Effect of Ethanol Extracts of Propolis (EEPs) against Staphylococcal Biofilm—Microscopic Studies

 , ,
, ,
Abstract
:1. Introduction
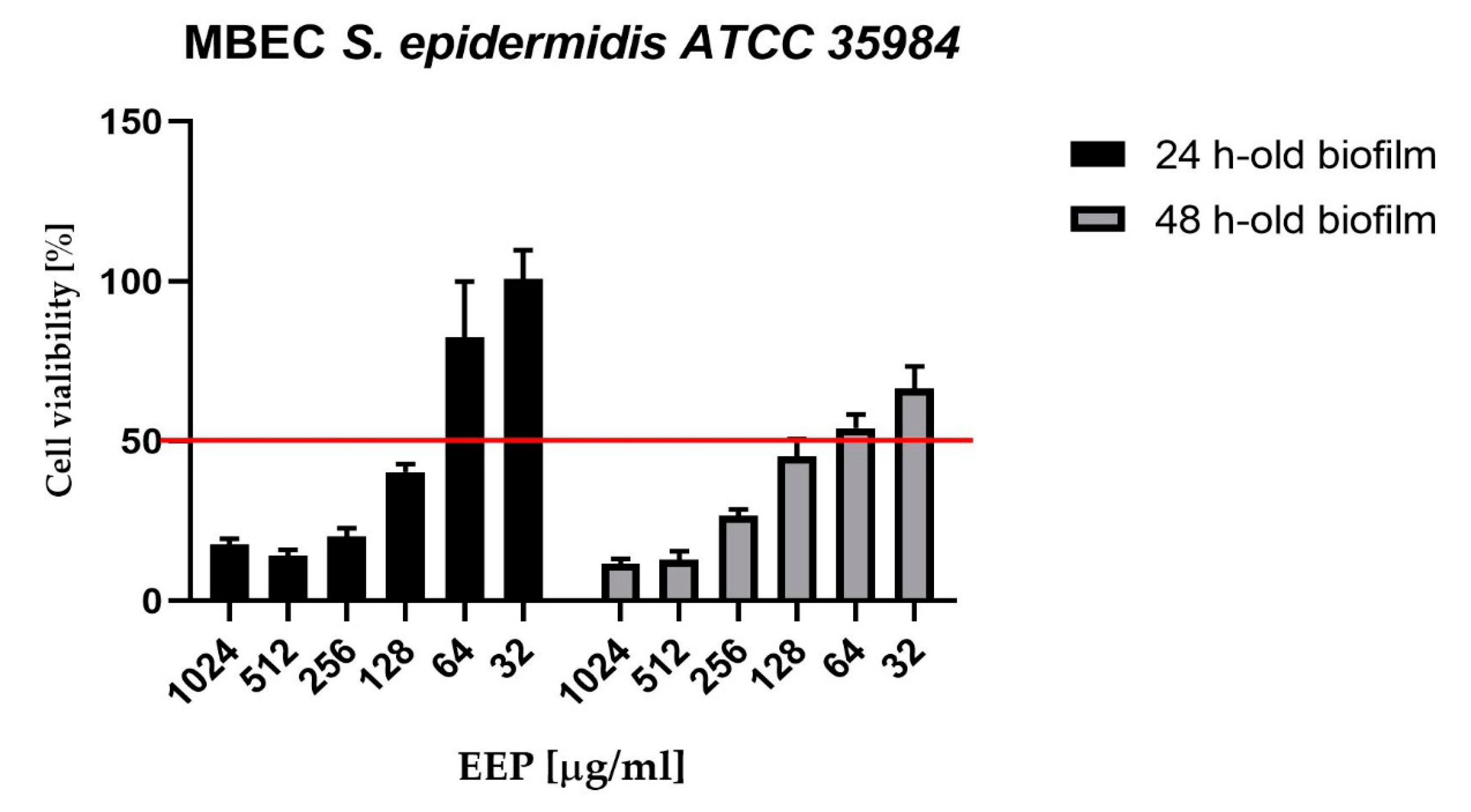
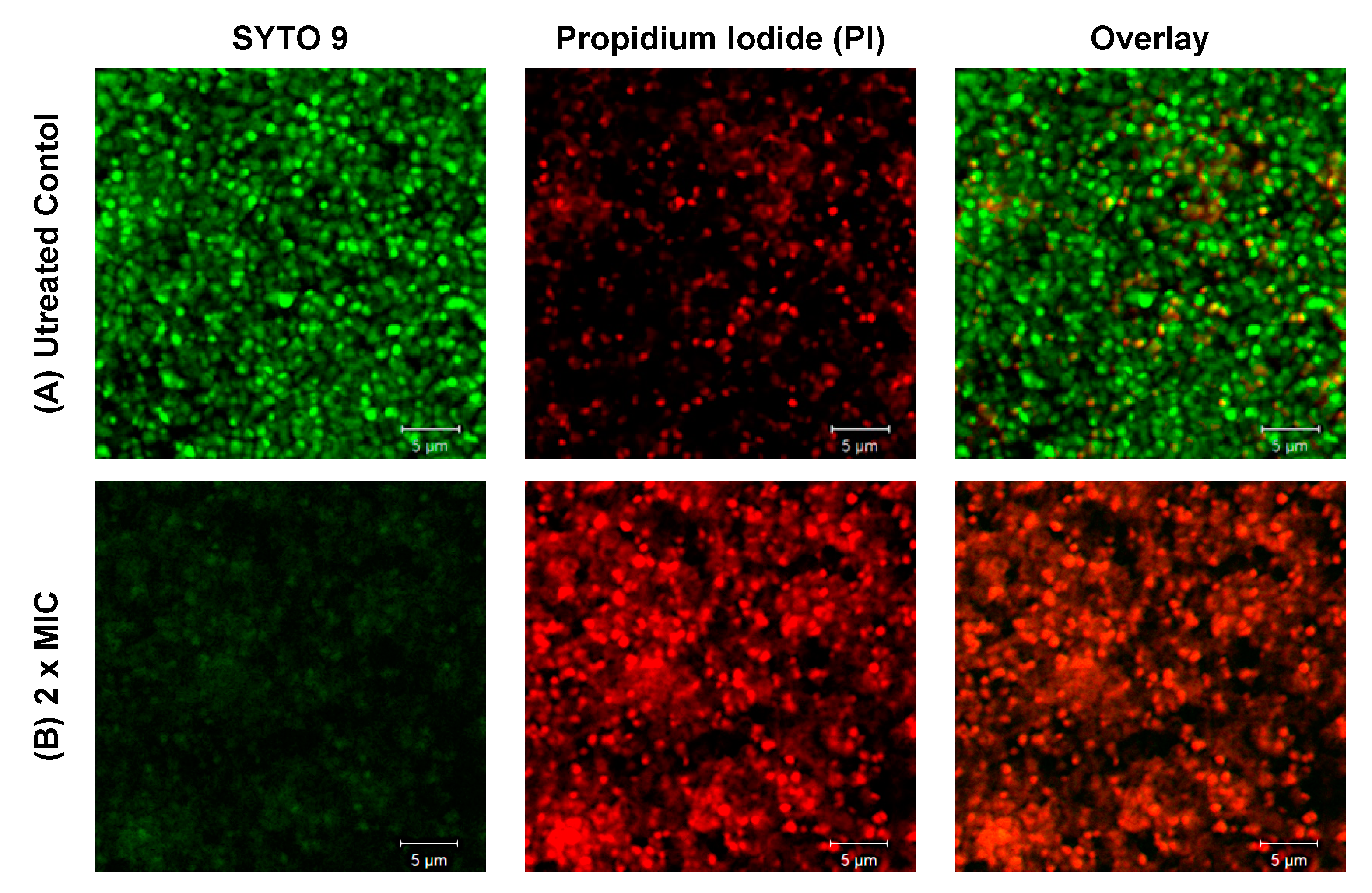
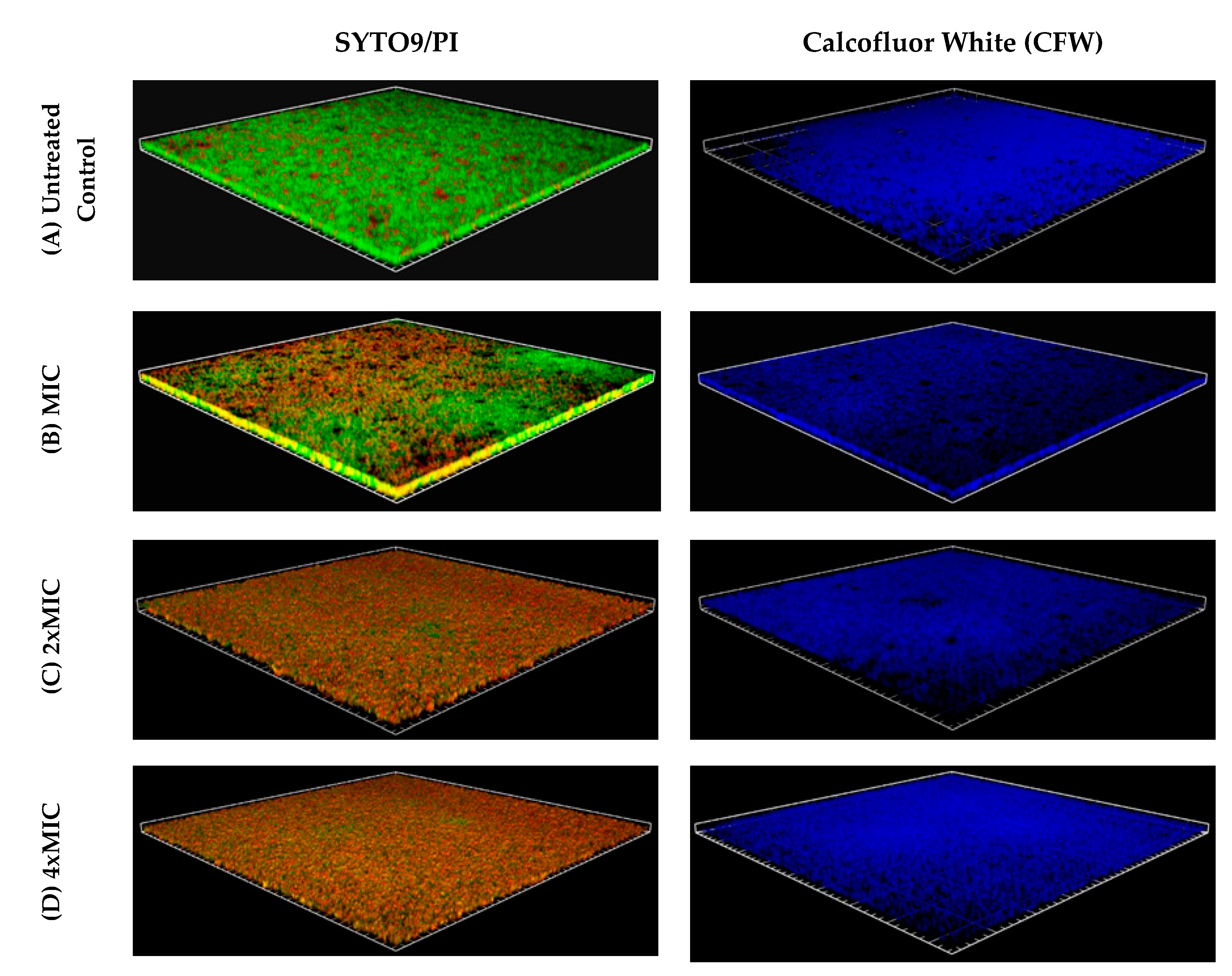
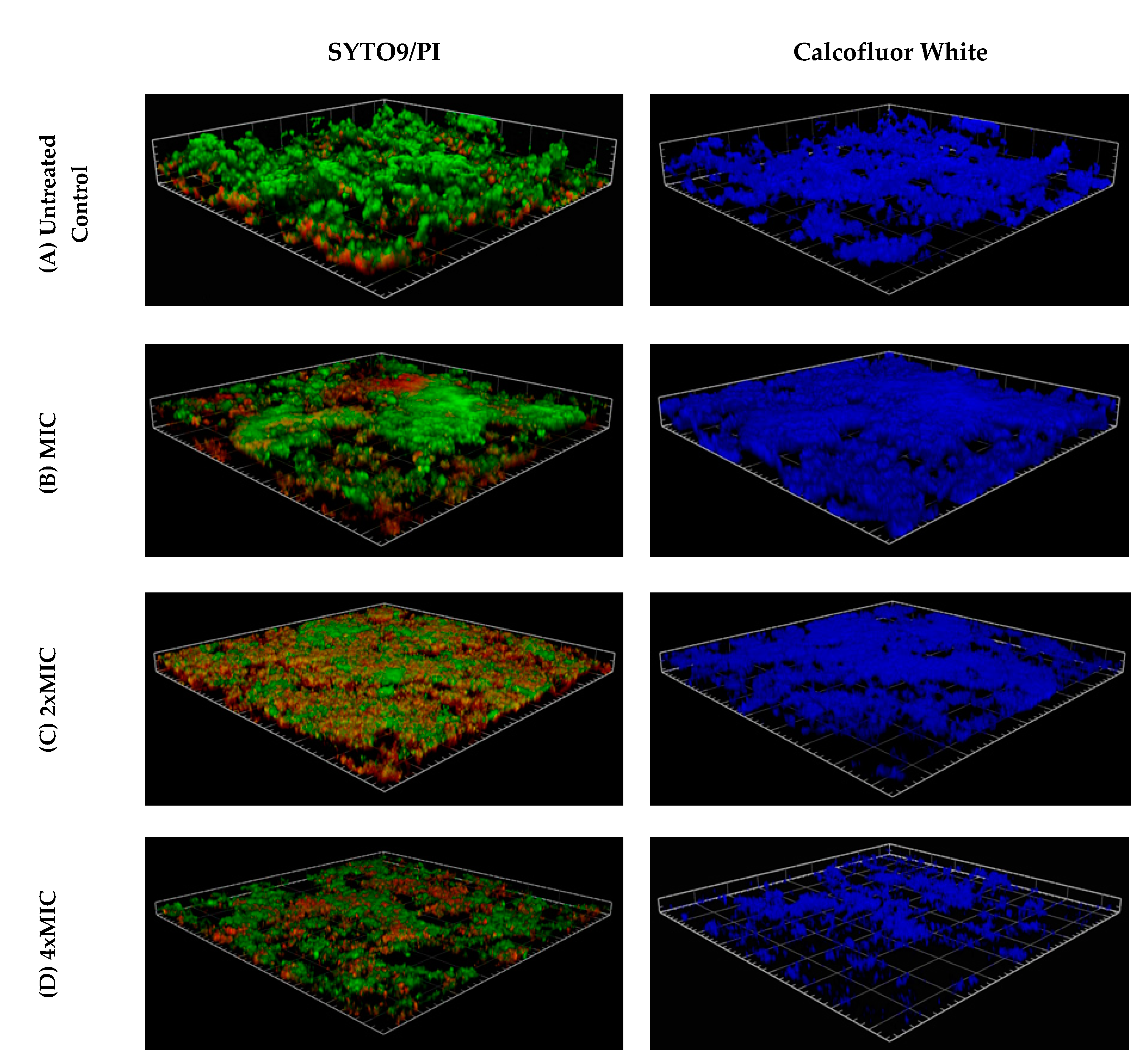
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Media
4.2. Propolis Extract Preparation
4.3. Total Phenolic Content
4.4. Minimum Inhibitory Concentration
4.5. Biofilm Formation
4.5.1. Inoculum Preparation
4.5.2. MBEC Assay
4.5.3. Biofilm Formation in an 8-Well Chamber Coverslip
4.5.4. Biofilm Formation on Aluminum Coupons
4.6. Biofilm Visualization: Confocal Microscopy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lacey, K.A.; Geoghegan, J.A.; McLoughlin, R.M. The role of Staphylococcus aureus virulence factors in skin infection and their potential as vaccine antigens. Pathogens 2016, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, T.L.; Carothers, K.E.; Lee, S.W. Virulence factor targeting of the bacterial pathogen Staphylococcus aureus for vaccine and therapeutics. Curr. Drug Targets. 2018, 19, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, C.; Ziebuhr, W.; Becker, K. Are coagulase-negative staphylococci virulent? Clin. Microbiol. Infect. 2019, 25, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal biofilm. Microbiol Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Resch, A.; Rosenstein, R.; Nerz, C.; Götz, F. Differential gene expression profiling of Staphylococcus aureus cultivated under biofilm and planktonic conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Sturdevant, D.E.; Otto, M. Genomewide analysis of gene expression in Staphylococcus epidermidis biofilms: Insights into the pathophysiology of S. epidermidis biofilms and the role of phenol-soluble modulins in formation of biofilms. J. Infect. Dis. 2005, 191, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Ricciardi, B.F.; Muthukrishnan, G.; Masters, E.; Ninomiya, M.; Lee, C.C.; Schwarz, E.M. Staphylococcus aureus evasion of host immunity in the setting of prosthetic joint infection: Biofilm and beyond. Curr. Rev. Musculoskelet. Med. 2018, 11, 389–400. [Google Scholar] [CrossRef]
- Mah, T.F.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Melchior, M.B.; Fink-Gremmels, J.; Gaastra, W. Comparative assessment of the antimicrobial susceptibility of Staphylococcus aureus isolates from bovine mastitis in biofilm versus planktonic culture. J. Vet. Med. Series B. 2006, 53, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Zvarych, V.; Stasevych, M.; Novikov, V.; Rusanov, E.; Vovk, M.; Szweda, P.; Grecka, K.; Milewski, S. Anthra [1,2-d][1,2,3]triazine-4,7,12(3H)-triones as a new class of antistaphylococcal agents: Synthesis and biological evaluation. Molecules 2019, 24, 4581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Belkum, A.; Melles, D.C.; Nouwen, J.; van Leeuwen, W.B.; van Wamel, W.; Vos, M.C.; Wertheim, H.F.; Verbrugh, H.A. Co-evolutionary aspects of human colonisation and infection by Staphylococcus aureus. Infect. Genet. Evol. 2009, 9, 32–47. [Google Scholar] [CrossRef] [PubMed]
- Munckhof, W.J.; Nimmo, G.R.; Schooneveldt, J.M.; Schlebusch, S.; Stephens, A.J.; Williams, G.; Huygens, F.; Giffard, P. Nasal carriage of Staphylococcus aureus, including community-associated methicillin-resistant strains, in Queensland adults. Clin. Microbiol Infect. 2009, 15, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Kloos, W.E.; Musselwhite, M.S. Distribution and persistence of Staphylococcus and Micrococcus species and other aerobic bacteria on human skin. Appl. Microbiol. 1975, 30, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Otto, M. Staphylococcus epidermidis the ‘accidental’ pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Uçkay, I.; Pittet, D.; Vaudaux, P.; Sax, H.; Lew, D.; Waldvogel, F. Foreign body infections due to Staphylococcus epidermidis. Ann. Med. 2009, 41, 109–119. [Google Scholar] [CrossRef]
- Yong, Y.Y.; Dykes, G.A.; Choo, W.S. Biofilm formation by staphylococci in health-related environments and recent reports on their control using natural compounds. Crit. Rev. Microbiol. 2019, 45, 201–222. [Google Scholar] [CrossRef]
- Grecka, K.; Kuś, P.M.; Okińczyc, P.; Worobo, R.W.; Walkusz, J.; Szweda, P. The anti-staphylococcal potential of ethanolic polish propolis extracts. Molecules 2019, 24, 1732. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.L.; Bloebaum, R.D. Observing the biofilm matrix of Staphylococcus epidermidis ATCC 35984 grown using the CDC biofilm reactor. Microsc. Microanal. 2010, 16, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Manukumar, H.M.; Rakesh, K.P.; Karthik, C.S.; Mallu, P.; Qin, H.-L.; Hussein Eissa, M.Y. Vision for medicine: Staphylococcus aureus biofilm war and unlocking key’s for anti-biofilm drug development. Microb. Pathog. 2018, 123, 339–347. [Google Scholar] [CrossRef]
- Craft, K.M.; Nguyen, J.M.; Berg, L.J. Townsend SD. Methicillin-resistant Staphylococcus aureus (MRSA): Antibiotic-resistance and the biofilm phenotype. Medchemcomm 2019, 10, 1231–1241. [Google Scholar] [CrossRef]
- Wojtyczka, R.D.; Kępa, M.; Idzik, D.; Kubina, R.; Kabała-Dzik, A.; Dziedzic, A.; Wąsik, T.J. In Vitro Antimicrobial activity of ethanolic extract of polish propolis against biofilm-forming Staphylococcus epidermidis strains. Evid. Based Complement. Alternat. Med. 2013, 2013, 590703. [Google Scholar] [CrossRef] [Green Version]
- Bryan, J.; Redden, P.; Traba, C. The mechanism of action of Russian propolis ethanol extracts against two antibiotic-resistant biofilm-forming bacteria. Lett. Appl. Microbiol. 2016, 62, 192–198. [Google Scholar] [CrossRef]
- Ambi, A.; Bryan, J.; Borbon, K.; Centeno, D.; Liu, T.; Chen, T.P.; Cattabiani, T.; Traba, C. Are Russian propolis ethanol extracts the future for the prevention of medical and biomedical implant contaminations? Phytomedicine 2017, 30, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Akca, A.E.; Akca, G.; Topçu, F.T.; Macit, E.; Pikdöken, L.; Özgen, I.Ş. The Comparative evaluation of the antimicrobial effect of propolis with chlorhexidine against oral pathogens: An in vitro study. Biomed. Res. Int. 2016, 2016, 3627463. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Dembogurski, D.S.; Silva Trentin, D.; Boaretto, A.G.; Rigo, G.V.; da Silva, R.C.; Tasca, T.; Macedo, A.J.; Carollo, C.A.; Silva, D.B. Brown propolis-metabolomic innovative approach to determine compounds capable of killing Staphylococcus aureus biofilm and Trichomonas vaginalis. Food Res. Int. 2018, 111, 661–673. [Google Scholar] [CrossRef] [PubMed]
- El-Guendouz, S.; Aazza, S.; Lyoussi, B.; Bankova, V.; Popova, M.; Neto, L.; Faleiro, M.L.; Miguel, M.D.G. Moroccan propolis: A natural antioxidant, antibacterial, and antibiofilm against Staphylococcus aureus with no induction of resistance after continuous exposure. Evid. Based Complement. Alternat. Med. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Stan, T.; Mărutescu, L.; Chifiriuc, M.C.; Lazăr, V. Anti-pathogenic effect of propolis extracts from different Romanian regions on Staphylococcus sp. clinical strains. Rom. Biotechnol. Lett. 2016, 21, 11166–11175. [Google Scholar]
- de Mélo Silva, I.S.; do Amorim Costa Gaspar, L.M.; Rocha, A.M.O.; da Costa, L.P.; Tada, D.B.; Franceschi, E.; Padilha, F.F. Encapsulation of red propolis in polymer nanoparticles for the destruction of pathogenic biofilms. AAPS PharmSciTech. 2020, 21, 49. [Google Scholar] [CrossRef]
- De Marco, S.; Piccioni, M.; Pagiotti, R.; Pietrella, D. Antibiofilm and antioxidant activity of propolis and bud poplar resins versus Pseudomonas aeruginosa. Evid. Based Complement. Alternat. Med. 2017, 2017, 5163575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meto, A.; Colombari, B.; Meto, A.; Boaretto, G.; Pinetti, D.; Marchetti, L.; Benvenuti, S.; Pellati, F.; Blasi, E. Propolis affects Pseudomonas aeruginosa growth, biofilm formation, eDNA release and phenazine production: Potential involvement of polyphenols. Microorganisms 2020, 8, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobaldini-Valerio, F.K.; Bonfim-Mendonça, P.S.; Rosseto, H.C.; Bruschi, M.L.; Henriques, M.; Negri, M.; Silva, S.; Svidzinski, T.L. Propolis: A potential natural product to fight Candida species infections. Future Microbiol. 2016, 11, 1035–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gucwa, K.; Kusznierewicz, B.; Milewski, S.; Van Dijck, P.; Szweda, P. Antifungal activity and synergism with azoles of polish propolis. Pathogens 2018, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Martorano-Fernandes, L.; Cavalcanti, Y.W.; de Almeida, L.F.D. Inhibitory effect of Brazilian red propolis on Candida biofilms developed on titanium surfaces. BMC Complement. Med. Ther. 2020, 20, 104. [Google Scholar] [CrossRef] [Green Version]
- Jaśkiewicz, M.; Janczura, A.; Nowicka, J.; Kamysz, W. Methods Used for the eradication of staphylococcal biofilms. Antibiotics 2019, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Sternberg, C.; Bjarnsholt, T.; Shirtliff, M. Methods for Dynamic Investigations of Surface-Attached In Vitro Bacterial and Fungal Biofilms. In Microbial Biofilms. Methods in Molecular Biology (Methods and Protocols); Donelli, G., Ed.; Humana Press: New York, NY, USA, 2014; Volume 1147. [Google Scholar]
- Feng, G.; Cheng, Y.; Wang, S.; Borca-Tasciuc, D.A.; Worobo, R.W.; Moraru, C.I. Bacterial attachment and biofilm formation on surfaces are reduced by small-diameter nanoscale pores: How small is small enough? npj Biofilms Microbiomes 2015, 1, 1–9. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | MIC (µg/mL) |
|---|---|
| S. epidermidis ATCC 35984 | 128 |
| S. epidermidis ATCC 12228 | 128 |
| S. aureus ATCC 29213 | 128 |
| S. aureus ATCC 25923 | 128 |
| MSSA 4471313 | 128 |
| MSSA 4475564 | 128 |
| MSSA 4466686 | 128 |
| MSSA 4467080 | 128 |
| MSSA 4467076 | 128 |
| MSSA 4468505 | 128 |
| MRSA 45300223 | 128 |
| MRSA 9944662 | 128 |
| MRSA 9935169 | 128 |
| No. | Number | Material | Ward | Antibiogram 1 |
|---|---|---|---|---|
| 1 | 4471313 | Nasal swab | Intensive care | Pen.–R, Met.–S, Clin.–S, Ery.-S |
| 2 | 4475564 | Nasal swab | Internal | Pen.–R, Met.–S, Clin.–R, Ery.-R |
| 3 | 4466686 | Sputum | Surgical | Pen.–R, Met.–S, Clin.–R, Ery.–R |
| 4 | 4467080 | Nasal swab | Internal | Pen.–R, Met.–S, Clin.–S, Ery.–S |
| 5 | 4467076 | A swab from the ear | Laryngology | Pen.–R, Met.–S, Clin.–S, Ery.–S |
| 6 | 4468505 | Nasal swab | Internal | Pen.–R, Met.–S, Clin.–R, Ery.–R |
| 7 | 45300223 | Blood | Pediatrics | Pen–R, Met–R, Clin–R, Ery–R |
| 8 | 9935169 | Wound | Dispensary | Pen.–R, Met.–R, Clin.–R, Ery.–R |
| 9 | 9944662 | Nasal swab | Dermatology | Pen.–R, Met.–R, Clin.–R, Ery.–R |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grecka, K.; Xiong, Z.R.; Chen, H.; Pełka, K.; Worobo, R.W.; Szweda, P. Effect of Ethanol Extracts of Propolis (EEPs) against Staphylococcal Biofilm—Microscopic Studies. Pathogens 2020, 9, 646. https://doi.org/10.3390/pathogens9080646
Grecka K, Xiong ZR, Chen H, Pełka K, Worobo RW, Szweda P. Effect of Ethanol Extracts of Propolis (EEPs) against Staphylococcal Biofilm—Microscopic Studies. Pathogens. 2020; 9(8):646. https://doi.org/10.3390/pathogens9080646
Chicago/Turabian StyleGrecka, Katarzyna, Zirui Ray Xiong, Hanyu Chen, Karolina Pełka, Randy W. Worobo, and Piotr Szweda. 2020. "Effect of Ethanol Extracts of Propolis (EEPs) against Staphylococcal Biofilm—Microscopic Studies" Pathogens 9, no. 8: 646. https://doi.org/10.3390/pathogens9080646