
Seasonal Effect on Bacterial Communities Associated with the Rhizospheres of Polhillia, Wiborgia and Wiborgiella Species in the Cape Fynbos, South Africa

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Soil Sampling
2.2. Total Bacterial Genomic DNA Extraction from Rhizosphere Soils
2.3. Library Preparation
2.4. Sequencing and Data Assembling
3. Results
3.1. Site Elevation and Physical Properties of Rhizosphere Soils
3.2. Chemical Properties of Rhizosphere Soils
3.3. Quantity and Quality of Raw Sequences Generated
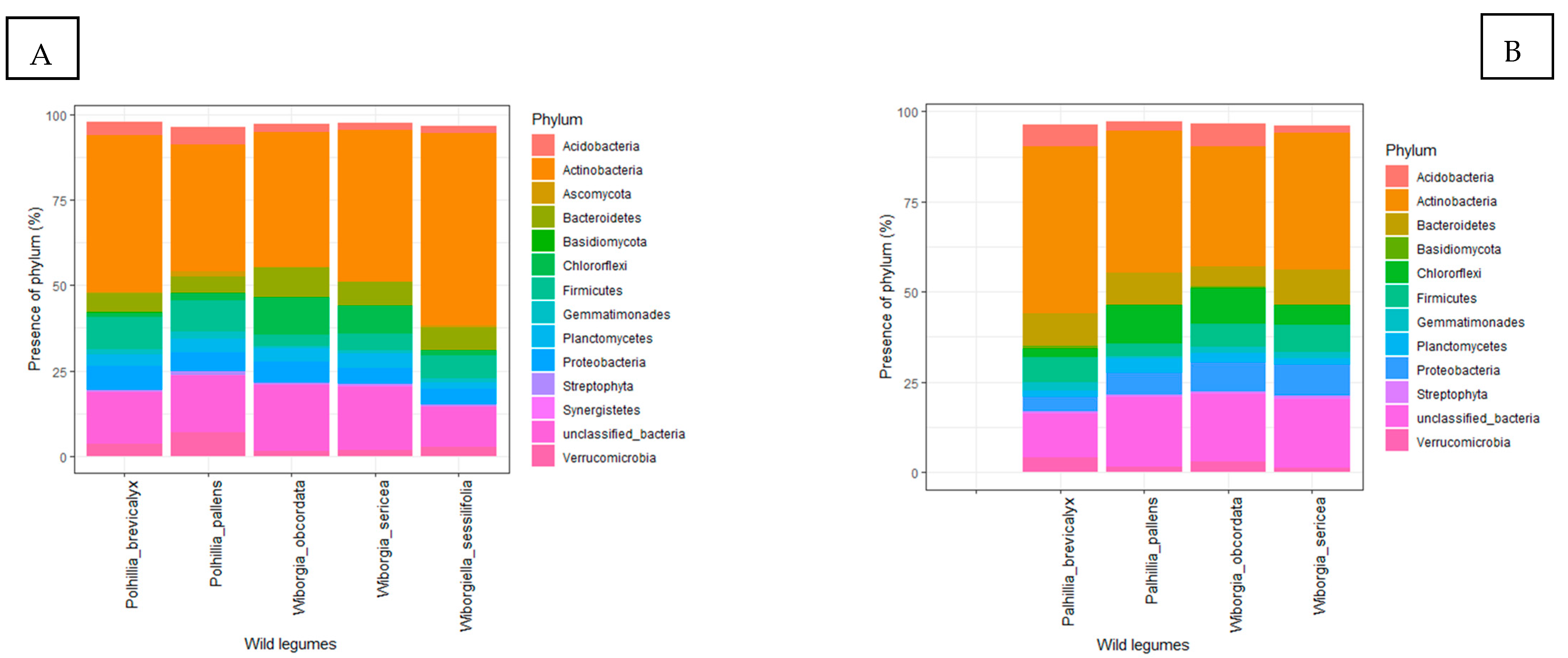
3.4. Rhizosphere Bacterial Communities
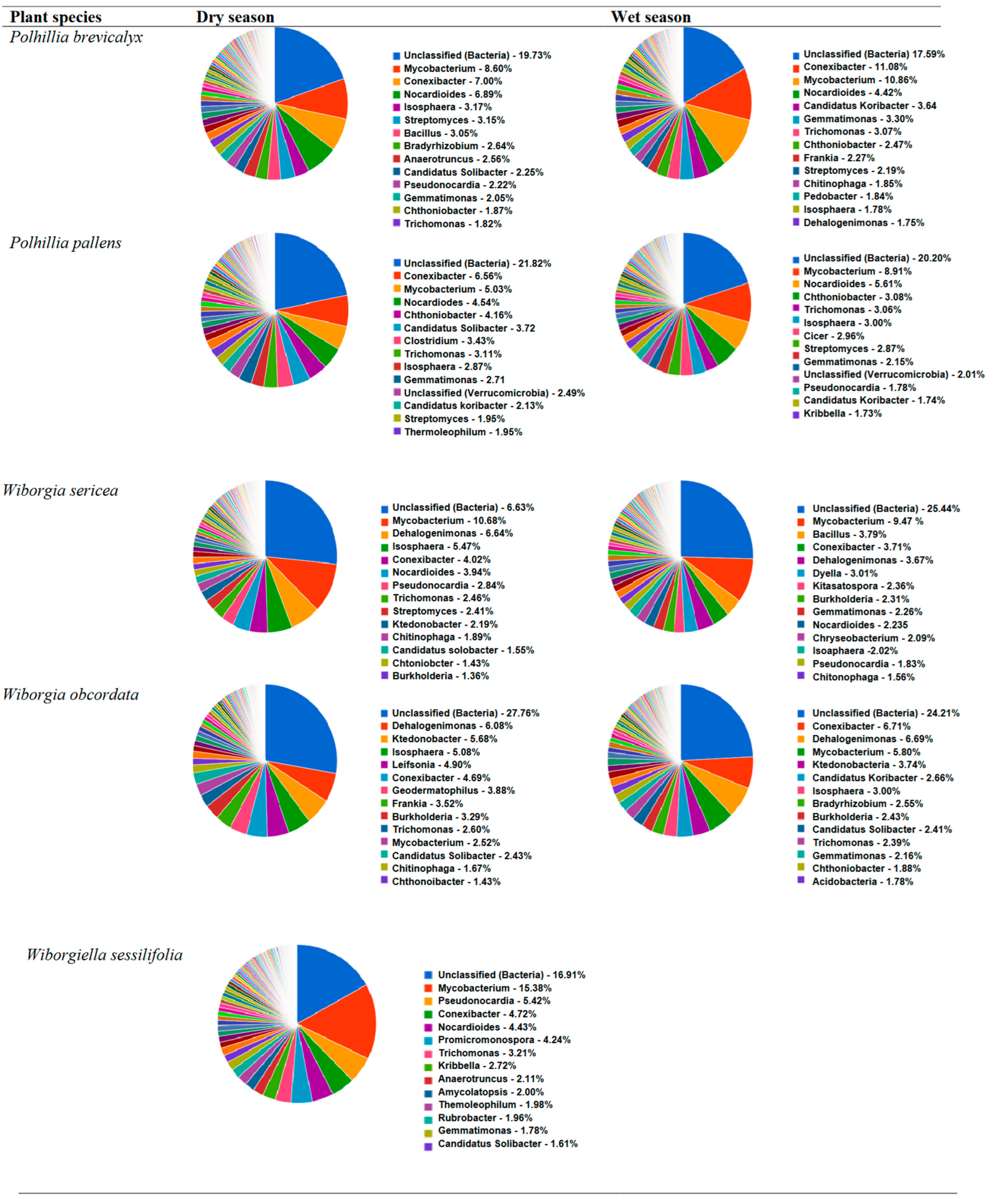
3.4.1. Polhillia Brevicalyx
3.4.2. Polhillia Pallens
3.4.3. Wiborgia Sericea
3.4.4. Wiborgia Obcordata
3.4.5. Wiborgiella Sessilifolia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Forest, F.; Colville, J.F.; Cowling, R.M. Evolutionary diversity patterns in the Cape flora of South Africa. In Phylogenetic Diversity; Springer: Berlin/Heidelberg, Germany, 2018; pp. 167–187. [Google Scholar]
- Cowling, R.M.; Pressey, R.L.; Rouget, M.; Lombard, A.T. A conservation plan for a global biodiversity hotspot—The Cape Floristic Region, South Africa. Biol. Conserv. 2003, 112, 191–216. [Google Scholar] [CrossRef] [Green Version]
- Forest, F.; Nänni, I.; Chase, M.W.; Crane, P.R.; Hawkins, J.A. Diversification of a large genus in a continental biodiversity hotspot: Temporal and spatial origin of Muraltia (Polygalaceae) in the Cape of South Africa. Mol. Phylogenet. Evol. 2007, 43, 60–74. [Google Scholar] [CrossRef]
- Boatwright, J.S.; Cupido, C.N. Aspalathus crewiana sp. nov.(Crotalarieae, Fabaceae) from the Western Cape Province, South Africa. Nord. J. Bot. 2011, 29, 513–517. [Google Scholar] [CrossRef]
- Lemaire, B.; Dlodlo, O.; Chimphango, S.; Stirton, C.; Schrire, B.; Boatwright, S.; Honnay, O.; Smets, E.; Sprent, J.; James, E.; et al. Symbiotic diversity, specificity and distribution of rhizobia in native legumes of the Core Cape Subregion (South Africa). FEMS Microbiol. Ecol. 2015, 91, 2–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postma, A.; Slabbert, E.; Postma, F.; Jacobs, K. Soil bacterial communities associated with natural and commercial Cyclopia spp. FEMS Microbiol. Ecol. 2016, 92, fiw016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brink, C.; Postma, A.; Jacobs, K. Rhizobial diversity and function in rooibos (Aspalathus linearis) and honeybush (Cyclopia spp.) plants: A review. S. Afr. J. Bot. 2017, 110, 80–86. [Google Scholar] [CrossRef]
- Mpai, T.; Jaiswal, S.K.; Dakora, F.D. Accumulation of phosphorus and carbon and the dependency on biological N-2 fixation for nitrogen nutrition in Polhillia, Wiborgia and Wiborgiella species growing in natural stands in cape fynbos, South Africa. Symbiosis 2020, 81, 65–78. [Google Scholar] [CrossRef]
- Boatwright, J.S.; Tilney, P.M.; Van Wyk, B.-E. Taxonomy of Wiborgiella (Crotalarieae, Fabaceae), a genus endemic to the greater Cape Region of South Africa. Syst. Bot. 2010, 35, 325–340. [Google Scholar] [CrossRef]
- Ahmad, M.H.; Uddin, M.R.; McLaughlin, W. Characterization of indigenous rhizobia from wild legumes. FEMS Microbiol. Lett. 1984, 24, 197–203. [Google Scholar] [CrossRef]
- Howieson, J.G.; De Meyer, S.E.; Vivas-Marfisi, A.; Ratnayake, S.; Ardley, J.K.; Yates, R.J. Novel Burkholderia bacteria isolated from Lebeckia ambigua–a perennial suffrutescent legume of the fynbos. Soil Biol. Biochem. 2013, 60, 55–64. [Google Scholar] [CrossRef]
- Lemaire, B.; Van Cauwenberghe, J.; Verstraete, B.; Chimphango, S.; Stirton, C.; Honnay, O.; Smets, E.; Sprent, J.; James, E.K.; Muasya, A.M. Characterization of the papilionoid–Burkholderia interaction in the Fynbos biome: The diversity and distribution of beta-rhizobia nodulating Podalyria calyptrata (Fabaceae, Podalyrieae). Syst. Appl. Microbiol. 2016, 39, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Mpai, T.; Jaiswal, S.K.; Cupido, C.N.; Dakora, F.D. Ecological adaptation and phylogenetic analysis of microsymbionts nodulating Polhillia, Wiborgia and Wiborgiella species in the Cape fynbos, South Africa. Sci. Rep. 2021, 11, 1–17. [Google Scholar] [CrossRef]
- Cowling, R.M.; Procheş, Ş.; Partridge, T.C. Explaining the uniqueness of the Cape flora: Incorporating geomorphic evolution as a factor for explaining its diversification. Mol. Phylogenet. Evol. 2009, 51, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Muofhe, M.L.; Dakora, F.D. Nitrogen nutrition in nodulated field plants of the shrub tea legume Aspalathus linearis assessed using 15 N natural abundance. Plant Soil 1999, 209, 181. [Google Scholar] [CrossRef]
- Cramer, M.D. Phosphate as a Limiting Resource: Introduction. Plant Soil 2020, 334, 1–10. [Google Scholar] [CrossRef]
- Maseko, S.T.; Dakora, F.D. Rhizosphere acid and alkaline phosphatase activity as a marker of P nutrition in nodulated Cyclopia and Aspalathus species in the Cape fynbos of South Africa. S. Afr. J. Bot. 2013, 89, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Kamutando, C.N.; Vikram, S.; Kamgan-Nkuekam, G.; Makhalanyane, T.P.; Greve, M.; Le Roux, J.J.; Richardson, D.M.; Cowan, D.; Valverde, A. Soil nutritional status and biogeography influence rhizosphere microbial communities associated with the invasive tree Acacia dealbata. Sci. Rep. 2017, 7, 6472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moroenyane, I.; Chimphango, S.B.M.; Wang, J.; Kim, H.K.; Adams, J.M. Deterministic assembly processes govern bacterial community structure in the Fynbos, South Africa. Microb. Ecol. 2016, 72, 313–323. [Google Scholar] [CrossRef]
- Wahdan, S.F.M.; Heintz-Buschart, A.; Sansupa, C.; Tanunchai, B.; Wu, Y.-T.; Schädler, M.; Noll, M.; Purahong, W.; Buscot, F. Targeting the active rhizosphere microbiome of trifolium pratense in grassland evidences a stronger-than-expected belowground biodiversity-ecosystem functioning link. Front. Microbiol. 2021, 12, 73. [Google Scholar] [CrossRef]
- Alkorta, I.; Epelde, L.; Garbisu, C. Environmental parameters altered by climate change affect the activity of soil microorganisms involved in bioremediation. FEMS Microbiol. Lett. 2017, 364, fnx200. [Google Scholar] [CrossRef]
- Jaiswal, S.K.; Mohammed, M.; Dakora, F.D. Microbial community structure in the rhizosphere of the orphan legume Kersting’s groundnut [Macrotyloma geocarpum (Harms) Marechal & Baudet]. Mol. Biol. Rep. 2019, 46, 4471–4481. [Google Scholar] [PubMed]
- Zhou, Y.; Zhu, H.; Fu, S.; Yao, Q. Variation in soil microbial community structure associated with different legume species is greater than that associated with different grass species. Front. Microbiol. 2017, 8, 1007. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.K.; Naamala, J.; Dakora, F.D. Nature and mechanisms of aluminium toxicity, tolerance and amelioration in symbiotic legumes and rhizobia. Biol. Fertil. Soils 2018, 54, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Brink, C.J.; Postma, A.; Slabbert, E.; Postma, F.; Muasya, A.M.; Jacobs, K. Bacterial communities associated with natural and commercially grown rooibos (Aspalathus linearis). Pedosphere 2020, 30, 778–790. [Google Scholar] [CrossRef]
- Slabbert, E.; Kongor, R.Y.; Esler, K.J.; Jacobs, K. Microbial diversity and community structure in Fynbos soil. Mol. Ecol. 2010, 19, 1031–1041. [Google Scholar] [CrossRef]
- Moroenyane, I.; Chimphango, S.B.M.; Dong, K.; Tripathi, B.; Singh, D.; Adams, J.M. Neutral models predict biogeographical patterns of soil microbes at a local scale in Mediterranean heathlands, South Africa. Trans. R. Soc. S. Afr. 2019, 74, 139–150. [Google Scholar] [CrossRef]
- Jacobs, K.; Conradie, T.; Jacobs, S. Microbial communities in the fynbos region of South Africa: What happens during woody alien plant invasions. Diversity 2020, 12, 254. [Google Scholar] [CrossRef]
- Zhang, F.; Zhou, G. Estimation of vegetation water content using hyperspectral vegetation indices: A comparison of crop water indicators in response to water stress treatments for summer maize. BMC Ecol. 2019, 19, 18. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Mabiala, S.T.; Joseph, G.; Lebonguy, A.A.; Banga, A.B. Diversity of the Bacterial Community of Three Soils Revealed by Illumina-Miseq Sequencing of 16S rRNA Gene in the South of Brazzaville, Congo. Am. J. Microbiol. Res. 2020, 8, 141–149. [Google Scholar] [CrossRef]
- Nessner Kavamura, V.; Taketani, R.G.; Lançoni, M.D.; Andreote, F.D.; Mendes, R.; de Melo, I. Water regime influences bulk soil and rhizosphere of Cereus jamacaru bacterial communities in the Brazilian Caatinga biome. PLoS ONE 2013, 8, e73606. [Google Scholar] [CrossRef]
- Aguirre-Garrido, J.F.; Montiel-Lugo, D.; Hernández-Rodríguez, C.; Torres-Cortes, G.; Millán, V.; Toro, N.; Martínez-Abarca, F.; Ramírez-Saad, H.C. Bacterial community structure in the rhizosphere of three cactus species from semi-arid highlands in central Mexico. Antonie Van Leeuwenhoek 2012, 101, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N.; Verma, P.; Kumar, S.; Kumar, V.; Kumar, M.; Sugitha, T.C.K.; Singh, B.P.; Saxena, A.K.; Dhaliwal, H.S. Actinobacteria from rhizosphere: Molecular diversity, distributions, and potential biotechnological applications. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 13–41. [Google Scholar]
- Trujillo, M.E.; Riesco, R.; Benito, P.; Carro, L. Endophytic actinobacteria and the interaction of Micromonospora and nitrogen fixing plants. Front. Microbiol. 2015, 6, 1341. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Martinez, V.; Dowd, S.; Sun, Y.; Allen, V. Tag-encoded pyrosequencing analysis of bacterial diversity in a single soil type as affected by management and land use. Soil Biol. Biochem. 2008, 40, 2762–2770. [Google Scholar] [CrossRef]
- Le, X.H.; Franco, C.M.M.; Ballard, R.A.; Drew, E.A. Isolation and characterisation of endophytic actinobacteria and their effect on the early growth and nodulation of lucerne (Medicago sativa L.). Plant Soil 2016, 405, 13–24. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Andreote, F.D.; Reinhold-Hurek, B.; Sessitsch, A.; van Overbeek, L.S.; van Elsas, J.D. Rice root-associated bacteria: Insights into community structures across 10 cultivars. FEMS Microbiol. Ecol. 2011, 77, 154–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, K.-J.; Hahn, D.; Hengstmann, U.L.F.; Liesack, W.; Janssen, P.H. Characterization and identification of numerically abundant culturable bacteria from the anoxic bulk soil of rice paddy microcosms. Appl. Environ. Microbiol. 1999, 65, 5042–5049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Thakur, D. Antimicrobial biosynthetic potential and diversity of culturable soil actinobacteria from forest ecosystems of Northeast India. Sci. Rep. 2020, 10, 4104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, R.M.; da Silveira, É.L.; Scaquitto, D.C.; Pedrinho, E.A.N.; Val-Moraes, S.P.; Wickert, E.; Carareto-Alves, L.M.; Lemos, E.G.d.M. Molecular characterization of bacterial populations of different soils. Braz. J. Microbiol. 2006, 37, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Conradie, T.; Jacobs, K. Seasonal and agricultural response of Acidobacteria present in two fynbos rhizosphere soils. Diversity 2020, 12, 277. [Google Scholar] [CrossRef]
- Vieira, S.; Luckner, M.; Wanner, G.; Overmann, J. Luteitalea pratensis gen. nov., sp. nov. a new member of subdivision 6 Acidobacteria isolated from temperate grassland soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 1408–1414. [Google Scholar] [CrossRef]
- Tarafdar, J.C.; Rathore, I.; Shiva, V. Effect of Bt-transgenic cotton on soil biological health. Appl. Biol. Res. 2012, 14, 15–23. [Google Scholar]
- Oliveira, C.A.; Alves, V.M.C.; Marriel, I.E.; Gomes, E.A.; Scotti, M.R.; Carneiro, N.P.; Guimaraes, C.T.; Schaffert, R.E.; Sá, N.M.H. Phosphate solubilizing microorganisms isolated from rhizosphere of maize cultivated in an oxisol of the Brazilian Cerrado Biome. Soil Biol. Biochem. 2009, 41, 1782–1787. [Google Scholar] [CrossRef]
- Ambrosini, A.; Beneduzi, A.; Stefanski, T.; Pinheiro, F.G.; Vargas, L.K.; Passaglia, L.M.P. Screening of plant growth promoting rhizobacteria isolated from sunflower (Helianthus annuus L.). Plant Soil 2012, 356, 245–264. [Google Scholar] [CrossRef]
- Verma, P.; Yadav, A.N.; Kazy, S.K.; Saxena, A.K.; Suman, A. Elucidating the diversity and plant growth promoting attributes of wheat (Triticum aestivum) associated acidotolerant bacteria from southern hills zone of India. Natl. J. Life Sci. 2013, 10, 219–226. [Google Scholar]
- Verma, P.; Yadav, A.N.; Kazy, S.K.; Saxena, A.K.; Suman, A. Evaluating the diversity and phylogeny of plant growth promoting bacteria associated with wheat (Triticum aestivum) growing in central zone of India. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 432–447. [Google Scholar]
- Babalola, O.O.; Alawiye, T.T.; Rodriguez Lopez, C.M.; Ayangbenro, A.S. Shotgun metagenomic sequencing data of sunflower rhizosphere microbial community in South Africa. Data Brief 2020, 31, 105831. [Google Scholar] [CrossRef]
- Maphosa, F.; de Vos, W.M.; Smidt, H. Exploiting the ecogenomics toolbox for environmental diagnostics of organohalide-respiring bacteria. Trends Biotechnol. 2010, 28, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Chen, J.; Zhou, H.; Farzana, S.; Tam, N.F.Y. Vertical distribution of dehalogenating bacteria in mangrove sediment and their potential to remove polybrominated diphenyl ether contamination. Mar. Pollut. Bull. 2017, 124, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.E.; Kent, A.D.; Brisson, V.L.; Gaudin, A.C.M. Agricultural management and plant selection interactively affect rhizosphere microbial community structure and nitrogen cycling. Microbiome 2019, 7, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puozaa, D.K.; Jaiswal, S.K.; Dakora, F.D. Phylogeny and distribution of Bradyrhizobium symbionts nodulating cowpea (Vigna unguiculata L. Walp) and their association with the physicochemical properties of acidic African soils. Syst. Appl. Microbiol. 2019, 42, 403–414. [Google Scholar] [CrossRef]
- Miyambo, T.; Makhalanyane, T.P.; Cowan, D.A.; Valverde, A. Plants of the fynbos biome harbour host species-specific bacterial communities. FEMS Microbiol. Lett. 2016, 363, fnw122. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef]
- Peoples, M.B.; Brockwell, J.; Hunt, J.R.; Swan, A.D.; Watson, L.; Hayes, R.C.; Li, G.D.; Hackney, B.; Nuttall, J.G.; Davies, S.L.; et al. Factors affecting the potential contributions of N2 fixation by legumes in Australian pasture systems. Crop. Pasture Sci. 2013, 63, 759–786. [Google Scholar] [CrossRef]
- Pimratch, S.; Jogloy, S.; Vorasoot, N.; Toomsan, B.; Patanothai, A.; Holbrook, C.C. Relationship between Biomass Production and Nitrogen Fixation under Drought-Stress Conditions in Peanut Genotypes with Different Levels of Drought Resistance. J. Agron. Crop. Sci. 2008, 194, 15–25. [Google Scholar] [CrossRef]
- Parvin, S.; Uddin, S.; Bourgault, M.; Roessner, U.; Tausz-Posch, S.; Armstrong, R.; O’Leary, G.; Fitzgerald, G.; Tausz, M. Water availability moderates N2 fixation benefit from elevated [CO2]: A 2-year free-air CO2 enrichment study on lentil (Lens culinaris MEDIK.) in a water limited agroecosystem. Plant. Cell Environ. 2018, 41, 2418–2434. [Google Scholar] [CrossRef] [PubMed]
- Taketani, R.G.; Lançoni, M.D.; Kavamura, V.N.; Durrer, A.; Andreote, F.D.; Melo, I.S. Dry season constrains bacterial phylogenetic diversity in a semi-arid rhizosphere system. Microb. Ecol. 2017, 73, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Liu, J.; Chen, X.; Shu, H.; Lin, X.; Zhou, Q.; Bramryd, T.; Shu, W.; Huang, L. Microbial community structure and function in sediments from e-waste contaminated rivers at Guiyu area of China. Environ. Pollut. 2018, 235, 171–179. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Collecting Site | Geographic Co-Ordinates | District |
|---|---|---|---|
| Polhillia brevicalyx | Ghwarriekop farm | 34°33′53″S 19°59′43″E | Overberg |
| Polhillia pallens | Witkoppies farm | 34°33′53″S 19°59′43″E | Overberg |
| Wiborgia obcordata | Bushmans Kloof | 32°07′14″S 19°06′28″E | Cederberg |
| Wiborgia sericea | Traveller’s Rest farm | 32°04′15″S 19°04′32″E | Cederberg |
| Wiborgiella sessilifolia | Bredasdorp/Elim Pass | 34°37′58″S 19°49′399″E | Overberg |
| Dry Season | Wet Season | |||
|---|---|---|---|---|
| Plant Species | Soil Type | Elevation (m) | θm Water Content (%) | |
| Polhillia brevicalyx | Clay | 310 | 8.21 ± 0.00 | 25.47 ± 0.31 |
| Polhillia pallens | Clay | 152 | 9.26 ± 0.01 | 26.55 ± 0.06 |
| Wiborgia sericea | Sandy | 120 | 6.37 ± 0.01 | 20.00 ± 0.00 |
| Wiborgia obcordata | Sandy | 441 | 3.93 ± 0.05 | 11.90 ± 0.03 |
| Wiborgiella sessilifolia | Loam | 311 | 5.42 ± 0.03 | |
| pH and Minerals (mg/kg) | Polhillia brevicalyx | Polhilliapallens | Wiborgiella sessilifolia | Wiborgia obcordata | Wiborgiasericea |
|---|---|---|---|---|---|
| Concentrations | |||||
| pH(H2O) | 5.53 ± 0.03 | 4.45 ± 0.05 | 8.20 ± 0.00 | 4.33 ± 0.09 | 4.47 ± 0.09 |
| Magnesium | 4.55 ± 0.02 | 2.67 ± 0.01 | 3.08 ± 0.01 | 0.25 ± 0.01 | 0.27 ± 0.01 |
| Potassium | 170.18 ± 4.83 | 234.00 ± 1.00 | 53.67 ± 1.76 | 27.33 ± 1.76 | 37.00 ± 0.58 |
| Sodium | 110.22 ± 3.42 | 130.00 ± 0.00 | 64.33 ± 0.88 | 4.33 ± 0.33 | 9.00 ± 0.00 |
| Phosphorus | 22.00 ± 0.00 | 43.50 ± 0.50 | 15.67 ± 0.33 | 6.00 ± 0.58 | 8.33 ± 0.33 |
| Copper | 0.84 ± 0.00 | 0.46 ± 0.02 | 0.12 ± 0.01 | 0,14 ± 0.00 | 0.14 ± 0.1 |
| Zinc | 7.28 ± 0.05 | 2.84 ± 0.17 | 0.41 ± 0.01 | 0.18 ± 0.01 | 0.66 ± 0.03 |
| Boron | 10.67 ± 8.92 | 0.61 ± 0.01 | 0.80 ± 0.05 | 0.07 ± 0.00 | 0.12 ± 0.1 |
| Carbon (%) | 4.22 ± 0.13 | 7.76 ± 0.04 | 2.82 ± 0.15 | 0.39 ± 0.02 | 0.38 ± 0.02 |
| Iron | 105.85 ± 0.45 | 174.60 ± 6.20 | 46.98 ± 0.05 | 66.19 ± 2.88 | 129.2 ± 5.45 |
| NH4+ (%) | 0.40 ± 0.01 | 0.60 ± 0.00 | 0.67 ± 0.48 | 0.04 ± 0.01 | 0.03 ± 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mpai, T.; Jaiswal, S.K.; Cupido, C.N.; Dakora, F.D. Seasonal Effect on Bacterial Communities Associated with the Rhizospheres of Polhillia, Wiborgia and Wiborgiella Species in the Cape Fynbos, South Africa. Microorganisms 2022, 10, 1992. https://doi.org/10.3390/microorganisms10101992
Mpai T, Jaiswal SK, Cupido CN, Dakora FD. Seasonal Effect on Bacterial Communities Associated with the Rhizospheres of Polhillia, Wiborgia and Wiborgiella Species in the Cape Fynbos, South Africa. Microorganisms. 2022; 10(10):1992. https://doi.org/10.3390/microorganisms10101992
Chicago/Turabian StyleMpai, Tiisetso, Sanjay K. Jaiswal, Christopher N. Cupido, and Felix D. Dakora. 2022. "Seasonal Effect on Bacterial Communities Associated with the Rhizospheres of Polhillia, Wiborgia and Wiborgiella Species in the Cape Fynbos, South Africa" Microorganisms 10, no. 10: 1992. https://doi.org/10.3390/microorganisms10101992