Molecular Epidemiology Reveals Low Genetic Diversity among Cryptococcus neoformans Isolates from People Living with HIV in Lima, Peru, during the Pre-HAART Era

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Mating Type and Serotype Analysis
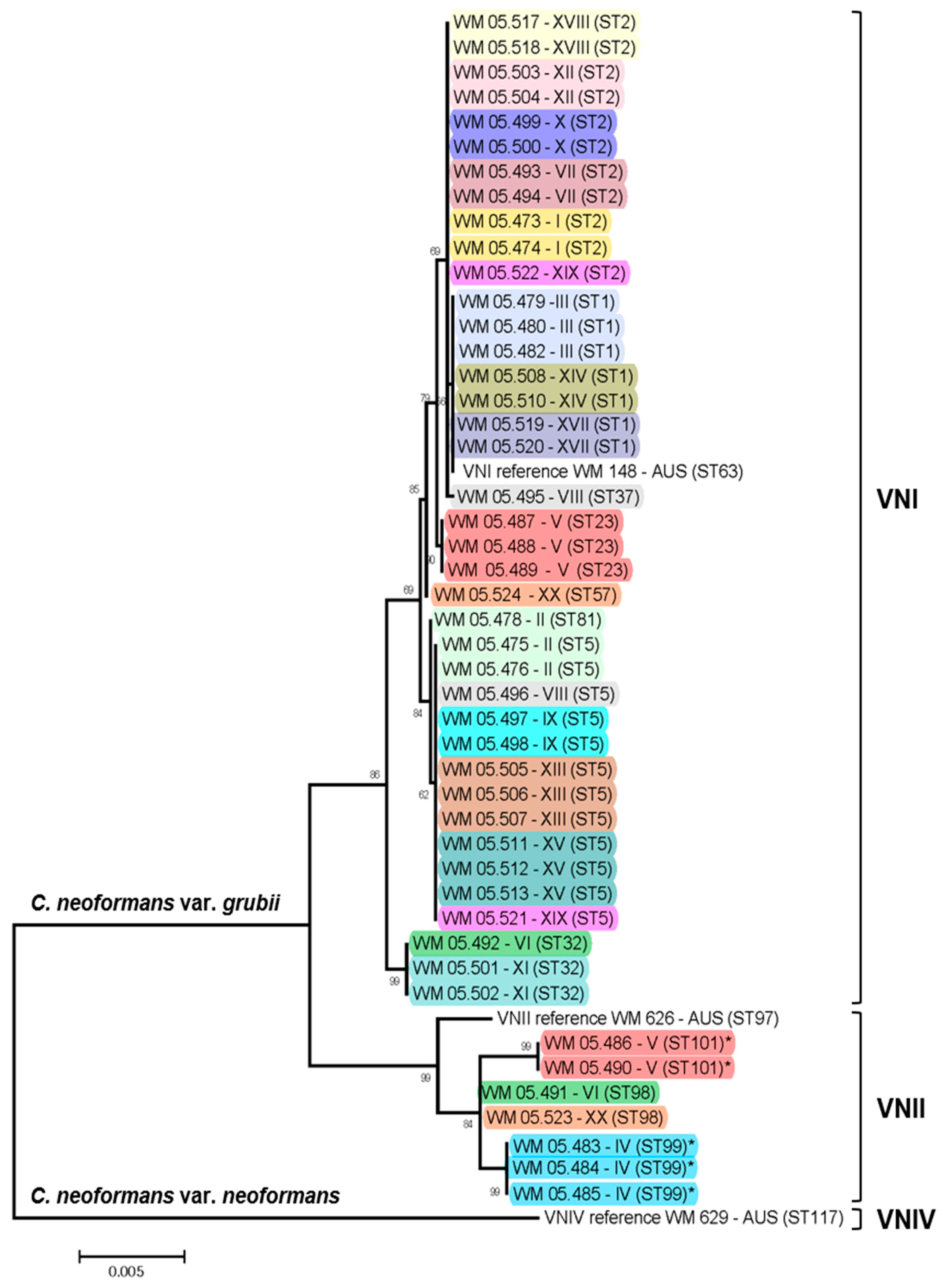
2.2. Major Molecular Type Determination
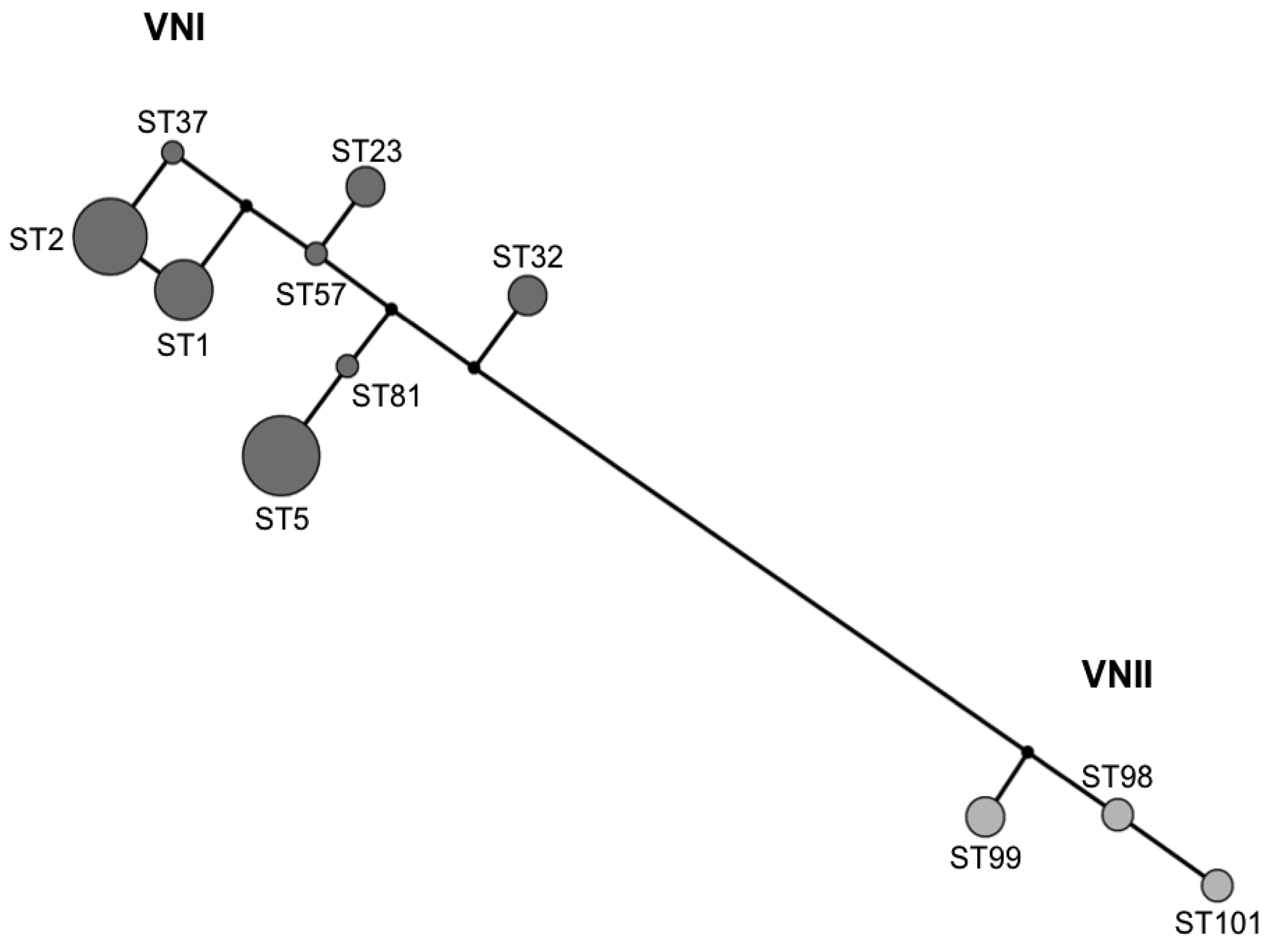
2.3. MLST Typing
2.4. Recombination and Clonality
3. Discussion
4. Materials and Methods
4.1. Isolates
4.2. DNA Extraction
4.3. Mating Type Determination
4.4. Serotyping
4.5. Molecular Typing
4.6. Recombination and Clonality
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kwon-Chung, K.J.; Fraser, J.A.; Doering, T.L.; Wang, Z.A.; Janbon, G.; Idnurm, A.; Bahn, Y.-S. Cryptococcus neoformans and cryptococcus gattii, the etiologic agents of cryptococcosis. Cold Spring Harb. Perspect. Med. 2014, 4, a019760. [Google Scholar] [CrossRef] [PubMed]
- Byrnes, E.J.; Bartlett, K.H.; Perfect, J.R.; Heitman, J. Cryptococcus gattii: An emerging fungal pathogen infecting humans and animals. Microbes Infect. 2011, 13, 895–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cogliati, M.; D’Amicis, R.; Zani, A.; Montagna, M.T.; Caggiano, G.; De Giglio, O.; Balbino, S.; De Donno, A.; Serio, F.; Susever, S.; et al. Environmental distribution of cryptococcus neoformans and c. Gattii around the mediterranean basin. FEMS Yeast Res. 2016, 16, fow086. [Google Scholar] [CrossRef] [PubMed]
- Boekhout, T.; Theelen, B.; Díaz, M.; Fell, J.W.; Hop, W.C.J.; Abeln, E.C.A.; Dromer, F.; Meyer, W. Hybrid genotypes in the pathogenic yeast cryptococcus neoformans. Microbiology 2001, 147, 891–907. [Google Scholar] [CrossRef] [Green Version]
- Meyer, W.; Castañeda, A.; Jackson, S.; Huynh, M.; Castañeda, E.; IberoAmerican Cryptococcal Study Group. Molecular typing of iberoamerican cryptococcus neoformans isolates. Emerg. Infect. Dis. 2003, 9, 189–195. [Google Scholar] [CrossRef]
- Meyer, W.; Marszewska, K.; Amirmostofian, M.; Igreja, R.P.; Hardtke, C.; Methling, K.; Viviani, M.A.; Chindamporn, A.; Sukroongreung, S.; John, M.A.; et al. Molecular typing of global isolates of cryptococcus neoformans var. Neoformans by polymerase chain reaction fingerprinting and randomly amplified polymorphic DNA-a pilot study to standardize techniques on which to base a detailed epidemiological survey. Electrophoresis 1999, 20, 1790–1799. [Google Scholar] [CrossRef]
- Meyer, W.; Aanensen, D.M.; Boekhout, T.; Cogliati, M.; Diaz, M.R.; Esposto, M.C.; Fisher, M.C.; Gilgado, F.; Hagen, F.; Kaocharoen, S.; et al. Consensus multi-locus sequence typing scheme for cryptococcus neoformans and cryptococcus gattii. Med. Mycol. 2009, 47, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Dromer, F.; Casadevall, A.; Perfect, J.; Sorrell, T. Cryptococcus neoformans: Latency and disease. In Cryptococcus: From Human Pathogen to Model Yeast; Heitman, J., Kozel, T., Kwon-Chung, K., Perfect, J., Eds.; American Society for Microbiology: Washington, DC, USA, 2011; pp. 431–440. [Google Scholar]
- Alanio, A.; Vernel-Pauillac, F.; Sturny-Leclere, A.; Dromer, F. Cryptococcus neoformans host adaptation: Toward biological evidence of dormancy. MBio 2015, 6, e02580-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musubire, A.K.; Boulware, D.R.; Meya, D.B.; Rhein, J. Diagnosis and management of cryptococcal relapse. J. AIDS Clin. Res. 2013, 3 (Suppl. 3), S3-003. [Google Scholar] [CrossRef]
- Desnos-Ollivier, M.; Patel, S.; Spaulding, A.R.; Charlier, C.; Garcia-Hermoso, D.; Nielsen, K.; Dromer, F. Mixed infections and in vivo evolution in the human fungal pathogen cryptococcus neoformans. MBio 2010, 1, e00091-10. [Google Scholar] [CrossRef] [Green Version]
- Guinea, J.; Hagen, F.; Peláez, T.; Boekhout, T.; Tahoune, H.; Torres-Narbona, M.; Bouza, E. Antifungal susceptibility, serotyping, and genotyping of clinical cryptococcus neoformans isolates collected during 18 years in a single institution in madrid, spain. Med. Mycol. 2010, 48, 942–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igreja, R.P.; Santos, L.M.D.; Wanke, B.; Gutierrez, G.M.C.; Kidd, S.E.; Meyer, W. Molecular epidemiology of cryptococcus neoformans isolates from aids patients of the brazilian city, rio de janeiro. Med. Mycol. 2004, 42, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mlinarić-Missoni, E.; Hagen, F.; Chew, W.H.M.; Važić-Babić, V.; Boekhout, T.; Begovac, J. In vitro antifungal susceptibilities and molecular typing of sequentially isolated clinical cryptococcus neoformans strains from croatia. J. Med. Microbiol. 2011, 60, 1487–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of hiv-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Pappas, P.G. Cryptococcal infections in non-hiv-infected patients. Trans. Am. Clin. Climatol. Assoc. 2013, 124, 61–79. [Google Scholar]
- Mitchell, T.G.; Perfect, J.R. Cryptococcosis in the era of aids--100 years after the discovery of cryptococcus neoformans. Clin. Microbiol. Rev. 1995, 8, 515–548. [Google Scholar] [CrossRef]
- DGE. Centro Nacional de Epidemiologia, Prevencion y Control de Enfermedades. 2020. Available online: https://www.dge.gob.pe/portal/index.php?option=com_content&view=article&id=656 (accessed on 21 April 2020).
- United Nations Programme on HIV and AIDS (UNAIDS). 2020. Available online: https://www.unaids.org/en/regionscountries/countries/peru (accessed on 21 April 2020).
- World Health Organization (WHO). 2020. Available online: https://www.who.int/gho/hiv/en/ (accessed on 21 April 2020).
- Dammert, P.; Bustamante, B.; Ticona, E.; Llanos-Cuentas, E.; Huaroto, L.; Chávez, V.; Campos, P. Treatment of cryptococcal meningitis in peruvian aids patients using amphotericin b and fluconazole. J. Infect. 2008, 57, 260–265. [Google Scholar] [CrossRef]
- Lofgren, S.M.; Abassi, M.; Rhein, J.; Boulware, D.R. Recent advances in aids-related cryptococcal meningitis treatment with an emphasis on resource limited settings. Expert Rev. Anti Infect. Ther. 2017, 15, 331–340. [Google Scholar] [CrossRef]
- Eza, D.; Cerrillo, G.; Moore, D.A.; Castro, C.; Ticona, E.; Morales, D.; Cabanillas, J.; Barrantes, F.; Alfaro, A.; Benavides, A.; et al. Postmortem findings and opportunistic infections in hiv-positive patients from a public hospital in peru. Pathol. Res. Pract. 2006, 202, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Litvintseva, A.P.; Thakur, R.; Vilgalys, R.; Mitchell, T.G. Mitchell. Multilocus sequence typing reveals three genetic subpopulations of cryptococcus neoformans var. Grubii (serotype a), including a unique population in botswana. Genetics 2005, 172, 2223–2238. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Paim, K.; Andrade-Silva, L.; Fonseca, F.M.; Ferreira, T.B.; Mora, D.J.; Andrade-Silva, J.; Khan, A.; Dao, A.; Reis, E.C.; Almeida, M.T.G.; et al. Mlst-based population genetic analysis in a global context reveals clonality amongst cryptococcus neoformans var. Grubii vni isolates from hiv patients in southeastern brazil. PLoS Negl. Trop. Dis. 2017, 11, e0005223. [Google Scholar] [CrossRef] [PubMed]
- Litvintseva, A.P.; Mitchell, T.G. Most environmental isolates of cryptococcus neoformans var. Grubii (serotype a) are not lethal for mice. Infect. Immun. 2009, 77, 3188–3195. [Google Scholar] [CrossRef] [Green Version]
- Rocha, D.F.S.; Cruz, K.S.; Santos, C.S.D.S.; Menescal, L.S.F.; Neto, J.R.D.S.; Pinheiro, S.B.; Silva, L.M.; Trilles, L.; De Souza, J.V.B. Mlst reveals a clonal population structure for cryptococcus neoformans molecular type vni isolates from clinical sources in amazonas, northern-brazil. PLoS ONE 2018, 13, e0197841. [Google Scholar] [CrossRef] [Green Version]
- Sanchini, A.; Smith, I.M.; Sedlacek, L.; Schwarz, R.; Tintelnot, K.; Rickerts, V. Molecular typing of clinical cryptococcus neoformans isolates collected in germany from 2004 to 2010. Med. Microbiol. Immunol. 2014, 203, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Van Wyk, M.; Govender, N.P.; Mitchell, T.G.; Litvintseva, A.P.; Sa, G. Multilocus sequence typing of serially collected isolates of cryptococcus from hiv-infected patients in south africa. J. Clin. Microbiol. 2014, 52, 1921–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélez, N.; Alvarado, M.; Parra-Giraldo, C.M.; Sánchez-Quitian, Z.A.; Escandón, P.; Castañeda, E. Genotypic diversity is independent of pathogenicity in colombian strains of cryptococcus neoformans and cryptococcus gattii in galleria mellonella. J. Fungi (Basel) 2018, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Danesi, P.; Firacative, C.; Cogliati, M.; Otranto, D.; Capelli, G.; Meyer, W. Multilocus sequence typing (mlst) and m13 pcr fingerprinting revealed heterogeneity amongst cryptococcus species obtained from italian veterinary isolates. FEMS Yeast Res. 2014, 14, 897–909. [Google Scholar] [CrossRef]
- Umeyama, T.; Ohno, H.; Minamoto, F.; Takagi, T.; Tanamachi, C.; Tanabe, K.; Kaneko, Y.; Yamagoe, S.; Kishi, K.; Fujii, T.; et al. Determination of epidemiology of clinically isolated cryptococcus neoformans strains in japan by multilocus sequence typing. Jpn. J. Infect. Dis. 2013, 66, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Kim, M.; Joo, S.I.; Hwang, S.M. Molecular epidemiology of clinical cryptococcus neoformans isolates in seoul, korea. Mycobiology 2014, 42, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Kaocharoen, S.; Ngamskulrungroj, P.; Firacative, C.; Trilles, L.; Piyabongkarn, D.; Banlunara, W.; Poonwan, N.; Chaiprasert, A.; Meyer, W.; Chindamporn, A. Molecular epidemiology reveals genetic diversity amongst isolates of the cryptococcus neoformans/c. Gattii species complex in thailand. PLoS Negl. Trop. Dis. 2013, 7, e2297. [Google Scholar] [CrossRef] [Green Version]
- Wiesner, D.L.; Moskalenko, O.; Corcoran, J.M.; McDonald, T.; Rolfes, M.A.; Njama-Meya, D.; Kajumbula, H.; Kambugu, A.; Bohjanen, P.R.; Knight, J.F.; et al. Cryptococcal genotype influences immunologic response and human clinical outcome after meningitis. MBio 2012, 3, e00196-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, L.M.; Meyer, W.; Firacative, C.; Thompson, G.R.; Samitz, E.; Sykes, J.E. Antifungal drug susceptibility and phylogenetic diversity among cryptococcus isolates from dogs and cats in north america. J. Clin. Microbiol. 2014, 52, 2061–2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, L.E.D.A.; Ferreira-Paim, K.; Ferreira, T.B.; Vilas-Boas, A.; Mora, D.J.; Manzato, V.M.; Fonseca, F.M.; Buosi, K.; Andrade-Silva, J.; Prudente, B.D.S.; et al. Genotypic analysis of clinical and environmental cryptococcus neoformans isolates from brazil reveals the presence of vnb isolates and a correlation with biological factors. PLoS ONE 2018, 13, e0193237. [Google Scholar] [CrossRef] [Green Version]
- Day, J.; Qihui, S.; Thanh, L.T.; Trieu, P.H.; Van, A.D.; Thu, N.H.; Chau, T.T.H.; Lan, N.P.H.; Chau, N.V.V.; Ashton, P.M.; et al. Comparative genomics of cryptococcus neoformans var. Grubii associated with meningitis in hiv infected and uninfected patients in vietnam. PLoS Negl. Trop. Dis. 2017, 11, e0005628. [Google Scholar] [CrossRef] [PubMed]
- Khayhan, K.; Hagen, F.; Pan, W.; Simwami, S.; Fisher, M.C.; Wahyuningsih, R.; Chakrabarti, A.; Chowdhary, A.; Ikeda, R.; Taj-Aldeen, S.J.; et al. Geographically structured populations of cryptococcus neoformans var. Grubii in asia correlate with hiv status and show a clonal population structure. PLoS ONE 2013, 8, e72222. [Google Scholar] [CrossRef]
- Cogliati, M.; Zamfirova, R.R.; Tortorano, A.M.; Viviani, M.A. Molecular epidemiology of italian clinical cryptococcus neoformans var. Grubii isolates. Med. Mycol 2013, 51, 499–506. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.H.; Ngamskulrungroj, P.; Varma, A.; Sionov, E.; Hwang, S.M.; Carriconde, F.; Meyer, W.; Litvintseva, A.P.; Lee, W.G.; Shin, J.H.; et al. Prevalence of the vnic genotype of cryptococcus neoformans in non-hiv-associated cryptococcosis in the republic of korea. FEMS Yeast Res. 2010, 10, 769–778. [Google Scholar] [CrossRef]
- Dou, H.; Wang, H.; Xie, S.; Chen, X.; Xu, Z.; Xu, Y. Molecular characterization of cryptococcus neoformans isolated from the environment in beijing, china. Med. Mycol. 2017, 55, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Sorrell, T.C.; Nimmo, G.; Speed, B.; Currie, B.; Ellis, D.; Marriott, D.; Pfeiffer, T.; Parr, D.; Byth, K. Epidemiology and host- and variety-dependent characteristics of infection due to cryptococcus neoformans in australia and new zealand. Australasian cryptococcal study group. Clin. Infect. Dis. 2000, 31, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Latin American Cryptococcal Study Group. The status of cryptococcosis in latin america. Meml. Inst. Oswaldo Cruz 2018, 113, e170554. [Google Scholar] [CrossRef] [Green Version]
- Cherniak, R.; Morris, L.C.; Belay, T.; Spitzer, E.D.; Casadevall, A. Variation in the structure of glucuronoxylomannan in isolates from patients with recurrent cryptococcal meningitis. Infect. Immun. 1995, 63, 1899–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon-Chung, K.J.; Varma, A. Do major species concepts support one, two or more species within cryptococcus neoformans? FEMS Yeast Res. 2006, 6, 574–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, M.E.; Pfaller, M.A.; Hajjeh, R.A.; Graviss, E.A.; Rees, J.; Spitzer, E.D.; Pinner, R.W.; Mayer, L.W. Molecular subtypes and antifungal susceptibilities of serial cryptococcus neoformans isolates in human immunodeficiency virus-associated cryptococcosis. Cryptococcal disease active surveillance group. J. Infect. Dis. 1996, 174, 812–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illnait-Zaragozi, M.T.; Martinez-Machin, G.F.; Fernandez-Andreu, C.M.; Hagen, F.; Boekhout, T.; Klaassen, C.H.; Meis, J.F. Microsatellite typing and susceptibilities of serial cryptococcus neoformans isolates from cuban patients with recurrent cryptococcal meningitis. BMC Infect. Dis. 2010, 10, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litvintseva, A.P.; Marra, R.E.; Nielsen, K.; Heitman, J.; Vilgalys, R.; Mitchell, T.G. Evidence of sexual recombination among cryptococcus neoformans serotype a isolates in sub-saharan africa. Eukaryottic 2003, 2, 1162–1168. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, C.; Colom, F.; Frases, S.; Mulet, E.; Abad, J.L.; Alio, J.L. Detection and identification of fungal pathogens by pcr and by its2 and 5.8s ribosomal DNA typing in ocular infections. J. Clin. Microbiol. 2001, 39, 2873–2879. [Google Scholar] [CrossRef] [Green Version]
- Halliday, C.L.; Bui, T.; Krockenberger, M.; Malik, R.; Ellis, D.H.; Carter, D.A. Presence of alpha and a mating types in environmental and clinical collections of cryptococcus neoformans var. Gattii strains from australia. J. Clin. Microbiol. 1999, 37, 2920–2926. [Google Scholar]
- Enache-Angoulvant, A.; Chandenier, J.; Symoens, F.; Lacube, P.; Bolognini, J.; Douchet, C.; Poirot, J.L.; Hennequin, C. Molecular identification of cryptococcus neoformans serotypes. J. Clin. Microbiol. 2007, 45, 1261–1265. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Stecher, G.; Nei, M.; Kumar, S.; Peterson, N. Mega5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Agapow, P.-M.; Burt, A. Indices of multilocus linkage disequilibrium. Mol. Ecol. Notes 2001, 1, 101–102. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Patient Number | WM Number | Isolation Point | Date of Isolation | Source a | Age/Gender | URA5-RFLP and PCR fingerprinting | AFLP Type | Mating Type/Serotype | CAP59 | GPD1 | IGS1 | LAC1 | PLB1 | SOD1 | URA5 | Sequence Type (ST) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | WM 05.473 | Relapse | 23/06/97 | CSF | 26/M | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 |
| WM 05.474 | Relapse | 27/10/98 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | ||
| II | WM 05.475 | Baseline | 20/10/99 | CSF | 74/M | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 |
| WM 05.478 | Failure | 19/07/00 | CSF | VNI | I | alpha/A | 1 | 1 | 1 | 5 | 2 | 1 | 1 | 81 | ||
| WM 05.476 | Failure | 30/10/00 | CSF | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 | ||
| III | WM 05.479 | Baseline | 29/11/99 | CSF | 26/M | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| WM 05.480 | Failure | 16/06/00 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| WM 05.482 | Failure | 17/08/00 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| IV | WM 05.483 | Baseline | 18/01/00 | CSF | VNII | II | alpha/A | 2 | 9 | 38 | 11 | 11 | 16 | 15 | 99 * | |
| WM 05.484 | Relapse | 28/04/00 | CSF | 23/F | VNII | II | alpha/A | 2 | 9 | 38 | 11 | 11 | 16 | 15 | 99 * | |
| WM 05.485 | Failure | 3/06/00 | CSF | VNII | II | alpha/A | 2 | 9 | 38 | 11 | 11 | 16 | 15 | 99 * | ||
| V | WM 05.486 | Baseline | 13/03/00 | Sputum b | VNII | II | alpha/A | 2 | 10 | 39 | 8 | 12 | 16 | 33 | 101 * | |
| WM 05.487 | Baseline | 11/09/00 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 2 | 1 | 1 | 2 | 23 | ||
| WM 05.488 | Baseline | 13/09/00 | Sputum b | 28/M | VNI | I | alpha/A | 7 | 1 | 1 | 2 | 1 | 1 | 2 | 23 | |
| WM 05.489 | 7th day treatment | 18/09/00 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 2 | 1 | 1 | 2 | 23 | ||
| WM 05.490 | 14th day treatment | 25/09/00 | CSF | VNII | II | alpha/A | 2 | 10 | 39 | 8 | 12 | 16 | 33 | 101 * | ||
| VI | WM 05.491 | Baseline | 2/01/01 | CSF | 38/M | VNII | II | alpha/A | 2 | 9 | 14 | 8 | 11 | 16 | 4 | 98 |
| WM 05.492 | Relapse | 15/06/01 | CSF | VNI | I | alpha/A | 1 | 1 | 10 | 3 | 4 | 1 | 1 | 32 | ||
| VII | WM 05.493 | Baseline | 20/06/01 | CSF | 22/F | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 |
| WM 05.494 | Failure | 21/09/01 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | ||
| VIII | WM 05.495 | Baseline | 28/10/00 | CSF | 52/F | VNI | I | alpha/A | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 37 |
| WM 05.496 | Relapse | 6/09/01 | CSF | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 | ||
| IX | WM 05.497 | Baseline | 23/08/01 | CSF | 40/F | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 |
| WM 05.498 | Failure | 28/01/02 | CSF | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 | ||
| X | WM 05.499 | Baseline | 19/05/98 | CSF | 35/M | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 |
| WM 05.500 | Failure | 5/08/98 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | ||
| XI | WM 05.501 | Baseline | 24/08/98 | CSF | 27/M | VNI | I | alpha/A | 1 | 1 | 10 | 3 | 4 | 1 | 1 | 32 |
| WM 05.502 | Relapse | 26/10/99 | CSF | VNI | I | alpha/A | 1 | 1 | 10 | 3 | 4 | 1 | 1 | 32 | ||
| XII | WM 05.503 | Baseline | 16/10/98 | CSF | 23/M | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 |
| WM 05.504 | Failure | 26/12/98 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | ||
| XIII | WM 05.505 | Baseline | 12/11/98 | CSF | 22/M | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 |
| WM 05.506 | Failure | 22/01/99 | CSF | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 | ||
| WM 05.507 | Failure | 23/03/99 | CSF | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 | ||
| XIV | WM 05.508 | Baseline | 15/01/99 | CSF | 32/M | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| WM 05.510 | Failure | 26/03/99 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| XV | WM 05.511 | Baseline | 3/03/99 | CSF | 37/M | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 |
| WM 05.513 | 14th day treatment | 18/03/99 | CSF | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 | ||
| WM 05.512 | Failure | 13/05/99 | CSF | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 | ||
| XVI | WM 05.515 | Baseline | 29/04/99 | CSF | 28/M | VNIII | III | alpha/AD c | ||||||||
| WM 05.516 | Failure | 8/07/99 | CSF | VNIII | III | alpha/AD c | ||||||||||
| XVII | WM 05.519 | Baseline | 20/05/00 | CSF | 26/M | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| WM 05.520 | Relapse | 9/08/00 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| XVIII | WM 05.517 | Baseline | 10/08/00 | CSF | 47/F | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 |
| WM 05.518 | Relapse | 11/01/01 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | ||
| XIX | WM 05.521 | Baseline | 30/12/00 | CSF | 29/F | VNI | I | alpha/A | 1 | 3 | 1 | 5 | 2 | 1 | 1 | 5 |
| WM 05.522 | Relapse | 1/06/01 | CSF | VNI | I | alpha/A | 7 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | ||
| XX | WM 05.523 | Baseline | 5/03/01 | CSF | 41/M | VNII | II | alpha/A | 2 | 9 | 14 | 8 | 11 | 16 | 4 | 98 |
| WM 05.524 | Relapse | 1/06/02 | CSF | VNI | I | alpha/A | 1 | 1 | 1 | 3 | 1 | 1 | 1 | 57 |
| Molecular Type | Sequence Type (ST) | Country | Reference |
|---|---|---|---|
| VNI | ST1 | France a, Peru, USA, | [24] |
| VNI | ST2 | Argentina, Brazil, Colombia, France, Germany, Malawi, Peru, South Africa, Tanzania, USA | [24,25,26,27,28,29,30] |
| VNI | ST5 | Belgium, Brazil, Colombia, France, Germany, Italy, China, Japan, Korea, Thailand, Kuwait, Peru, Qatar, South Africa, Uganda, Malawi, USA b, Vietnam | [11,24,25,27,28,29,30,31,32,33,34,35,36,37,38,39] |
| VNI | ST23 | Belgium a, Brazil, Colombia, France a, Italy, Germany, Japan a, Kuwait, Korea, Peru, South Africa, Uganda, USA a,b | [11,24,25,28,29,30,36,39,40,41] |
| VNI | ST32 | Brazil, Belgium, China, Colombia, Germany, Japan, Peru, South Africa, USA b, Tanzania, Thailand, Uganda, Vietnam, Zaire | [24,28,29,30,32,36,38] |
| VNI | ST37 | Italy, Peru | [31] |
| VNI | ST57 | China, Peru | [42] |
| VNI | ST81 | Italy, Peru, Thailand | [31,34] |
| VNII | ST98 | Mexico, Peru | [5] |
| VNII | ST99 | Peru | Current Study |
| VNII | ST101 | Peru | Current Study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van de Wiele, N.; Neyra, E.; Firacative, C.; Gilgado, F.; Serena, C.; Bustamante, B.; Meyer, W. Molecular Epidemiology Reveals Low Genetic Diversity among Cryptococcus neoformans Isolates from People Living with HIV in Lima, Peru, during the Pre-HAART Era. Pathogens 2020, 9, 665. https://doi.org/10.3390/pathogens9080665
van de Wiele N, Neyra E, Firacative C, Gilgado F, Serena C, Bustamante B, Meyer W. Molecular Epidemiology Reveals Low Genetic Diversity among Cryptococcus neoformans Isolates from People Living with HIV in Lima, Peru, during the Pre-HAART Era. Pathogens. 2020; 9(8):665. https://doi.org/10.3390/pathogens9080665
Chicago/Turabian Stylevan de Wiele, Nathalie, Edgar Neyra, Carolina Firacative, Felix Gilgado, Carolina Serena, Beatriz Bustamante, and Wieland Meyer. 2020. "Molecular Epidemiology Reveals Low Genetic Diversity among Cryptococcus neoformans Isolates from People Living with HIV in Lima, Peru, during the Pre-HAART Era" Pathogens 9, no. 8: 665. https://doi.org/10.3390/pathogens9080665