

The Role of Mycangial Fungi Associated with Ambrosia Beetles (Euwallacea interjectus) in Fig Wilt Disease: Dual Inoculation of Fusarium kuroshium and Ceratocystis ficicola Can Bring Fig Saplings to Early Symptom Development
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
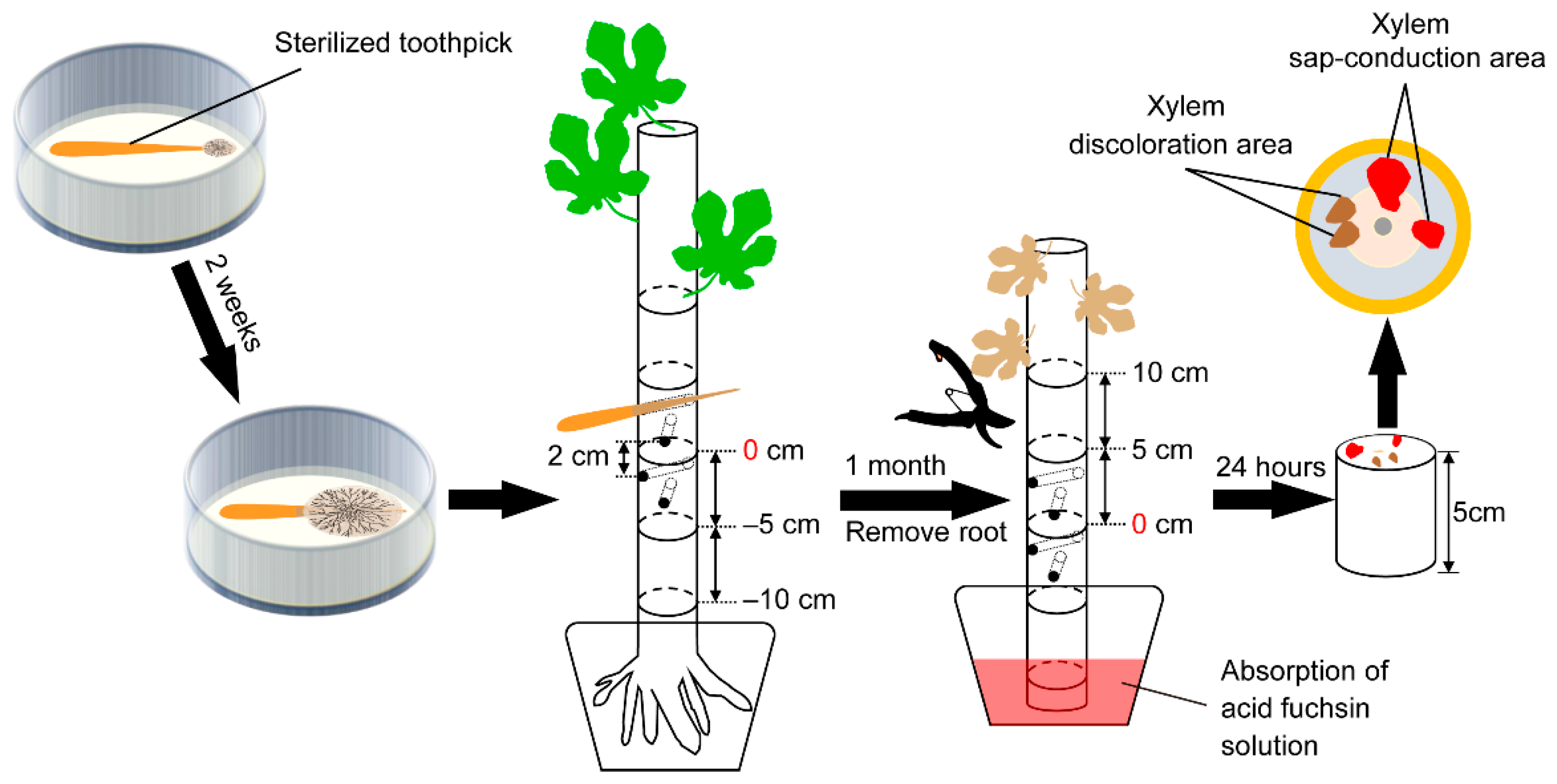
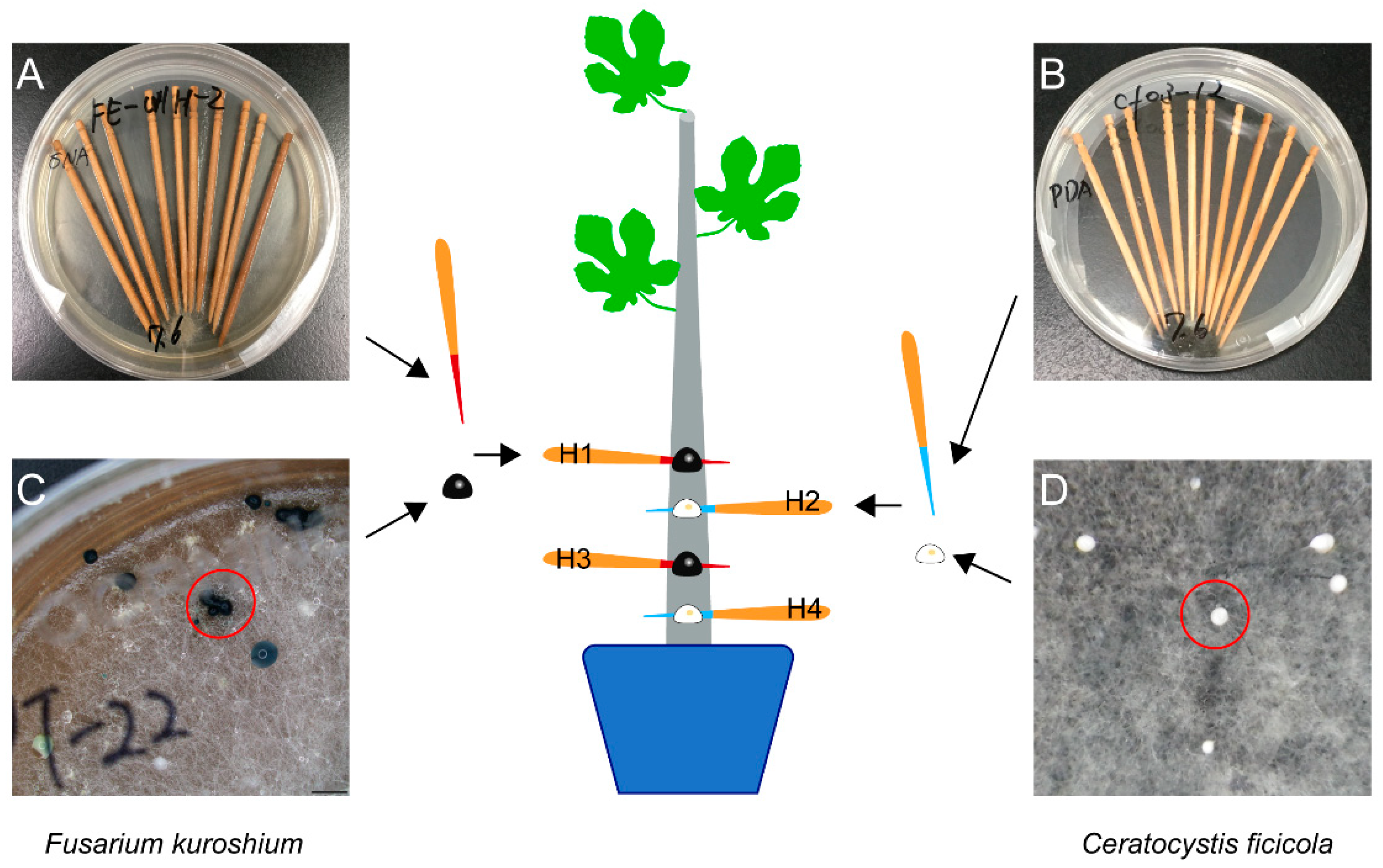
2.1. Inoculation Test
2.2. Classification of Wilt Symptoms
2.3. Xylem Sap-Conduction Test
2.4. Re-Isolation of Inoculated Fungi
2.5. Statistical Analysis
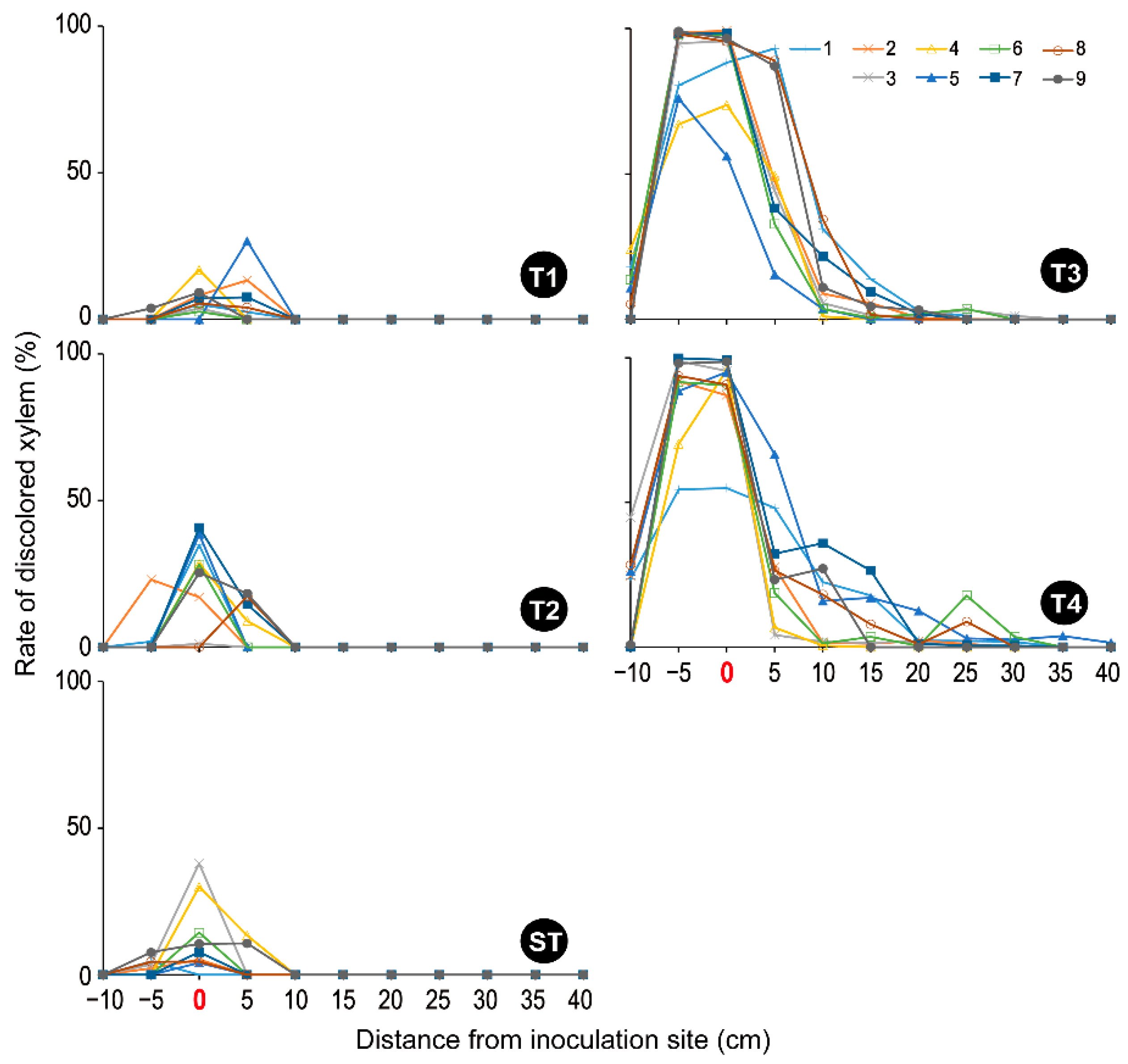
3. Results
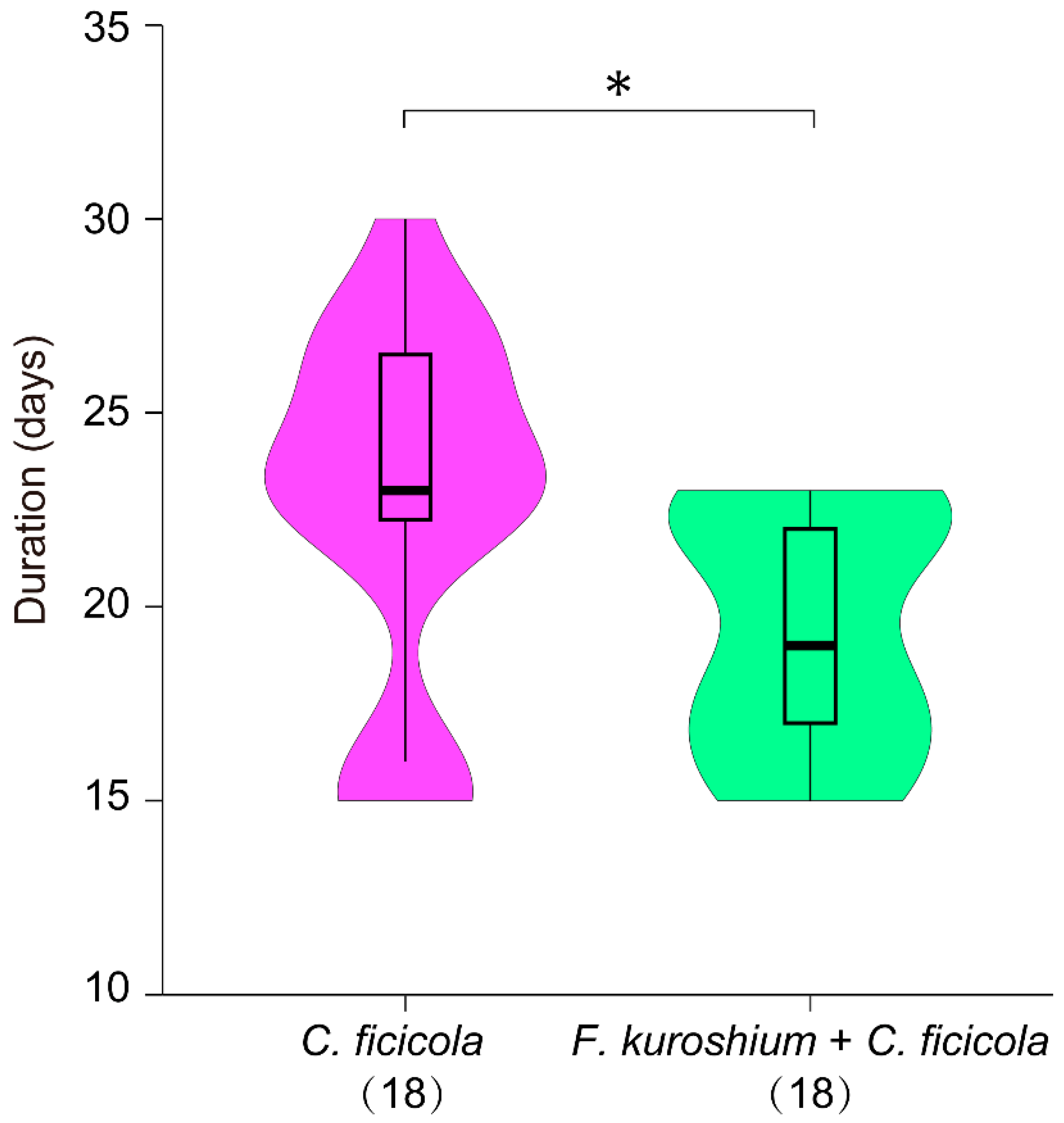
3.1. Symptom Development
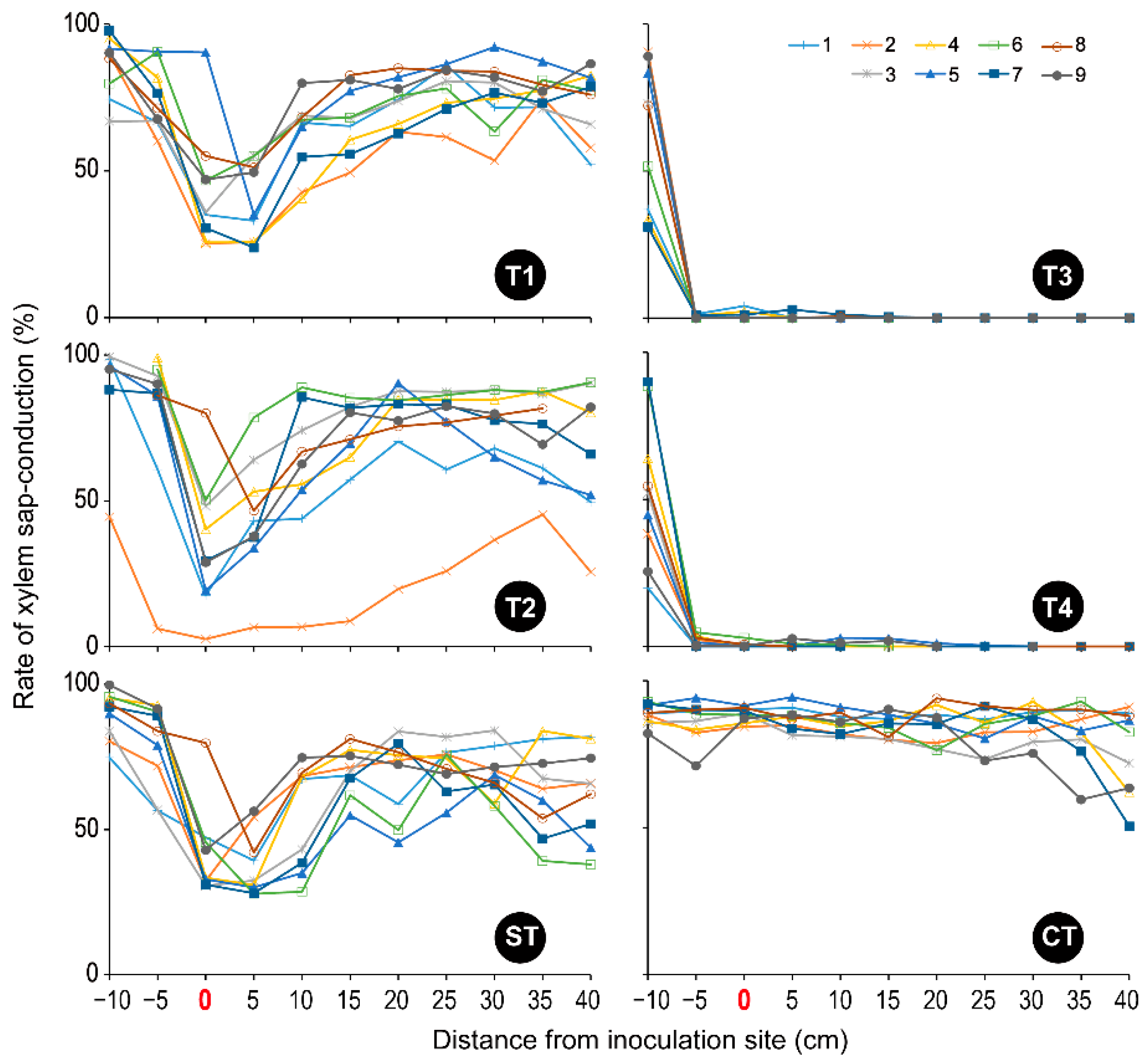
3.2. Xylem Sap-Conduction
3.3. Re-Isolation of Inoculated Fungi
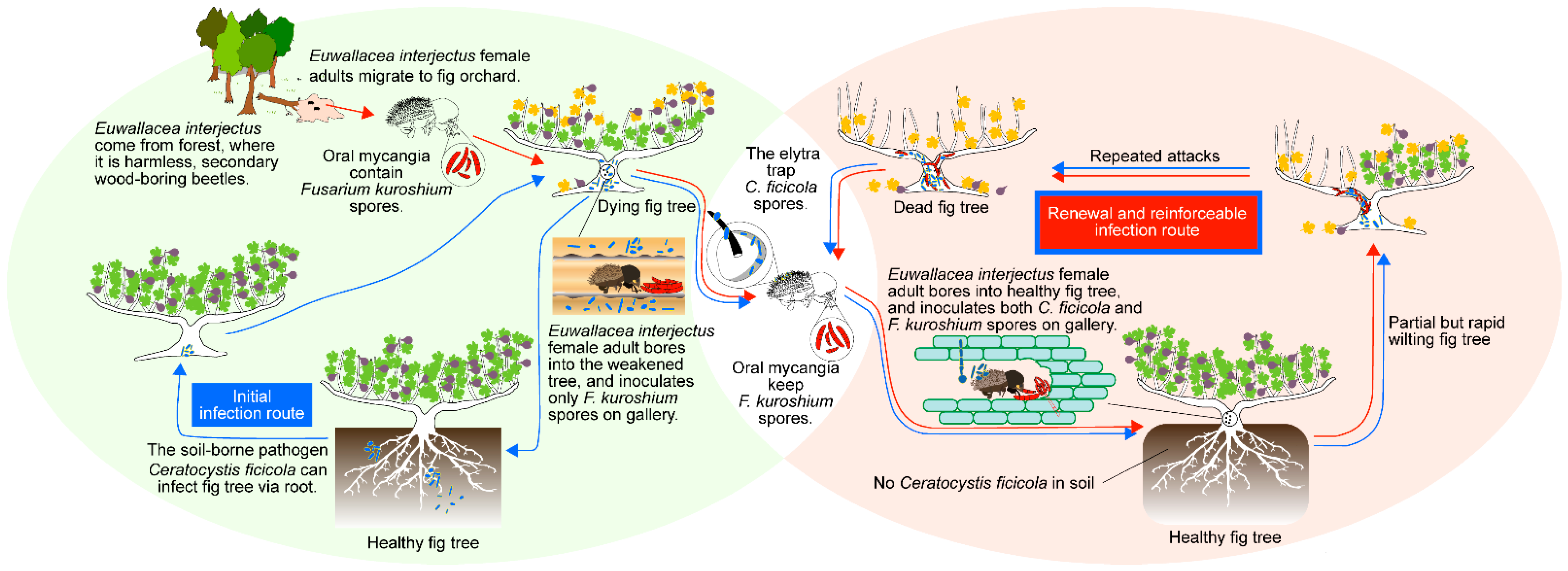
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kato, K.; Yokota, K.; Miyagawa, T. A new disease, Ceratocystis canker of fig caused by Ceratocystis fimbriata Ellis et Halsted. Plant Prot. 1982, 36, 55–59. [Google Scholar]
- Kajitani, Y.; Masuya, H. Ceratocystis ficicola sp. nov., a causal fungus of fig canker in Japan. Mycoscience 2011, 52, 349–353. [Google Scholar] [CrossRef]
- Yakushiji, H.; Morita, T.; Jikumaru, S. Ceratocystis canker resistance in BC1 populations of interspecific hybridization of fig (Ficus carica) and F. erecta. Sci. Hortic. 2019, 252, 71–76. [Google Scholar] [CrossRef]
- Kajii, C.; Morita, T.; Jikumaru, S.; Kajimura, H.; Yamaoka, Y.; Kuroda, K. Xylem dysfunction in Ficus carica infected with wilt fungus Ceratocystis ficicola and the role of the vector beetle Euwallacea interjectus. IAWA J. 2013, 34, 301–312. [Google Scholar] [CrossRef]
- Kajitani, Y. Infection strategies of pathogens observed in fig wilt disease and grapevine swelling arm. Plant-Pathog. Fungi Conf. 2017, 17, 9–20. [Google Scholar]
- Morita, T.; Hara, H.; Mise, D.; Jikumaru, S. A case study of Ceratocystis canker epidemic in relation with Euwallacea interjectus infestation. Ann. Rept. Kansai Plant Prot. 2012, 54, 29–34. [Google Scholar] [CrossRef]
- Morita, T.; Jikumaru, S.; Kuroda, K. Disease development in Ficus carica plants after inoculation with Ceratocystis ficicola. (1) Relation between xylem dysfunction and wilt symptoms. Jpn. J. Phytopathol. 2016, 82, 301–309. [Google Scholar] [CrossRef]
- Morita, T.; Jikumaru, S.; Yakushiji, H. Evaluation of Ceratocystis canker resistance between BC1, interspecific hybrids of Ficus carica and F. erecta, and existing resistant fig varieties, F. carica, in relation to infection via soil. Ann. Rept. Kansai Plant Prot. 2020, 62, 149–152. [Google Scholar] [CrossRef]
- Hosomi, A.; Kawaradani, M. Degree of resistance to Ceratocystis canker by Ceratocystis fimbriata in fig varieties ‘Zidi’ and ‘King’. Ann. Rept. Kansai Plant Prot. 2004, 46, 29–32. [Google Scholar] [CrossRef]
- Morita, T.; Mise, D.; Jikumaru, S. Relative resistance of Ficus erecta and cultivars of F. carica to Ceratocystis fimbriata using their seedlings. Ann. Rept. Kansai Plant Prot. 2011, 53, 51–52. [Google Scholar] [CrossRef]
- Tsopelas, P.; Soulioti, N.; Wingfiield, M.J.; Barnes, I.; Marincowitz, S.; Tjamos, E.C.; Paplomatas, E.J. Ceratocystis ficicola causing a serious disease of Ficus carica in Greece. Phytopathol. Mediterr. 2021, 60, 337–349. [Google Scholar] [CrossRef]
- Landi, L.; Braccini, C.L.; Knížek, M.; Pereyra, V.A.; Marvaldi, A.E. A newly detected exotic ambrosia beetle in Argentina: Euwallacea interjectus (Coleoptera: Curculionidae: Scolytinae). Fla. Entomol. 2019, 102, 240–242. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Ernstsons, A.S.; Sun, R.; Hulcr, J.; Gao, L. The infestation and habitat of the ambrosia beetle Euwallacea interjectus (Coleoptera: Curculionidae: Scolytinae) in the riparian zone of Shanghai, China. Agric. For. Entomol. 2020, 23, 104–109. [Google Scholar] [CrossRef]
- Aoki, T.; Smith, J.A.; Kasson, M.T.; Freeman, S.; Geiser, D.M.; Geering, A.D.; O’Donnell, K. Three novel Ambrosia Fusarium Clade species producing clavate macroconidia known (F. floridanum and F. obliquiseptatum) or predicted (F. tuaranense) to be farmed by Euwallacea spp. (Coleoptera: Scolytinae) on woody hosts. Mycologia 2019, 111, 919–935. [Google Scholar] [CrossRef] [PubMed]
- Kajitani, Y. The possibility of transmission of fig Ceratocystis canker disease by an ambrosia beetle (Xyleborus interjectus Eichhoff). Ann. Phytopathol. Soc. Jpn. 1996, 62, 275. [Google Scholar]
- Kajitani, Y. The dispersal period of the Xyleborus interjectus (Coleoptera, Scolytidae), a vector of the fig Ceratocystis canker, and the organ carrying the causal fungus. Ann. Phytopathol. Soc. Jpn. 1999, 65, 377. [Google Scholar]
- Jiang, Z.-R.; Kinoshita, S.; Sasaki, O.; Cognato, A.I.; Kajimura, H. Non-destructive observation of the mycangia of Euwallacea interjectus (Blandford) (Coleoptera: Curculionidae: Scolytinae) using X-ray computed tomography. Entomol. Sci. 2019, 22, 173–181. [Google Scholar] [CrossRef]
- Jiang, Z.-R.; Masuya, H.; Kajimura, H. Novel symbiotic association between Euwallacea ambrosia beetle and Fusarium fungus on fig trees in Japan. Front. Microbiol. 2021, 12, 725210. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.-R.; Masuya, H.; Kajimura, H. Fungal flora in adult females of the rearing population of ambrosia beetle Euwallacea interjectus (Blandford) (Coleoptera: Curculionidae: Scolytinae): Does it differ from the wild population? Diversity 2022, 14, 535. [Google Scholar] [CrossRef]
- Na, F.; Carrillo, J.D.; Mayorquin, J.S.; Ndinga-Muniania, C.; Stajich, J.E.; Stouthamer, R.; Huang, Y.T.; Lin, Y.T.; Chen, C.Y.; Eskalen, A. Two novel fungal symbionts Fusarium kuroshium sp. nov. and Graphium kuroshium sp. nov. of Kuroshio shot hole borer (Euwallacea sp. nr. fornicatus) cause Fusarium dieback on woody host species in California. Plant Dis. 2018, 102, 1154–1164. [Google Scholar] [CrossRef]
- Kasson, M.T.; O’Donnell, K.; Rooney, A.P.; Sink, S.; Ploetz, R.C.; Ploetz, J.N.; Konkol, J.L.; Carrillo, D.; Freeman, S.; Mendel, Z.; et al. An inordinate fondness for Fusarium: Phylogenetic diversity of fusaria cultivated by ambrosia beetles in the genus Euwallacea on avocado and other plant hosts. Fungal Genet. Biol. 2013, 56, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Al-Hatmi, A.M.; Ahmed, S.A.; Van Diepeningen, A.D.; Drogari-Apiranthitou, M.; Verweij, P.E.; Meis, J.F.; De Hoog, G.S. Fusarium metavorans sp. nov.: The frequent opportunist ‘FSSC6’. Med. Mycol. 2018, 56, S144–S152. [Google Scholar] [CrossRef] [PubMed]
- Borman, A.M.; Johnson, E.M. Name changes for fungi of medical importance, 2018 to 2019. J. Clin. Microbiol. 2021, 59, e01811-20. [Google Scholar] [CrossRef] [PubMed]
- Mendel, Z.; Protasov, A.; Sharon, M.; Zveibil, A.; Ben Yehuda, S.; O’Donnell, K.; Rabaglia, R.; Wysoki, M.; Freeman, S. An Asian ambrosia beetle Euwallacea fornicatus and its novel symbiotic fungus Fusarium sp. pose a serious threat to the Israeli avocado industry. Phytoparasitica 2012, 40, 235–238. [Google Scholar] [CrossRef]
- Egonyu, J.P.; Torto, B. Responses of the ambrosia beetle Xylosandrus compactus (Coleoptera: Curculionidea: Scolytinae) to volatile constituents of its symbiotic fungus Fusarium solani (Hypocreales: Nectriaceae). Arthropod-Plant Int. 2018, 12, 9–20. [Google Scholar] [CrossRef]
- Kuroda, K.; Chuma, I.; Kihara, T.; Murakami, T.; Takashina, K.; Hiraoka, D.; Kameyama, N. First report of Fusarium solani species complex as a causal agent of Erythrina variegata decline and death after gall formation by Quadrastichus erythrinae on Okinawa Island, Japan. J. Gen. Plant Pathol. 2017, 83, 344–357. [Google Scholar] [CrossRef]
- Takashina, K.; Chuma, I.; Kajimura, H.; Kameyama, N.; Goto, C.; Kuroda, K. Pathogenicity and distribution of Fusarium solani isolates associated with Erythrina decline in Japan. Plant Dis. 2020, 104, 731–742. [Google Scholar] [CrossRef]
- Back, M.A.; Haydock, P.P.J.; Jenkinson, P. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathol. 2002, 51, 683–697. [Google Scholar] [CrossRef]
- Hajihassani, A.; Smiley, R.W.; Afshar, F.J. Effects of co-inoculation with Pratylenchus thornei and Fusarium culmorum on growth and yield of winter wheat. Plant Dis. 2013, 97, 1470–1477. [Google Scholar] [CrossRef]
- Upadhaya, A.; Yan, G.; Secor, G.; Robinson, A.P. Effects of co-inoculation with Pratylenchus penetrans and Fusarium oxysporum on growth and yield of potato cultivar Red Norland. Am. J. Potato Res. 2020, 97, 246–255. [Google Scholar] [CrossRef]
- Freeman, S.; Sharon, M.; Maymon, M.; Mendel, Z.; Protasov, A.; Aoki, T.; Eskalen, A.; O’Donnell, K. Fusarium euwallaceae sp. nov.—A symbiotic fungus of Euwallacea sp., an invasive ambrosia beetle in Israel and California. Mycologia 2013, 105, 1595–1606. [Google Scholar] [CrossRef] [PubMed]
- Sumida, S.; Morita, T.; Kuroda, K. Invasion strategy of Ceratocystis ficicola from soil to host (Ficus carica). Tree For. Health 2016, 20, 30–31. [Google Scholar]
- Morita, T.; Mise, D.; Jikumaru, S. Observation of external and internal disease symptoms in the fig cultivar ‘Houraishi’ planter in soil contaminated with Ceratocystis ficicola. Bull. Hiroshima Pref. Technol. Res. Inst. Agric. Technol. Res. Ctr. 2018, 92, 1–9. [Google Scholar]
- Morita, T.; Jikumaru, S. A case study of Ceratocystis canker epidemic following the invasion of Euwallacea interjectus. Plant Prot. 2013, 67, 275–278. [Google Scholar]
- Jiang, Z.-R.; Kajimura, H. Earwig preying on ambrosia beetle: Evaluating predatory process and prey preference. J. Appl. Entomol. 2020, 144, 743–750. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Test Code | No. of Saplings | Inoculum | |
|---|---|---|---|---|
| Source | Morphology | |||
| Wound inoculation | T1 | 18 | Fusarium kuroshiuma | Hyphae + conidial masses |
| T2 | 18 | Neocosmospora metavoransb | Hyphae | |
| T3 | 18 | Ceratocystis ficicolac | Hyphae + ascospore masses | |
| T4 | 18 | F. kuroshium + C. ficicolad | Hyphae + conidial/ascospore masses | |
| Control | ST | 18 | Sterilized toothpicks e | — |
| CT | 18 | None f | — | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Z.-R.; Morita, T.; Jikumaru, S.; Kuroda, K.; Masuya, H.; Kajimura, H. The Role of Mycangial Fungi Associated with Ambrosia Beetles (Euwallacea interjectus) in Fig Wilt Disease: Dual Inoculation of Fusarium kuroshium and Ceratocystis ficicola Can Bring Fig Saplings to Early Symptom Development. Microorganisms 2022, 10, 1912. https://doi.org/10.3390/microorganisms10101912
Jiang Z-R, Morita T, Jikumaru S, Kuroda K, Masuya H, Kajimura H. The Role of Mycangial Fungi Associated with Ambrosia Beetles (Euwallacea interjectus) in Fig Wilt Disease: Dual Inoculation of Fusarium kuroshium and Ceratocystis ficicola Can Bring Fig Saplings to Early Symptom Development. Microorganisms. 2022; 10(10):1912. https://doi.org/10.3390/microorganisms10101912
Chicago/Turabian StyleJiang, Zi-Ru, Takeshige Morita, Shota Jikumaru, Keiko Kuroda, Hayato Masuya, and Hisashi Kajimura. 2022. "The Role of Mycangial Fungi Associated with Ambrosia Beetles (Euwallacea interjectus) in Fig Wilt Disease: Dual Inoculation of Fusarium kuroshium and Ceratocystis ficicola Can Bring Fig Saplings to Early Symptom Development" Microorganisms 10, no. 10: 1912. https://doi.org/10.3390/microorganisms10101912