targets BSL Family Members, Which Act as Susceptibility Factors to Suppress Host Immunity. Plant Physiol. 2019, 180, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Faris, J.D.; Zhang, Z.; Lu, H.; Lu, S.; Reddy, L.; Cloutier, S.; Fellers, J.P.; Meinhardt, S.W.; Rasmussen, J.B.; Xu, S.S.; et al. A Unique Wheat Disease Resistance-like Gene Governs Effector-Triggered Susceptibility to Necrotrophic Pathogens. Proc. Natl. Acad. Sci. USA 2010, 107, 13544–13549. [Google Scholar] [CrossRef] [Green Version]
- See, P.T.; Marathamuthu, K.A.; Iagallo, E.M.; Oliver, R.P.; Moffat, C.S. Evaluating the Importance of the Tan Spot ToxA–Tsn1 Interaction in Australian Wheat Varieties. Plant Pathol. 2018, 67, 1066–1075. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhang, Z.; Faris, J.D.; Oliver, R.P.; Syme, R.; McDonald, M.C.; McDonald, B.A.; Solomon, P.S.; Lu, S.; Shelver, W.L.; et al. The Cysteine Rich Necrotrophic Effector SnTox1 Produced by Stagonospora Nodorum Triggers Susceptibility of Wheat Lines Harboring Snn1. PLoS Pathog. 2012, 8, e1002467. [Google Scholar] [CrossRef] [Green Version]
- Friesen, T.L.; Faris, J.D. Molecular Mapping of Resistance to Pyrenophora tritici-repentis Race 5 and Sensitivity to Ptr ToxB in Wheat. TAG Theor. Appl. Genet. Theor. Angew. Genet. 2004, 109, 464–471. [Google Scholar] [CrossRef]
- Figueroa, M.; Manning, V.A.; Pandelova, I.; Ciuffetti, L.M. Persistence of the Host-Selective Toxin Ptr ToxB in the Apoplast. Mol. Plant-Microbe Interact. 2015, 28, 1082–1090. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Zhai, Y.; Feng, L.; Karimi, H.Z.; Rutter, B.D.; Zeng, L.; Choi, D.S.; Zhang, B.; Gu, W.; Chen, X.; et al. A Phytophthora Effector Suppresses Trans-Kingdom RNAi to Promote Disease Susceptibility. Cell Host Microbe 2019, 25, 153–165.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, S.; Kumari, P.; Lapin, D.; Griebel, T.; Hickman, R.; Guo, W.; Zhang, R.; Parker, J.E.; Beynon, J.; Denby, K.; et al. Downy Mildew Effector HaRxL21 Interacts with the Transcriptional Repressor TOPLESS to Promote Pathogen Susceptibility. PLoS Pathog. 2020, 16, e1008835. [Google Scholar] [CrossRef]
- Zhu, W.; Wei, W.; Fu, Y.; Cheng, J.; Xie, J.; Li, G.; Yi, X.; Kang, Z.; Dickman, M.B.; Jiang, D. A Secretory Protein of Necrotrophic Fungus Sclerotinia sclerotiorum That Suppresses Host Resistance. PLoS ONE 2013, 8, e53901. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Yang, G.; Ma, M.; Liu, X.; Li, B.; Xie, J.; Fu, Y.; Chen, T.; Yu, Y.; Chen, W.; et al. An Effector of a Necrotrophic Fungal Pathogen Targets the Calcium-Sensing Receptor in Chloroplasts to Inhibit Host Resistance. Mol. Plant Pathol. 2020, 21, 686–701. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Tang, C.; Wang, X.; Sun, S.; Zhao, J.; Kang, Z.; Wang, X. An Effector Protein of the Wheat Stripe Rust Fungus Targets Chloroplasts and Suppresses Chloroplast Function. Nat. Commun. 2019, 10, 5571. [Google Scholar] [CrossRef] [Green Version]
- Qi, T.; Guo, J.; Liu, P.; He, F.; Wan, C.; Islam, M.A.; Tyler, B.M.; Kang, Z.; Guo, J. Stripe Rust Effector PstGSRE1 Disrupts Nuclear Localization of ROS-Promoting Transcription Factor TaLOL2 to Defeat ROS-Induced Defense in Wheat. Mol. Plant 2019, 12, 1624–1638. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Y.; Wang, Y.; Du, Y.; Song, C.; Song, P.; Yang, Q.; He, F.; Bai, X.; Huang, L.; et al. Glycine-Serine-Rich Effector PstGSRE4 in Puccinia striiformis f. sp. tritici Inhibits the Activity of Copper Zinc Superoxide Dismutase to Modulate Immunity in Wheat. PLoS Pathog. 2022, 18, e1010702. [Google Scholar] [CrossRef]
- Zheng, X.; Fang, A.; Qiu, S.; Zhao, G.; Wang, J.; Wang, S.; Wei, J.; Gao, H.; Yang, J.; Mou, B.; et al. Ustilaginoidea virens Secretes a Family of Phosphatases That Stabilize the Negative Immune Regulator OsMPK6 and Suppress Plant Immunity. Plant Cell 2022, 34, 3088–3109. [Google Scholar] [CrossRef]
- Peng, Z.; Hu, Y.; Zhang, J.; Huguet-Tapia, J.C.; Block, A.K.; Park, S.; Sapkota, S.; Liu, Z.; Liu, S.; White, F.F. Xanthomonas translucens Commandeers the Host Rate-Limiting Step in ABA Biosynthesis for Disease Susceptibility. Proc. Natl. Acad. Sci. USA 2019, 116, 20938–20946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Wang, S.; Sun, R.; He, X.; Liu, Y.; Song, C. Xanthomonas oryzae Pv. oryzae Type III Effector PthXo3JXOV Suppresses Innate Immunity, Induces Susceptibility and Binds to Multiple Targets in Rice. FEMS Microbiol. Lett. 2018, 365, fny037. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Mukaihara, T. Ralstonia solanacearum Type III Effector RipAL Targets Chloroplasts and Induces Jasmonic Acid Production to Suppress Salicylic Acid-Mediated Defense Responses in Plants. Plant Cell Physiol. 2018, 59, 2576–2589. [Google Scholar] [CrossRef]
- Hu, Y.; Huang, H.; Cheng, X.; Shu, X.; White, A.P.; Stavrinides, J.; Köster, W.; Zhu, G.; Zhao, Z.; Wang, Y. A Global Survey of Bacterial Type III Secretion Systems and Their Effectors. Environ. Microbiol. 2017, 19, 3879–3895. [Google Scholar] [CrossRef] [PubMed]
- Ruano-Gallego, D.; Sanchez-Garrido, J.; Kozik, Z.; Núñez-Berrueco, E.; Cepeda-Molero, M.; Mullineaux-Sanders, C.; Naemi-Baghshomali Clark, J.; Slater, S.L.; Wagner, N.; Glegola-Madejska, I.; et al. Type III Secretion System Effectors Form Robust and Flexible Intracellular Virulence Networks. Science 2021, 371, eabc9531. [Google Scholar] [CrossRef]
- Wagner, S.; Grin, I.; Malmsheimer, S.; Singh, N.; Torres-Vargas, C.E.; Westerhausen, S. Bacterial Type III Secretion Systems: A Complex Device for the Delivery of Bacterial Effector Proteins into Eukaryotic Host Cells. FEMS Microbiol. Lett. 2018, 365, fny201. [Google Scholar] [CrossRef]
- Collmer, A.; Badel, J.L.; Charkowski, A.O.; Deng, W.-L.; Fouts, D.E.; Ramos, A.R.; Rehm, A.H.; Anderson, D.M.; Schneewind, O.; van Dijk, K.; et al. Pseudomonas syringae Hrp Type III Secretion System and Effector Proteins. Proc. Natl. Acad. Sci. USA 2000, 97, 8770–8777. [Google Scholar] [CrossRef] [Green Version]
- Xin, X.-F.; He, S.Y. Pseudomonas syringae pv. tomato DC3000: A Model Pathogen for Probing Disease Susceptibility and Hormone Signaling in Plants. Annu. Rev. Phytopathol. 2013, 51, 473–498. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.-L.; Collmer, A. Defining Essential Processes in Plant Pathogenesis with Pseudomonas syringae pv. tomato DC3000 Disarmed Polymutants and a Subset of Key Type III Effectors. Mol. Plant Pathol. 2018, 19, 1779–1794. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Tian, F.; Wamboldt, Y.; Alfano, J.R. The Majority of the Type III Effector Inventory of Pseudomonas syringae pv. tomato DC3000 Can Suppress Plant Immunity. Mol. Plant-Microbe Interact. 2009, 22, 1069–1080. [Google Scholar] [CrossRef] [Green Version]
- Nakano, M.; Ichinose, Y.; Mukaihara, T. Ralstonia solanacearum Type III Effector RipAC Targets SGT1 to Suppress Effector-Triggered Immunity. Plant Cell Physiol. 2020, 61, 2067–2076. [Google Scholar] [CrossRef] [PubMed]
- Abramovitch, R.B.; Kim, Y.-J.; Chen, S.; Dickman, M.B.; Martin, G.B. Pseudomonas Type III Effector AvrPtoB Induces Plant Disease Susceptibility by Inhibition of Host Programmed Cell Death. EMBO J. 2003, 22, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Abramovitch, R.B.; Martin, G.B. AvrPtoB: A Bacterial Type III Effector That Both Elicits and Suppresses Programmed Cell Death Associated with Plant Immunity. FEMS Microbiol. Lett. 2005, 245, 1–8. [Google Scholar] [CrossRef]
- Xian, L.; Yu, G.; Wei, Y.; Rufian, J.S.; Li, Y.; Zhuang, H.; Xue, H.; Morcillo, R.J.L.; Macho, A.P. A Bacterial Effector Protein Hijacks Plant Metabolism to Support Pathogen Nutrition. Cell Host Microbe 2020, 28, 548–557.e7. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Xu, N.; Bhandari, D.D.; Lapin, D.; Sun, X.; Luo, X.; Wang, Y.; Cao, J.; Wang, H.; Coaker, G.; et al. Bacterial Effector Targeting of a Plant Iron Sensor Facilitates Iron Acquisition and Pathogen Colonization. Plant Cell 2021, 33, 2015–2031. [Google Scholar] [CrossRef]
- Yu, Y.; Streubel, J.; Balzergue, S.; Champion, A.; Boch, J.; Koebnik, R.; Feng, J.; Verdier, V.; Szurek, B. Colonization of Rice Leaf Blades by an African Strain of Xanthomonas oryzae pv. oryzae Depends on a New TAL Effector That Induces the Rice Nodulin-3 Os11N3 Gene. Mol. Plant-Microbe Interact. 2011, 24, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Cernadas, R.A.; Doyle, E.L.; Niño-Liu, D.O.; Wilkins, K.E.; Bancroft, T.; Wang, L.; Schmidt, C.L.; Caldo, R.; Yang, B.; White, F.F.; et al. Code-Assisted Discovery of TAL Effector Targets in Bacterial Leaf Streak of Rice Reveals Contrast with Bacterial Blight and a Novel Susceptibility Gene. PLoS Pathog. 2014, 10, e1003972. [Google Scholar] [CrossRef] [PubMed]
- Dou, D.; Zhou, J.-M. Phytopathogen Effectors Subverting Host Immunity: Different Foes, Similar Battleground. Cell Host Microbe 2012, 12, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Kazan, K.; Lyons, R. Intervention of Phytohormone Pathways by Pathogen Effectors. Plant Cell 2014, 26, 2285–2309. [Google Scholar] [CrossRef] [Green Version]
- Cox, K.L.; Meng, F.; Wilkins, K.E.; Li, F.; Wang, P.; Booher, N.J.; Carpenter, S.C.D.; Chen, L.-Q.; Zheng, H.; Gao, X.; et al. TAL Effector Driven Induction of a SWEET Gene Confers Susceptibility to Bacterial Blight of Cotton. Nat. Commun. 2017, 8, 15588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, A.R.; Morbitzer, R.; Lahaye, T.; Staskawicz, B.J. TALE-Induced BHLH Transcription Factors That Activate a Pectate Lyase Contribute to Water Soaking in Bacterial Spot of Tomato. Proc. Natl. Acad. Sci. USA 2017, 114, E897–E903. [Google Scholar] [CrossRef] [Green Version]
- Zlobin, N.; Lebedeva, M.; Monakhova, Y.; Ustinova, V.; Taranov, V. An ERF121 Transcription Factor from Brassica oleracea Is a Target for the Conserved TAL-Effectors from Different Xanthomonas campestris pv. campestris Strains. Mol. Plant Pathol. 2021, 22, 618–624. [Google Scholar] [CrossRef]
- Xue, L.; Tang, B.; Chen, W.; Luo, J. DeepT3: Deep Convolutional Neural Networks Accurately Identify Gram-Negative Bacterial Type III Secreted Effectors Using the N-Terminal Sequence. Bioinformatics 2019, 35, 2051–2057. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Y.; Pu, X.; Li, M. Effective Prediction of Bacterial Type IV Secreted Effectors by Combined Features of Both C-Termini and N-Termini. J. Comput. Aided Mol. Des. 2017, 31, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Dhroso, A.; Eidson, S.; Korkin, D. Genome-Wide Prediction of Bacterial Effector Candidates across Six Secretion System Types Using a Feature-Based Statistical Framework. Sci. Rep. 2018, 8, 17209. [Google Scholar] [CrossRef]
- Plett, J.M.; Kemppainen, M.; Kale, S.D.; Kohler, A.; Legué, V.; Brun, A.; Tyler, B.M.; Pardo, A.G.; Martin, F. A Secreted Effector Protein of Laccaria bicolor Is Required for Symbiosis Development. Curr. Biol. 2011, 21, 1197–1203. [Google Scholar] [CrossRef]
- Field, K.J.; Pressel, S. Unity in Diversity: Structural and Functional Insights into the Ancient Partnerships between Plants and Fungi. New Phytol. 2018, 220, 996–1011. [Google Scholar] [CrossRef]
- Nishad, R.; Ahmed, T.; Rahman, V.J.; Kareem, A. Modulation of Plant Defense System in Response to Microbial Interactions. Front. Microbiol. 2020, 11, 1298. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 2022, 11, 386. [Google Scholar] [CrossRef]
- Jaiswal, S.K.; Mohammed, M.; Ibny, F.Y.I.; Dakora, F.D. Rhizobia as a Source of Plant Growth-Promoting Molecules: Potential Applications and Possible Operational Mechanisms. Front. Sustain. Food Syst. 2021, 4, 619676. [Google Scholar] [CrossRef]
- Kloppholz, S.; Kuhn, H.; Requena, N. A Secreted Fungal Effector of Glomus intraradices Promotes Symbiotic Biotrophy. Curr. Biol. CB 2011, 21, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Plett, J.M.; Daguerre, Y.; Wittulsky, S.; Vayssières, A.; Deveau, A.; Melton, S.J.; Kohler, A.; Morrell-Falvey, J.L.; Brun, A.; Veneault-Fourrey, C.; et al. Effector MiSSP7 of the Mutualistic Fungus Laccaria bicolor Stabilizes the Populus JAZ6 Protein and Represses Jasmonic Acid (JA) Responsive Genes. Proc. Natl. Acad. Sci. USA 2014, 111, 8299–8304. [Google Scholar] [CrossRef] [Green Version]
- Voß, S.; Betz, R.; Heidt, S.; Corradi, N.; Requena, N. RiCRN1, a Crinkler Effector from the Arbuscular Mycorrhizal Fungus Rhizophagus irregularis, Functions in Arbuscule Development. Front. Microbiol. 2018, 9, 2068. [Google Scholar] [CrossRef]
- Wang, P.; Jiang, H.; Boeren, S.; Dings, H.; Kulikova, O.; Bisseling, T.; Limpens, E. A Nuclear-Targeted Effector of Rhizophagus irregularis Interferes with Histone 2B Mono-Ubiquitination to Promote Arbuscular Mycorrhization. New Phytol. 2021, 230, 1142–1155. [Google Scholar] [CrossRef]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial Effects of Endophytic Fungi Colonization on Plants. Appl. Microbiol. Biotechnol. 2019, 103, 3327–3340. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lutzoni, F. Diversity and Host Range of Foliar Fungal Endophytes: Are Tropical Leaves Biodiversity Hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden Fungi, Emergent Properties: Endophytes and Microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [Green Version]
- Cord-Landwehr, S.; Melcher, R.L.J.; Kolkenbrock, S.; Moerschbacher, B.M. A Chitin Deacetylase from the Endophytic Fungus Pestalotiopsis sp. Efficiently Inactivates the Elicitor Activity of Chitin Oligomers in Rice Cells. Sci. Rep. 2016, 6, 38018. [Google Scholar] [CrossRef]
- Wawra, S.; Fesel, P.; Widmer, H.; Timm, M.; Seibel, J.; Leson, L.; Kesseler, L.; Nostadt, R.; Hilbert, M.; Langen, G.; et al. The Fungal-Specific β-Glucan-Binding Lectin FGB1 Alters Cell-Wall Composition and Suppresses Glucan-Triggered Immunity in Plants. Nat. Commun. 2016, 7, 13188. [Google Scholar] [CrossRef]
- Zeng, J.; Xu, T.; Cao, L.; Tong, C.; Zhang, X.; Luo, D.; Han, S.; Pang, P.; Fu, W.; Yan, J.; et al. The Role of Iron Competition in the Antagonistic Action of the Rice Endophyte Streptomyces Sporocinereus OsiSh-2 Against the Pathogen Magnaporthe Oryzae. Microb. Ecol. 2018, 76, 1021–1029. [Google Scholar] [CrossRef]
- Rajani, P.; Rajasekaran, C.; Vasanthakumari, M.M.; Olsson, S.B.; Ravikanth, G.; Uma Shaanker, R. Inhibition of Plant Pathogenic Fungi by Endophytic Trichoderma spp. through Mycoparasitism and Volatile Organic Compounds. Microbiol. Res. 2021, 242, 126595. [Google Scholar] [CrossRef]
- Abdel-Rahim, I.R.; Abo-Elyousr, K.A.M. Talaromyces pinophilus Strain AUN-1 as a Novel Mycoparasite of Botrytis cinerea, the Pathogen of Onion Scape and Umbel Blights. Microbiol. Res. 2018, 212–213, 1–9. [Google Scholar] [CrossRef]
- Mehetre, S.T.; Mukherjee, P.K. Trichoderma Improves Nutrient Use Efficiency in Crop Plants. In Nutrient Use Efficiency: From Basics to Advances; Rakshit, A., Singh, H.B., Sen, A., Eds.; Springer: New Delhi, India, 2015; pp. 173–180. ISBN 978-81-322-2169-2. [Google Scholar]
- Li, R.-X.; Cai, F.; Pang, G.; Shen, Q.-R.; Li, R.; Chen, W. Solubilisation of Phosphate and Micronutrients by Trichoderma harzianum and Its Relationship with the Promotion of Tomato Plant Growth. PLoS ONE 2015, 10, e0130081. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.Q.; Lan, F.; Qiao, Y.M.; Wei, J.G.; Huang, R.S.; Li, L.B. Endophytic Fungi Harbored in the Root of Sophora tonkinensis Gapnep: Diversity and Biocontrol Potential against Phytopathogens. MicrobiologyOpen 2017, 6, e00437. [Google Scholar] [CrossRef]
- Mohamad, O.A.A.; Li, L.; Ma, J.-B.; Hatab, S.; Xu, L.; Guo, J.-W.; Rasulov, B.A.; Liu, Y.-H.; Hedlund, B.P.; Li, W.-J. Evaluation of the Antimicrobial Activity of Endophytic Bacterial Populations from Chinese Traditional Medicinal Plant Licorice and Characterization of the Bioactive Secondary Metabolites Produced by Bacillus atrophaeus Against Verticillium dahliae. Front. Microbiol. 2018, 9, 924. [Google Scholar] [CrossRef] [Green Version]
- Saravanakumar, K.; Fan, L.; Fu, K.; Yu, C.; Wang, M.; Xia, H.; Sun, J.; Li, Y.; Chen, J. Cellulase from Trichoderma harzianum Interacts with Roots and Triggers Induced Systemic Resistance to Foliar Disease in Maize. Sci. Rep. 2016, 6, 35543. [Google Scholar] [CrossRef]
- Siddaiah, C.N.; Satyanarayana, N.R.; Mudili, V.; Kumar Gupta, V.; Gurunathan, S.; Rangappa, S.; Huntrike, S.S.; Srivastava, R.K. Elicitation of Resistance and Associated Defense Responses in Trichoderma hamatum Induced Protection against Pearl Millet Downy Mildew Pathogen. Sci. Rep. 2017, 7, 43991. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Ji, S.; Wang, Z.; Zhang, H.; Wang, Y.; Liu, Z. Trichoderma asperellum Xylanases Promote Growth and Induce Resistance in Poplar. Microbiol. Res. 2021, 248, 126767. [Google Scholar] [CrossRef]
- Veloso, J.; Díaz, J. The Non-Pathogenic Fusarium Oxysporum Fo47 Induces Distinct Responses in Two Closely Related Solanaceae Plants against the Pathogen Verticillium dahliae. J. Fungi 2021, 7, 344. [Google Scholar] [CrossRef]
- La Spada, F.; Stracquadanio, C.; Riolo, M.; Pane, A.; Cacciola, S.O. Trichoderma Counteracts the Challenge of Phytophthora nicotianae Infections on Tomato by Modulating Plant Defense Mechanisms and the Expression of Crinkler, Necrosis-Inducing Phytophthora Protein 1, and Cellulose-Binding Elicitor Lectin Pathogenic Effectors. Front. Plant Sci. 2020, 11, 583539. [Google Scholar] [CrossRef]
- Durán, P.; Thiergart, T.; Garrido-Oter, R.; Agler, M.; Kemen, E.; Schulze-Lefert, P.; Hacquard, S. Microbial Interkingdom Interactions in Roots Promote Arabidopsis Survival. Cell 2018, 175, 973–983.e14. [Google Scholar] [CrossRef]
- Levy, A.; Salas Gonzalez, I.; Mittelviefhaus, M.; Clingenpeel, S.; Herrera Paredes, S.; Miao, J.; Wang, K.; Devescovi, G.; Stillman, K.; Monteiro, F.; et al. Genomic Features of Bacterial Adaptation to Plants. Nat. Genet. 2017, 50, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Eitzen, K.; Sengupta, P.; Kroll, S.; Kemen, E.; Doehlemann, G. A Fungal Member of the Arabidopsis thaliana Phyllosphere Antagonizes Albugo laibachii via a GH25 Lysozyme. eLife 2021, 10, e65306. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shen, D.; Ge, C.; Du, Y.; Lin, L.; Liu, J.; Bai, T.; Jing, M.; Qian, G.; Dou, D. Filamentous Phytophthora Pathogens Deploy Effectors to Interfere with Bacterial Growth and Motility. Front. Microbiol. 2020, 11, 581511. [Google Scholar] [CrossRef]
- Snelders, N.C.; Kettles, G.J.; Rudd, J.J.; Thomma, B.P.H.J. Plant Pathogen Effector Proteins as Manipulators of Host Microbiomes? Mol. Plant Pathol. 2018, 19, 257–259. [Google Scholar] [CrossRef] [PubMed]
- Kettles, G.J.; Bayon, C.; Sparks, C.A.; Canning, G.; Kanyuka, K.; Rudd, J.J. Characterization of an Antimicrobial and Phytotoxic Ribonuclease Secreted by the Fungal Wheat Pathogen Zymoseptoria tritici. New Phytol. 2018, 217, 320–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán-Guzmán, P.; Alemán-Duarte, M.I.; Delaye, L.; Herrera-Estrella, A.; Olmedo-Monfil, V. Identification of Effector-like Proteins in Trichoderma spp. and Role of a Hydrophobin in the Plant-Fungus Interaction and Mycoparasitism. BMC Genet. 2017, 18, 16. [Google Scholar] [CrossRef] [Green Version]
- Laur, J.; Ramakrishnan, G.B.; Labbé, C.; Lefebvre, F.; Spanu, P.D.; Bélanger, R.R. Effectors Involved in Fungal–Fungal Interaction Lead to a Rare Phenomenon of Hyperbiotrophy in the Tritrophic System Biocontrol Agent–Powdery Mildew–Plant. New Phytol. 2018, 217, 713–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vannier, N.; Agler, M.; Hacquard, S. Microbiota-Mediated Disease Resistance in Plants. PLoS Pathog. 2019, 15, e1007740. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, C.; Liang, W.; Liu, S. Rhizosphere Microbiome: The Emerging Barrier in Plant-Pathogen Interactions. Front. Microbiol. 2021, 12, 772420. [Google Scholar] [CrossRef]
- Redkar, A.; Sabale, M.; Zuccaro, A.; Di Pietro, A. Determinants of Endophytic and Pathogenic Lifestyle in Root Colonizing Fungi. Curr. Opin. Plant Biol. 2022, 67, 102226. [Google Scholar] [CrossRef] [PubMed]
- Shayanthan, A.; Ordoñez, P.A.C.; Oresnik, I.J. The Role of Synthetic Microbial Communities (SynCom) in Sustainable Agriculture. Front. Agron. 2022, 4, 896307. [Google Scholar] [CrossRef]
- Liu, H.; Qiu, Z.; Ye, J.; Verma, J.P.; Li, J.; Singh, B.K. Effective Colonisation by a Bacterial Synthetic Community Promotes Plant Growth and Alters Soil Microbial Community. J. Sustain. Agric. Environ. 2022, 1, 30–42. [Google Scholar] [CrossRef]
- Marín, O.; González, B.; Poupin, M.J. From Microbial Dynamics to Functionality in the Rhizosphere: A Systematic Review of the Opportunities with Synthetic Microbial Communities. Front. Plant Sci. 2021, 12, 650609. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, L.K.; Miyauchi, S.; Uhlmann, C.; Garrido-Oter, R.; Langen, G.; Wawra, S.; Niu, Y.; Guan, R.; Robertson-Albertyn, S.; Bulgarelli, D.; et al. The Fungal Root Endophyte Serendipita vermifera Displays Inter-Kingdom Synergistic Beneficial Effects with the Microbiota in Arabidopsis thaliana and Barley. ISME J. 2022, 16, 876–889. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, R.L.; Vismans, G.; Yu, K.; Song, Y.; de Jonge, R.; Burgman, W.P.; Burmølle, M.; Herschend, J.; Bakker, P.A.H.M.; Pieterse, C.M.J. Disease-Induced Assemblage of a Plant-Beneficial Bacterial Consortium. ISME J. 2018, 12, 1496–1507. [Google Scholar] [CrossRef] [Green Version]
- Hacquard, S.; Spaepen, S.; Garrido-Oter, R.; Schulze-Lefert, P. Interplay Between Innate Immunity and the Plant Microbiota. Annu. Rev. Phytopathol. 2017, 55, 565–589. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Chen, X.; Kemppainen, M.; Pardo, A.G.; Veneault-Fourrey, C.; Kohler, A.; Martin, F.M. The Small Secreted Effector Protein MiSSP7.6 of Laccaria bicolor Is Required for the Establishment of Ectomycorrhizal Symbiosis. Environ. Microbiol. 2020, 22, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Pellegrin, C.; Daguerre, Y.; Ruytinx, J.; Guinet, F.; Kemppainen, M.; dit Frey, N.F.; Puech-Pagès, V.; Hecker, A.; Pardo, A.G.; Martin, F.M.; et al. Laccaria bicolor MiSSP8 Is a Small-Secreted Protein Decisive for the Establishment of the Ectomycorrhizal Symbiosis. Environ. Microbiol. 2019, 21, 3765–3779. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Rodriguez-Moreno, L.; Mansurkhodzaev, A.; Wang, P.; van den Berg, W.; Gasciolli, V.; Cottaz, S.; Fort, S.; Thomma, B.P.H.J.; Bono, J.-J.; et al. A Lysin Motif Effector Subverts Chitin-Triggered Immunity to Facilitate Arbuscular Mycorrhizal Symbiosis. New Phytol. 2020, 225, 448–460. [Google Scholar] [CrossRef] [Green Version]
- Plett, J.M.; Plett, K.L.; Wong-Bajracharya, J.; de Freitas Pereira, M.; Costa, M.D.; Kohler, A.; Martin, F.; Anderson, I.C. Mycorrhizal Effector PaMiSSP10b Alters Polyamine Biosynthesis in Eucalyptus Root Cells and Promotes Root Colonization. New Phytol. 2020, 228, 712–727. [Google Scholar] [CrossRef] [PubMed]
- Ratu, S.T.N.; Teulet, A.; Miwa, H.; Masuda, S.; Nguyen, H.P.; Yasuda, M.; Sato, S.; Kaneko, T.; Hayashi, M.; Giraud, E.; et al. Rhizobia Use a Pathogenic-like Effector to Hijack Leguminous Nodulation Signalling. Sci. Rep. 2021, 11, 2034. [Google Scholar] [CrossRef] [PubMed]
- Xin, D.-W.; Liao, S.; Xie, Z.-P.; Hann, D.R.; Steinle, L.; Boller, T.; Staehelin, C. Functional Analysis of NopM, a Novel E3 Ubiquitin Ligase (NEL) Domain Effector of Rhizobium sp. Strain NGR234. PLoS Pathog. 2012, 8, e1002707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenzel, M.; Friedrich, L.; Göttfert, M.; Zehner, S. The Type III-Secreted Protein NopE1 Affects Symbiosis and Exhibits a Calcium-Dependent Autocleavage Activity. Mol. Plant-Microbe Interact. MPMI 2010, 23, 124–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nostadt, R.; Hilbert, M.; Nizam, S.; Rovenich, H.; Wawra, S.; Martin, J.; Küpper, H.; Mijovilovich, A.; Ursinus, A.; Langen, G.; et al. A Secreted Fungal Histidine- and Alanine-Rich Protein Regulates Metal Ion Homeostasis and Oxidative Stress. New Phytol. 2020, 227, 1174–1188. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhong, D.; Xie, W.; He, Y.; Zheng, Y.; Lin, Y.; Chen, Z.; Han, Y.; Tian, D.; Liu, W.; et al. Functional Identification of Novel Cell Death-Inducing Effector Proteins from Magnaporthe oryzae. Rice 2019, 12, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas-Marina, M.A.; Isordia-Jasso, M.I.; Islas-Osuna, M.A.; Delgado-Sánchez, P.; Jiménez-Bremont, J.F.; Rodríguez-Kessler, M.; Rosales-Saavedra, M.T.; Herrera-Estrella, A.; Casas-Flores, S. The Epl1 and Sm1 Proteins from Trichoderma atroviride and Trichoderma virens Differentially Modulate Systemic Disease Resistance against Different Life Style Pathogens in Solanum lycopersicum. Front. Plant Sci. 2015, 6, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djonović, S.; Pozo, M.J.; Dangott, L.J.; Howell, C.R.; Kenerley, C.M. Sm1, a Proteinaceous Elicitor Secreted by the Biocontrol Fungus Trichoderma virens Induces Plant Defense Responses and Systemic Resistance. Mol. Plant-Microbe Interact. 2006, 19, 838–853. [Google Scholar] [CrossRef] [Green Version]
- Rai, M.; Agarkar, G. Plant–Fungal Interactions: What Triggers the Fungi to Switch among Lifestyles? Crit. Rev. Microbiol. 2016, 42, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xie, J.; Fu, Y.; Cheng, J.; Qu, Z.; Zhao, Z.; Cheng, S.; Chen, T.; Li, B.; Wang, Q.; et al. A 2-Kb Mycovirus Converts a Pathogenic Fungus into a Beneficial Endophyte for Brassica Protection and Yield Enhancement. Mol. Plant 2020, 13, 1420–1433. [Google Scholar] [CrossRef]
- Czislowski, E.; Zeil-Rolfe, I.; Aitken, E.A.B. Effector Profiles of Endophytic Fusarium Associated with Asymptomatic Banana (Musa sp.) Hosts. Int. J. Mol. Sci. 2021, 22, 2508. [Google Scholar] [CrossRef] [PubMed]
- Constantin, M.E.; Fokkens, L.; de Sain, M.; Takken, F.L.W.; Rep, M. Number of Candidate Effector Genes in Accessory Genomes Differentiates Pathogenic from Endophytic Fusarium Oxysporum Strains. Front. Plant Sci. 2021, 12, 761740. [Google Scholar] [CrossRef]
- Sonah, H.; Deshmukh, R.K.; Bélanger, R.R. Computational Prediction of Effector Proteins in Fungi: Opportunities and Challenges. Front. Plant Sci. 2016, 7, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.A.; Bertazzoni, S.; Turo, C.J.; Syme, R.A.; Hane, J.K. Bioinformatic Prediction of Plant–Pathogenicity Effector Proteins of Fungi. Curr. Opin. Microbiol. 2018, 46, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Nur, M.; Wood, K.; Michelmore, R. EffectorO: Motif-Independent Prediction of Effectors in Oomycete Genomes Using Machine Learning and Lineage Specificity. bioRxiv 2021. In Print. [Google Scholar] [CrossRef]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Manners, J.M.; Singh, K.B.; Taylor, J.M. Advances and Challenges in Computational Prediction of Effectors from Plant Pathogenic Fungi. PLoS Pathog. 2015, 11, e1004806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperschneider, J.; Dodds, P.N. EffectorP 3.0: Prediction of Apoplastic and Cytoplasmic Effectors in Fungi and Oomycetes. Mol. Plant-Microbe Interact. 2021, 35, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Gardiner, D.M.; Dodds, P.N.; Tini, F.; Covarelli, L.; Singh, K.B.; Manners, J.M.; Taylor, J.M. EffectorP: Predicting Fungal Effector Proteins from Secretomes Using Machine Learning. New Phytol. 2016, 210, 743–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Singh, K.B.; Taylor, J.M. Improved Prediction of Fungal Effector Proteins from Secretomes with EffectorP 2.0. Mol. Plant Pathol. 2018, 19, 2094–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperschneider, J.; Dodds, P.N.; Singh, K.B.; Taylor, J.M. ApoplastP: Prediction of Effectors and Plant Proteins in the Apoplast Using Machine Learning. New Phytol. 2018, 217, 1764–1778. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wang, P.; Han, S.; Wang, L.; Zhao, Y.; Juan, L. FunEffector-Pred: Identification of Fungi Effector by Activate Learning and Genetic Algorithm Sampling of Imbalanced Data. IEEE Access 2020, 8, 57674–57683. [Google Scholar] [CrossRef]
- Wagner, N.; Avram, O.; Gold-Binshtok, D.; Zerah, B.; Teper, D.; Pupko, T. Effectidor: An Automated Machine-Learning-Based Web Server for the Prediction of Type-III Secretion System Effectors. Bioinformatics 2022, 38, 2341–2343. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.A.B.; Rozano, L.; Debler, J.W.; Mancera, R.L.; Moolhuijzen, P.M.; Hane, J.K. An Automated and Combinative Method for the Predictive Ranking of Candidate Effector Proteins of Fungal Plant Pathogens. Sci. Rep. 2021, 11, 19731. [Google Scholar] [CrossRef]
- Ramalingam, J.; Raveendra, C.; Savitha, P.; Vidya, V.; Chaithra, T.L.; Velprabakaran, S.; Saraswathi, R.; Ramanathan, A.; Arumugam Pillai, M.P.; Arumugachamy, S.; et al. Gene Pyramiding for Achieving Enhanced Resistance to Bacterial Blight, Blast, and Sheath Blight Diseases in Rice. Front. Plant Sci. 2020, 11, 591457. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, M.G.C.; Martin, F.; Variar, M.; Bhatt, J.C.; L Perez-Quintero, A.; Leung, H.; Leach, J.E.; Vera Cruz, C.M. Accumulating Candidate Genes for Broad-Spectrum Resistance to Rice Blast in a Drought-Tolerant Rice Cultivar. Sci. Rep. 2021, 11, 21502. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.S.-A.; Mukhtar, M.S.; Mansoor, S. Genome Editing: Targeting Susceptibility Genes for Plant Disease Resistance. Trends Biotechnol. 2018, 36, 898–906. [Google Scholar] [CrossRef]
- Kieu, N.P.; Lenman, M.; Wang, E.S.; Petersen, B.L.; Andreasson, E. Mutations Introduced in Susceptibility Genes through CRISPR/Cas9 Genome Editing Confer Increased Late Blight Resistance in Potatoes. Sci. Rep. 2021, 11, 4487. [Google Scholar] [CrossRef] [PubMed]


| Effector Classification | Organism Type | Organism | Effector Name and Uniprot ID | Function | References |
|---|---|---|---|---|---|
| Resistance or Defense Associated Effectors | Biotrophic fungus | Cladosporium fulvum | Avr4 | Induces ETI when recognized by host resistance protein Cf-4; protects fungal cell walls against hydrolysis by plant chitinases | [91,92] |
| Biotrophic fungus | Cladosporium fulvum | Avr4E | Induces ETI; recognized by resistance protein Hcr9-4E | [93] | |
| Biotrophic fungus | Cladosporium fulvum | Ecp6 | Induces ETI when recognized by resistance protein Cf-ECP6; binds to fungal chitin to prevent chitin-triggered immunity in host | [94,95] | |
| Biotrophic fungus | Melamspora lini | AvrM | Induces ETI in host; recognized by resistance protein M | [96] | |
| Hemibiotrophic fungus | Magnaporthe oryzae | AvrPia (B9WZW9) | Induces ETI in host; recognized by resistance protein RGA5 | [97] | |
| Hemibiotrophic fungus | Magnaporthe oryzae | AVR-Pik (C4B8B8) | Induces ETI in host; recognized by resistance protein Pik | [98] | |
| Hemibiotrophic fungus | Magnaporthe oryzae | PWT3 | Recognized by host resistance protein Rwt3 | [99] | |
| Hemibiotrophic oomycete | Phytophthora infestans | AVRamr3 | Recognized by host resistance protein Rpi-amr3 | [100] | |
| Hemibiotrophic fungus | Ascochyta lentis | AlAvr1 | Unidentified resistance gene; ETI induced in host | [101] | |
| Biotrophic fungus | Puccinia polysora | AvrRppC | Recognized by host resistance protein RppC | [102] | |
| Bacteria | Ralstonia solanacearum | RipB | Recognized by host resistance protein Roq1 | [103] | |
| Bacteria | Ralstonia solanacearum | RipJ | Unidentified resistance gene; ETI induced in host | [104] | |
| Bacteria | Ralstonia solanacearum | RipAZ1 | Unidentified resistance gene; ETI induced in host | [105] | |
| Bacteria | Pseudomonas syringae pv. syringae strain 61 | HopA1Pss61 | Recognized by RPS6 resistance protein; ETI induced | [106] | |
| Susceptibility Associated Effectors | Biotrophic fungus | Ustilago maydis | Umrip1 | Targets susceptibility factor ZmLox3, ZmLox3 represses ROS burst | [81] |
| Hemibiotrophic oomycete | Phytophthora infestans | Pi02860 | Targets susceptibility factor NRL1. NLR1 promotes degradation of positive regulator of immunity, StSWAP70 | [107,108] | |
| Hemibiotrophic oomycete | Phytophthora infestans | Pi04314/RD24 | Targets PP1 catalytic subunits causing their re-localization from the nucleolus to the nucleoplasm; Pi04314-PP1c holoenzymes negatively regulate salicylic acid and jasmonic acid pathways | [109] | |
| Hemibiotrophic oomycete | Phytophthora sojae | PsAvh52 | Targets susceptibility factor GmTAP1, causing relocation from the cytoplasm to the nucleus. GmTAP1 promotes H3K9 acetylation to promote disease susceptibility | [110] | |
| Hemibiotrophic oomycete | Phytophthora infestans | PiAvr2 | Interacts with BRI1-SUPPRESSOR1-like (BSL) BSL1, BSL2, and BSL3; BSL1 and BSL3 suppress INF1-triggered cell death (PTI) | [111,112] | |
| Necrotrophic fungus | Pyrenophora tritici-repentis | ToxA (Host-selective toxin) | Targets Tsn1, susceptibility factor involved in ToxA-triggered cell death which favors necrotrophy | [113,114] | |
| Necrotrophic fungus | Parastagonospora nodorum | SnTox1 (Host-selective toxin) | Targets Snn1, susceptibility factor involved in SnTox1-triggered cell death which favors necrotrophy; protects fungus from host chitinases | [90,115] | |
| Necrotophic fungus | Pyrenophora tritici-repentis | PtrToxB (Host-selective toxin) | Targets Tsc2, susceptibility factor involved in PtrToxB triggered cell death which favors necrotrophy | [116,117] | |
| Hemibiotrophic fungus | Phytophthora sp. | PSR2 | Inhibits secondary siRNA (PPR-siRNAs) production in Arabidopsis to promote disease susceptibility | [118] | |
| Biotrophic oomycete | Hyaloperonospora arabidopsidis | HaRxL21 | Responsible for transcriptional repression via interaction with TPL/TPR1 Arabidopsis proteins | [119] | |
| Necrotrophic fungus | Sclerotinia sclerotiorum | SsITL | Inhibits SA accumulation through interaction with CAS receptor in chloroplast | [120,121] | |
| Biotrophic fungus | Puccinia striiformis f. sp. tritici | Pst_12806 | Reduces photosynthesis and ROS accumulation; interacts with TaISP, a subunit of Cyt b6/f in the chloroplast | [122] | |
| Biotrophic fungus | Puccinia striiformis f. sp. tritici | PstGSRE1 | Disrupts nuclear localization of a ROS associated transcription factor TaLOL2 to suppress ROS-mediated cell death | [123] | |
| Biotrophic fungus | Puccinia striiformis f. sp. tritici | PstGSRE4 | Inhibits the enzyme activity of wheat copper zinc superoxide dismutase TaCZSOD2 reducing H2O2 accumulation and HR | [124] | |
| Biotrophic fungus | Puccinia striiformis f. sp. tritici | Pst18363 | Pst18363 stabilizes TaNUDX23, which suppresses ROS accumulation inducing susceptibility | [48] | |
| Biotrophic fungus | Ustilaginoidea virens | SCRE6 | Interacts with and dephosphorylates the target OsMPK6 for its stabilization, suppressing plant immunity | [125] | |
| Bacteria | Xanthomonas translucens pv. undulosa | Tal8 | Upregulates expression of the host gene 9-cis-epoxycarotenoid dioxygenase (TaNCED-5BS) involved in the biosynthesis of abscisic acid; decreases ex-pression of defense gene TaNPR1 | [126] | |
| Bacteria | Xanthomonas oryzae pv. oryzae | PthXo3JXOV | Upregulates expression of the susceptibility gene OsSWEET14 to trigger sugar release; effector also inhibits HR and callose deposition | [127] | |
| Bacteria | Ralstonia solanacearum | RipAL | Putative lipase that catalyzes the release of linoleic acid from chloroplast lipids; induces JA production and suppresses SA signaling | [128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Todd, J.N.A.; Carreón-Anguiano, K.G.; Islas-Flores, I.; Canto-Canché, B. Microbial Effectors: Key Determinants in Plant Health and Disease. Microorganisms 2022, 10, 1980. https://doi.org/10.3390/microorganisms10101980
Todd JNA, Carreón-Anguiano KG, Islas-Flores I, Canto-Canché B. Microbial Effectors: Key Determinants in Plant Health and Disease. Microorganisms. 2022; 10(10):1980. https://doi.org/10.3390/microorganisms10101980
Chicago/Turabian StyleTodd, Jewel Nicole Anna, Karla Gisel Carreón-Anguiano, Ignacio Islas-Flores, and Blondy Canto-Canché. 2022. "Microbial Effectors: Key Determinants in Plant Health and Disease" Microorganisms 10, no. 10: 1980. https://doi.org/10.3390/microorganisms10101980