Human Bocavirus in Brazil: Molecular Epidemiology, Viral Load and Co-Infections


Abstract
:1. Introduction
2. Materials and Methods
2.1. Stool Collection and Ethics Aspects
2.2. Nucleic Acid Extraction
2.3. HBoV Detection and Quantification
2.4. Molecular Characterization and Genotyping
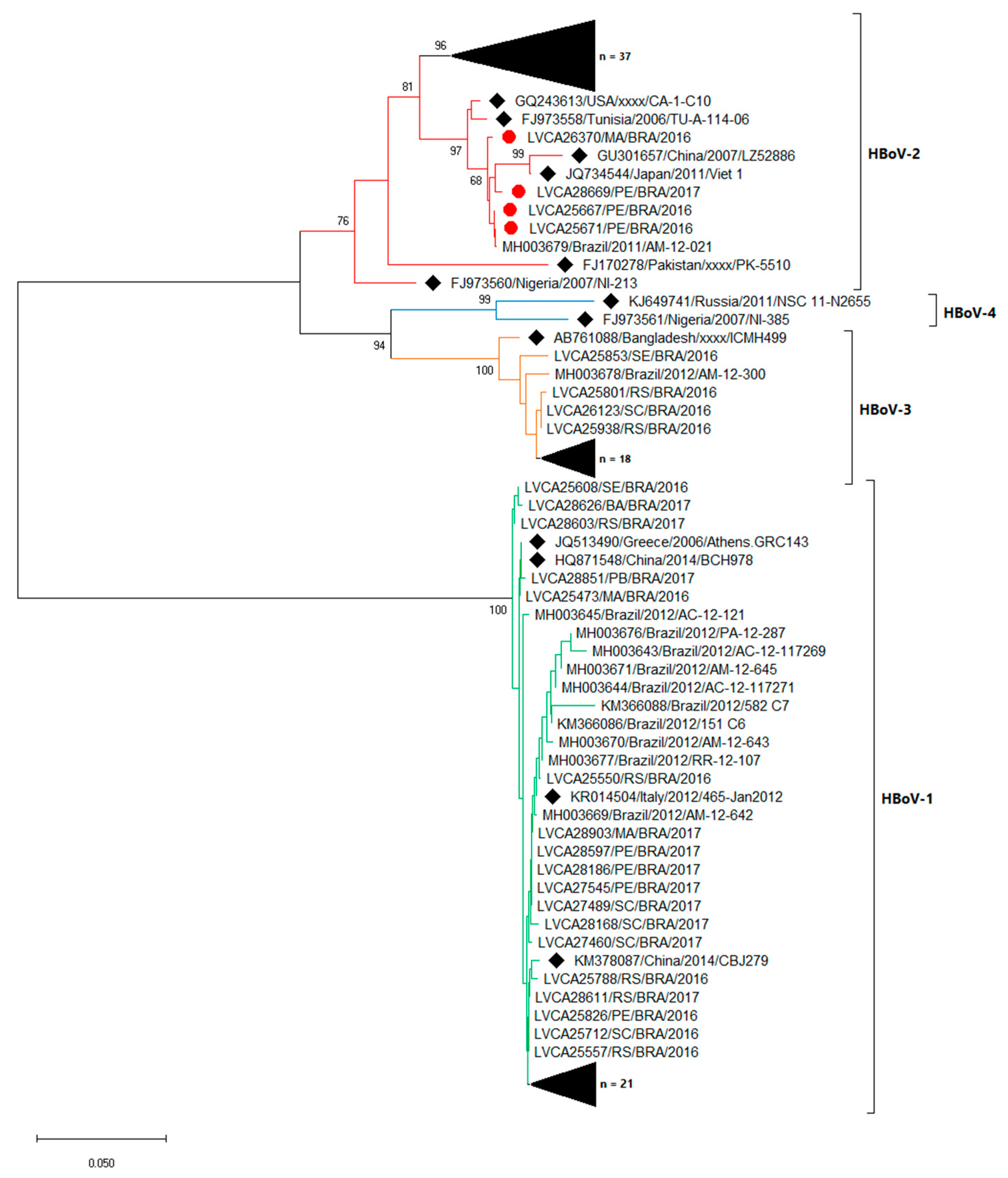
2.5. Phylogenetic Analysis
2.6. Gastroenteric Viruses Detection and Quantification
2.7. Statistical Analysis
3. Results
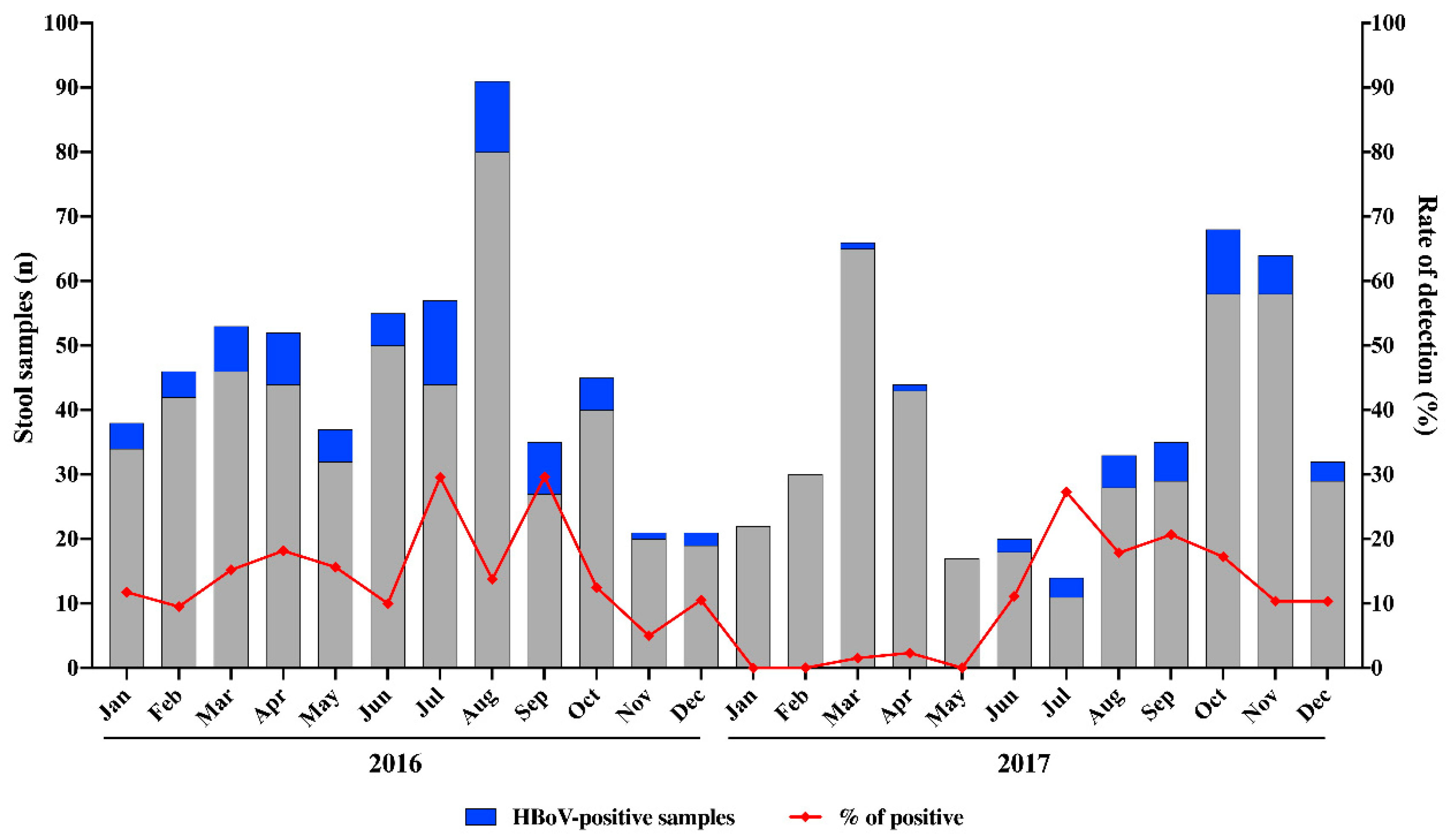
3.1. HBoV Epidemiology
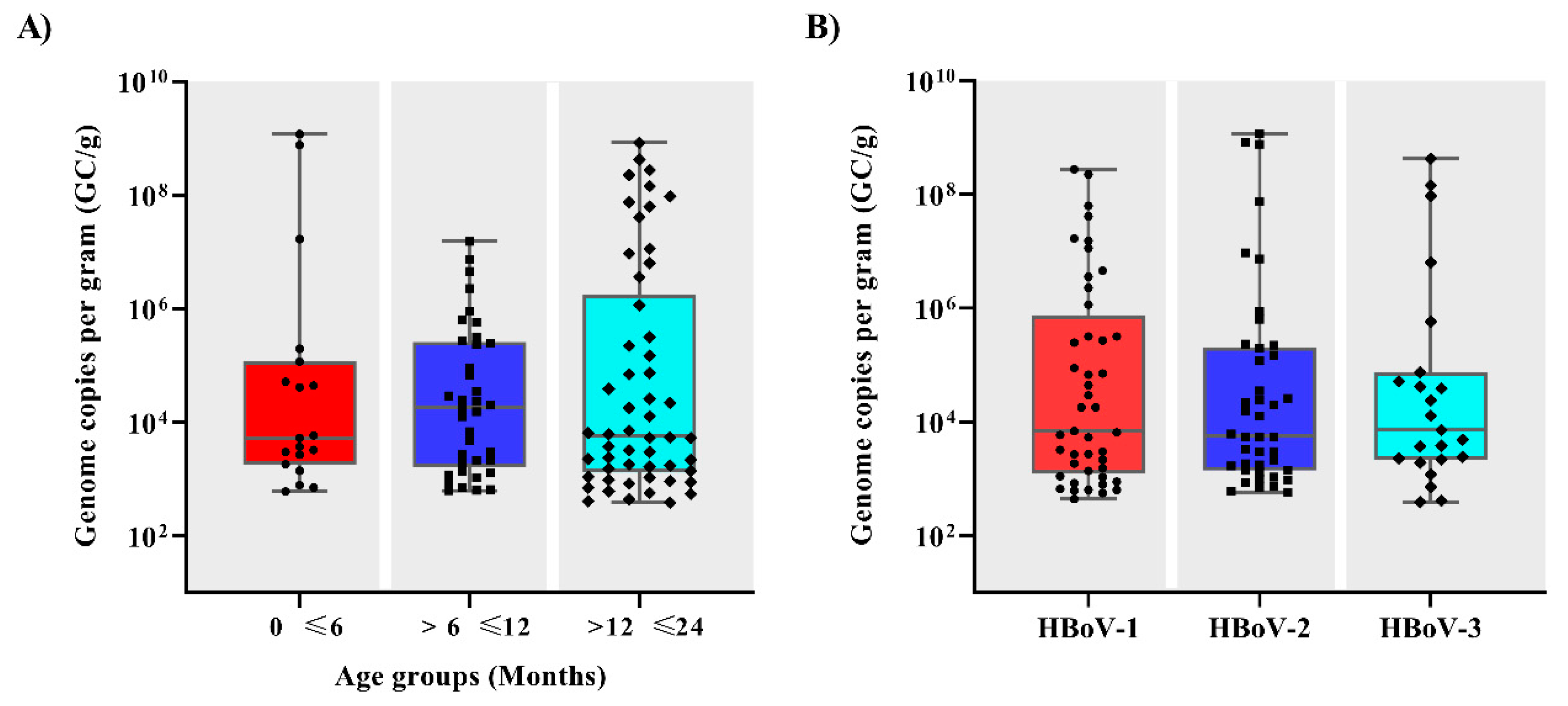
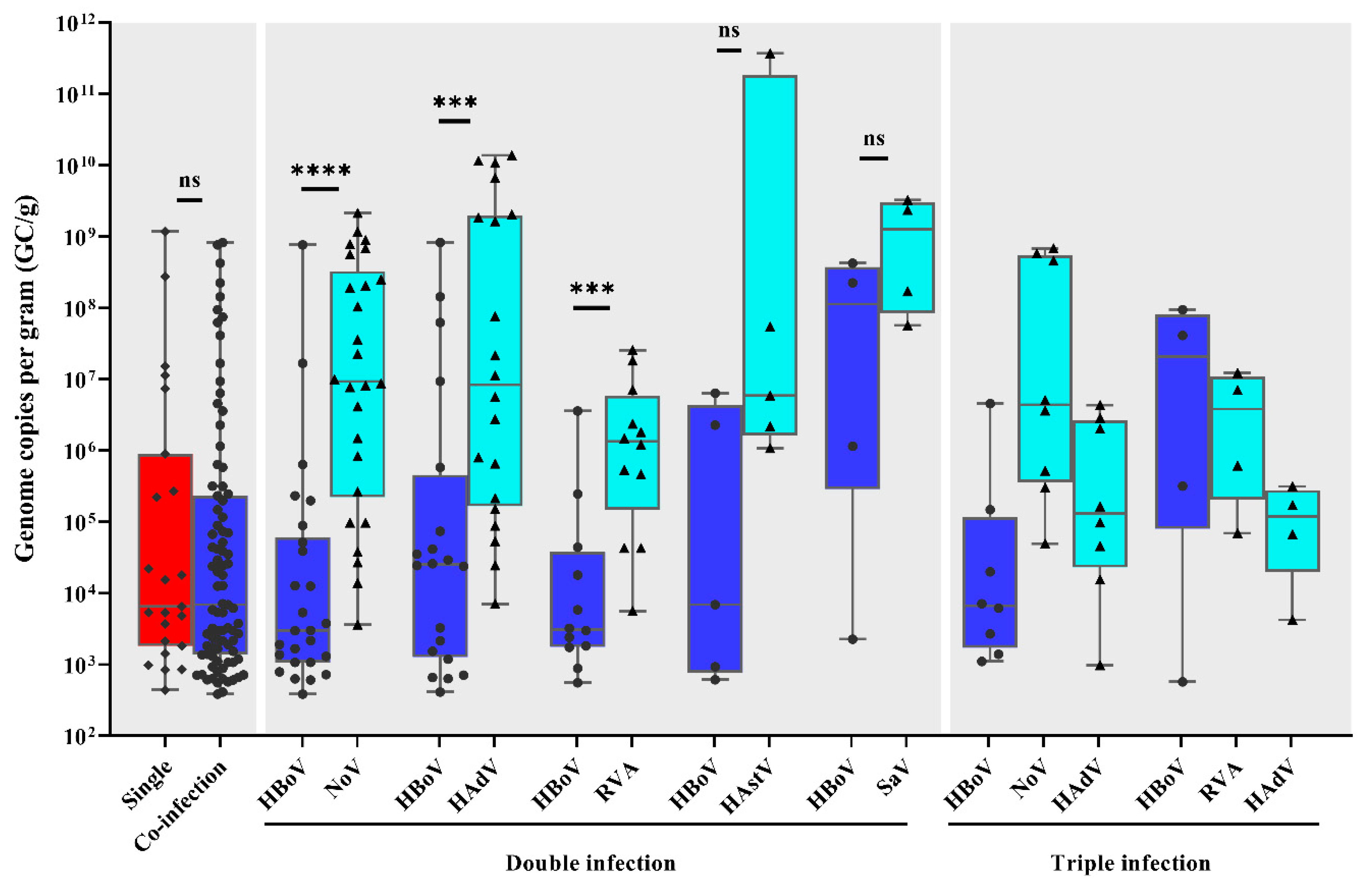
3.2. Viral Load and Co-Infections
3.3. Molecular Characterization
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Walker, C.L.F.; Rudan, I.; Liu, L.; Nair, H.; Theodoratou, E.; Bhutta, Z.A.; O’Brien, K.L.; Campbell, H.; Black, R.E. Global burden of childhood pneumonia and diarrhoea. Lancet 2013, 381, 1405–1416. [Google Scholar] [CrossRef]
- Troeger, C.; Forouzanfar, M.; Rao, P.C.; Khalil, I.; Brown, A.; Reiner, R.C.; Fullman, N.; Thompson, R.L.; Abajobir, A.; Ahmed, M.; et al. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 909–948. [Google Scholar] [CrossRef] [Green Version]
- Wilhelmi, I.; Roman, E.; Sanchez-Fauquier, A. Viruses causing gastroenteritis. Clin. Microbiol. Infect. 2003, 9, 247–262. [Google Scholar] [CrossRef] [Green Version]
- Bányai, K.; Estes, M.K.; Martella, V.; Parashar, U.D. Viral gastroenteritis. Lancet 2018, 392, 175–186. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Nicholson, M.R.; Van Horn, G.T.; Tang, Y.-W.; Vinjé, J.; Payne, D.C.; Edwards, K.M.; Chappell, J.D. Using Multiplex Molecular Testing to Determine the Etiology of Acute Gastroenteritis in Children. J. Pediatr. 2016, 176, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glass, R.I.; Bresee, J.S. Astroviruses, Enteric Adenoviruses, and Other Gastroenteritis Viral Infections. Trop. Infect. Dis. Princ. Pathog. Pract. 2011, 416–419. [Google Scholar] [CrossRef]
- Allander, T.; Tammi, M.T.; Eriksson, M.; Bjerkner, A.; Tiveljung-Lindell, A.; Andersson, B. Cloning of a human parvovirus by molecular screening of respiratory tract samples. Proc. Natl. Acad. Sci. USA 2005, 102, 12891–12896. [Google Scholar] [CrossRef] [Green Version]
- Arthur, J.L.; Higgins, G.D.; Davidson, G.P.; Givney, R.C.; Ratcliff, R.M. A Novel Bocavirus Associated with Acute Gastroenteritis in Australian Children. PLoS Patho. 2009, 5. [Google Scholar] [CrossRef]
- Kapoor, A.; Slikas, E.; Simmonds, P.; Chieochansin, T.; Naeem, A.; Shaukat, S.; Alam, M.M.; Sharif, S.; Angez, M.; Zaidi, S.; et al. A new bocavirus species in human stool. J. Infect. Dis. 2009, 199, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A.; Simmonds, P.; Slikas, B.; Li, L.; Bodhidatta, L.; Sethabutr, O.; Triki, H.; Bahri, O.; Oderinde, B.; Baba, M.; et al. Human bocaviruses are highly diverse, dispersed, recombination prone, and prevalent enteric infections. J. Infect. Dis. 2010, 201, 1633–1643. [Google Scholar] [CrossRef] [PubMed]
- Paloniemi, M.; Lappalainen, S.; Salminen, M.; Kätkä, M.; Kantola, K.; Hedman, L.; Hedman, K.; Söderlund-Venermo, M.; Vesikari, T. Human bocaviruses are commonly found in stools of hospitalized children without causal association to acute gastroenteritis. Eur. J. Pediatr. 2014, 173, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Khurshid, A.; Shaukat, S.; Sharif, S.; Suleman, R.M.; Angez, M.; Nisar, N.; Aamir, U.B.; Naeem, M.; Zaidi, S.S.Z. Human bocavirus in Pakistani children with gastroenteritis. J. Med. Virol. 2015, 87, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Ong, D.S.Y.; Schuurman, R.; Heikens, E. Human bocavirus in stool: A true pathogen or an innocent bystander? J. Med. Virol. 2016, 74, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Rosa, G.L.; Libera, S.D.; Iaconelli, M.; Donia, D.; Cenko, F.; Xhelilaj, G.; Cozza, P.; Divizia, M. Human bocavirus in children with acute gastroenteritis in Albania. J. Med. Virol. 2016, 88, 906–910. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Chen, Y.; Chen, J.; Hu, P.; Zheng, T.; Xu, X.; Pei, X. Prevalence and clinical profile of human bocavirus in children with acute gastroenteritis in Chengdu, West China, 2012–2013. J. Med. Virol. 2017, 89, 1743–1748. [Google Scholar] [CrossRef] [PubMed]
- Schildgen, O. Human Bocavirus: Lessons Learned to Date. Pathogens 2013, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Guido, M.; Tumolo, M.R.; Verri, T.; Romano, A.; Serio, F.; De Giorgi, M.; De Donno, A.; Bagordo, F.; Zizza, A. Human bocavirus: Current knowledge and future challenges. World J. Gastroenterol. 2016, 22, 8684–8697. [Google Scholar] [CrossRef]
- Netshikweta, R.; Chidamba, L.; Nadan, S.; Taylor, M.B.; Page, N.A. Molecular epidemiology of human bocavirus infection in hospitalized children with acute gastroenteritis in South Africa, 2009–2015. J. Med. Virol. 2019, jmv.25634. [Google Scholar] [CrossRef]
- Soares, L.S.; Lima, A.B.F.; Pantoja, K.C.; Lobo, P.S.; Cruz, J.F.; Guerra, S.F.S.; Bezerra, D.A.M.; Bandeira, R.S.; Mascarenhas, J.D.P. Molecular epidemiology of human bocavirus in children with acute gastroenteritis from North Region of Brazil. J. Med. Microbiol. 2019, 68, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Kantola, K.; Sadeghi, M.; Antikainen, J.; Kirveskari, J.; Delwart, E.; Hedman, K.; Söderlund-Venermo, M. Real-Time Quantitative PCR Detection of Four Human Bocaviruses. J. Clin. Microbiol. 2010, 48, 4044–4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Hernroth, B.E.; Conden-Hansson, A.-C.; Rehnstam-Holm, A.-S.; Girones, R.; Allard, A.K. Environmental factors influencing human viral pathogens and their potential indicator organisms in the blue mussel, Mytilus edulis: The first Scandinavian report. Appl. Environ. Microbiol. 2002, 68, 4523–4533. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, T.; Kojima, S.; Shinohara, M.; Uchida, K.; Fukushi, S.; Hoshino, F.B.; Takeda, N.; Katayama, K. Broadly reactive and highly sensitive assay for Norwalk-like viruses based on real-time quantitative reverse transcription-PCR. J. Clin. Microbiol. 2003, 41, 1548–1557. [Google Scholar] [CrossRef] [Green Version]
- Oka, T.; Katayama, K.; Hansman, G.S.; Kageyama, T.; Ogawa, S.; Wu, F.-T.; White, P.A.; Takeda, N. Detection of human sapovirus by real-time reverse transcription-polymerase chain reaction. J. Med. Virol. 2006, 78, 1347–1353. [Google Scholar] [CrossRef]
- Zeng, S.-Q.; Halkosalo, A.; Salminen, M.; Szakal, E.D.; Puustinen, L.; Vesikari, T. One-step quantitative RT-PCR for the detection of rotavirus in acute gastroenteritis. J. Virol. Methods 2008, 153, 238–240. [Google Scholar] [CrossRef]
- Dai, Y.; Xu, Q.; Wu, X.; Hu, G.; Tang, Y.; Li, J.; Chen, Q.; Nie, J. Development of real-time and nested RT-PCR to detect astrovirus and one-year survey of astrovirus in Jiangmen City, China. Arch. Virol. 2010, 155, 977–982. [Google Scholar] [CrossRef]
- Leitão, G.A.A.; Olivares, A.I.O.; Pimenta, Y.C.; Delgado, I.F.; Miagostovich, M.P.; Leite, J.P.G.; Moraes, M.T.B. de Human Bocavirus genotypes 1 and 2 detected in younger Amazonian children with acute gastroenteritis or respiratory infections, respectively. Int. J. Infect. Dis. 2020, 95, 32–37. [Google Scholar] [CrossRef]
- Albuquerque, M.C.M.; Rocha, L.N.; Benati, F.J.; Soares, C.C.; Maranhão, A.G.; Ramírez, M.L.; Erdman, D.; Santos, N. Human Bocavirus Infection in Children with Gastroenteritis, Brazil. Emerg. Infect. Dis. 2007, 13, 1756–1758. [Google Scholar] [CrossRef]
- De Sousa, T.T.; Souza, M.; Fiaccadori, F.S.; Borges, A.M.T.; da Costa, P.S.; das Dôres de Paula Cardoso, D. Human bocavirus 1 and 3 infection in children with acute gastroenteritis in Brazil. Mem. Inst. Oswaldo Cruz 2012, 107, 800–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, G.S.; Sampaio, M.L.S.; Menezes, A.D.L.; Tigre, D.M.; Costa, L.F.M.; Chinalia, F.A.; Sardi, S.I. Human bocavirus in acute gastroenteritis in children in Brazil. J. Med. Virol. 2016, 88, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Portes, S.A.R.; Carvalho-Costa, F.A.; Rocha, M.S.; Fumian, T.M.; Maranhão, A.G.; de Assis, R.M.; da Penha Trindade Pinheiro Xavier, M.; Rocha, M.S.; Miagostovich, M.P.; Leite, J.P.G.; et al. Enteric viruses in HIV-1 seropositive and HIV-1 seronegative children with diarrheal diseases in Brazil. PLoS ONE 2017, 12, e0183196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, L.R.P.; Calvet, F.C.; Sousa, K.L.; Silva, V.P.; Lobo, P.S.; Penha, E.T.; Guerra, S.F.S.; Bezerra, D.A.M.; Mascarenhas, J.D.P.; Pinheiro, H.H.C.; et al. Prevalence of rotavirus and human bocavirus in immunosuppressed individuals after renal transplantation in the Northern Region of Brazil. J. Med. Virol. 2019, 91, 2125–2133. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.C.L.; Dábilla, N.A.S.; Almeida, T.N.; Fiaccadori, F.S.; Souza, T.T.; Cardoso, D.; Arantes, A.; Souza, M. Human bocavirus detection and quantification in fecal and serum specimens from recipients of allogeneic hematopoietic stem cell transplantation: A longitudinal study. J. Med. Virol. 2019. [Google Scholar] [CrossRef]
- Vicente, D.; Cilla, G.; Montes, M.; Pérez-Yarza, E.G.; Pérez-Trallero, E. Human Bocavirus, a Respiratory and Enteric Virus. Emerg. Infect. Dis. 2007, 13, 636–637. [Google Scholar] [CrossRef]
- Cheng, W.; Jin, Y.; Duan, Z.; Xu, Z.; Qi, H.; Zhang, Q.; Yu, J.; Zhu, L.; Jin, M.; Liu, N.; et al. Human Bocavirus in Children Hospitalized for Acute Gastroenteritis: A Case-Control Study. Clin. Infect. Dis. 2008, 47, 161–167. [Google Scholar] [CrossRef]
- Yu, J.-M.; Li, D.-D.; Xu, Z.-Q.; Cheng, W.-X.; Zhang, Q.; Li, H.-Y.; Cui, S.-X.; Miao-Jin; Yang, S.-H.; Fang, Z.-Y. Human bocavirus infection in children hospitalized with acute gastroenteritis in China. J. Clin. Virol. 2008, 42, 280–285. [Google Scholar] [CrossRef]
- Levican, J.; Navas, E.; Orizola, J.; Avendaño, L.F.; Gaggero, A. Human Bocavirus in Children with Acute Gastroenteritis, Chile, 1985–2010. Emerg. Infect. Dis. 2013, 19. [Google Scholar] [CrossRef]
- Rikhotso, M.C.; Khumela, R.; Kabue, J.P.; Traoré-Hoffman, A.N.; Potgieter, N. Predominance of Human Bocavirus Genotype 1 and 3 in Outpatient Children with Diarrhea from Rural Communities in South Africa, 2017–2018. Pathogens 2020, 9, 245. [Google Scholar] [CrossRef] [Green Version]
- Rikhotso, M.C.; Kabue, J.P.; Ledwaba, S.E.; Traoré, A.N.; Potgieter, N. Prevalence of Human Bocavirus in Africa and Other Developing Countries between 2005 and 2016: A Potential Emerging Viral Pathogen for Diarrhea. J. Trop. Med. 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasure, N.; Gopalkrishna, V. Molecular epidemiology and clinical severity of Human Bocavirus (HBoV) 1–4 in children with acute gastroenteritis from Pune, Western India. J. Med. Virol. 2017, 89, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Karalar, L.; Lindner, J.; Schimanski, S.; Kertai, M.; Segerer, H.; Modrow, S. Prevalence and clinical aspects of human bocavirus infection in children. Clin. Microbiol. Infect. 2010, 16, 633–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hustedt, J.W.; Christie, C.; Hustedt, M.M.; Esposito, D.; Vazquez, M. Seroepidemiology of Human Bocavirus Infection in Jamaica. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Turin, C.G.; Ochoa, T.J. The Role of Maternal Breast Milk in Preventing Infantile Diarrhea in the Developing World. Curr. Trop. Med. Rep. 2014, 1, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Smuts, H.; Hardie, D. Human Bocavirus in Hospitalized Children, South Africa. Emerg. Infect. Dis. 2006, 12, 1457–1458. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Yip, C.C.Y.; Que, T.; Lee, R.A.; Au-Yeung, R.K.H.; Zhou, B.; So, L.; Lau, Y.; Chan, K.; Woo, P.C.Y.; et al. Clinical and Molecular Epidemiology of Human Bocavirus in Respiratory and Fecal Samples from Children in Hong Kong. J. Infect. Dis. 2007, 196, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.I.; Chung, J.; Han, T.H.; Song, M.; Hwang, E. Detection of Human Bocavirus in Children Hospitalized because of Acute Gastroenteritis. J. Infect. Dis. 2007, 196, 994–997. [Google Scholar] [CrossRef] [Green Version]
- Chhabra, P.; Payne, D.C.; Szilagyi, P.G.; Edwards, K.M.; Staat, M.A.; Shirley, S.H.; Wikswo, M.; Nix, W.A.; Lu, X.; Parashar, U.D.; et al. Etiology of Viral Gastroenteritis in Children <5 Years of Age in the United States, 2008–2009. J. Infect. Dis. 2013, 208, 790–800. [Google Scholar] [CrossRef] [Green Version]
- Proença-Modena, J.L.; Gagliardi, T.B.; Escremim de Paula, F.; Iwamoto, M.A.; Criado, M.F.; Camara, A.A.; Acrani, G.O.; Cintra, O.A.L.; Cervi, M.C.; de Paula Arruda, L.K.; et al. Detection of Human Bocavirus mRNA in Respiratory Secretions Correlates with High Viral Load and Concurrent Diarrhea. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Cheng, W.; Li, B.; Li, J.; Lan, B.; Duan, Z. Development of a Real-Time PCR Assay for Detecting and Quantifying Human Bocavirus 2. J. Clin. Microbiol. 2011, 49, 1537–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Cheng, W.; Xu, Z.; Liu, N.; Yu, J.; Li, H.; Jin, M.; Li, D.; Zhang, Q.; Duan, Z. High prevalence of human bocavirus 2 and its role in childhood acute gastroenteritis in China. J. Clin. Virol. 2011, 52, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, S.; Allen, D.J.; Aladin, F.; Gallimore, C.; Iturriza-Gómara, M. Human Bocaviruses Are Not Significantly Associated with Gastroenteritis: Results of Retesting Archive DNA from a Case Control Study in the UK. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, A.; Kesti, O.; Elenius, V.; Eskola, A.L.; Døllner, H.; Altunbulakli, C.; Akdis, C.A.; Söderlund-Venermo, M.; Jartti, T. Human bocaviruses and paediatric infections. Lancet Child. Adolesc. Health 2019, 3, 418–426. [Google Scholar] [CrossRef]
- Neske, F.; Blessing, K.; Tollmann, F.; Schubert, J.; Rethwilm, A.; Kreth, H.W.; Weissbrich, B. Real-Time PCR for Diagnosis of Human Bocavirus Infections and Phylogenetic Analysis. J. Clin. Microbiol. 2007, 45, 2116–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Zhang, X.; Xu, H. Don’t Overlook Digestive Symptoms in Patients with 2019 Novel Coronavirus Disease (COVID-19). Clin. Gastroenterol. Hepatol. 2020, 18, 1636–1637. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, Y.; Gao, R.; Lu, R.; Han, K.; Wu, G.; Tan, W. Detection of SARS-CoV-2 in Different Types of Clinical Specimens. JAMA 2020. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Fan, J.; Yu, F.; Feng, B.; Lou, B.; Zou, Q.; Xie, G.; Lin, S.; Wang, R.; Yang, X.; et al. Viral load dynamics and disease severity in patients infected with SARS-CoV-2 in Zhejiang province, China, January-March 2020: Retrospective cohort study. BMJ 2020, m1443. [Google Scholar] [CrossRef] [Green Version]
- Lamers, M.M.; Beumer, J.; van der Vaart, J.; Knoops, K.; Puschhof, J.; Breugem, T.I.; Ravelli, R.B.G.; Paul van Schayck, J.; Mykytyn, A.Z.; Duimel, H.Q.; et al. SARS-CoV-2 productively infects human gut enterocytes. Science 2020, 369, 50–54. [Google Scholar] [CrossRef]
- Jartti, T.; Hedman, K.; Jartti, L.; Ruuskanen, O.; Allander, T.; Söderlund-Venermo, M. Human bocavirus—The first 5 years. Rev. Med. Virol. 2012, 22, 46–64. [Google Scholar] [CrossRef]
- Kenmoe, S.; Vernet, M.-A.; Njankouo-Ripa, M.; Penlap, V.B.; Vabret, A.; Njouom, R. Phylogenic analysis of human bocavirus detected in children with acute respiratory infection in Yaounde, Cameroon. BMC Res. Notes 2017, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, N.; Peret, T.C.T.; Humphrey, C.D.; Albuquerque, M.C.M.; Silva, R.C.; Benati, F.J.; Lu, X.; Erdman, D.D. Human bocavirus species 2 and 3 in Brazil. J. Clin. Virol. 2010, 48, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Han, T.-H.; Kim, C.-H.; Park, S.-H.; Kim, E.-J.; Chung, J.-Y.; Hwang, E.-S. Detection of Human Bocavirus-2 in children with acute Gastroenteritis in South Korea. Arch. Virol. 2009, 154, 1923–1927. [Google Scholar] [CrossRef] [PubMed]
- Chow, B.D.W.; Ou, Z.; Esper, F.P. Newly recognized bocaviruses (HBoV, HBoV2) in children and adults with gastrointestinal illness in the United States. J. Clin. Virol. 2010, 47, 143–147. [Google Scholar] [CrossRef]
- Romani, S.; Mohebbi, S.R.; Khanyaghma, M.; Azimzadeh, P.; Bozorgi, S.M.; Damavand, B.; Jadali, F. Detection of human Bocavirus 1, 2 and 3 from patients with acute gastroenteritis. Gastroenterol. Hepatol. Bed Bench 2013, 6, S77–S81. [Google Scholar] [PubMed]
- De, R.; Liu, L.; Qian, Y.; Zhu, R.; Deng, J.; Wang, F.; Sun, Y.; Dong, H.; Jia, L.; Zhao, L. Risk of acute gastroenteritis associated with human bocavirus infection in children: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0184833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khamrin, P.; Malasao, R.; Chaimongkol, N.; Ukarapol, N.; Kongsricharoern, T.; Okitsu, S.; Hayakawa, S.; Ushijima, H.; Maneekarn, N. Circulating of human bocavirus 1, 2, 3, and 4 in pediatric patients with acute gastroenteritis in Thailand. Infect. Genet. Evol. 2012, 12, 565–569. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HBoV Detection | Positive/Tested (%) | ||
|---|---|---|---|
| Single-Infection | 23/886 (20.9) | ||
| Co-Infection | 87/886 (79.1) | ||
| Co-infections | HBoV + norovirus | 26/110 (23.7) | |
| HBoV + HAdV | 20/110 (18.2) | ||
| HBoV + RVA | 12/110 (11) | ||
| HBoV + astrovirus | 5/110 (4.5) | ||
| HBoV + sapovirus | 4/110 (3.6) | ||
| HBoV + norovirus + HAdV | 8/110 (7.3) | ||
| HBoV + RVA + HAdV | 4/110 (3.6) | ||
| HBoV + RVA + norovirus | 3/110 (2.7) | ||
| HBoV + norovirus+ sapovirus | 1/110 (0.9) | ||
| HBoV + RVA + norovirus+ HAdV | 3/110 (2.7) | ||
| HBoV + norovirus+ HAdV + astrovirus | 1/110 (0.9) | ||
| Age groups (months) | |||
| 0 to ≤6 | 19/212 (9) | ||
| >6 to ≤12 | 37/331 (11.2) | ||
| >12 to ≤24 | 54/343 (15.7) * | ||
| Gender | |||
| Male | 69/488 (14.1) | ||
| Female | 41/391 (10.5) | ||
| Genotypes | |||
| HBoV-1 | 39/102 (38) | ||
| HBoV-2 | 41/102 (40) | ||
| HBoV-3 | 22/102 (22) | ||
| Regions | 2016 | 2017 | |
| Northeast | 27/154 (17.5) | 23/180 (12.7) | 50/334 (14.9) |
| Southeast | 9/70 (12.8) | 4/31 (12.9) | 13/101 (12.8) |
| South | 37/254 (14.5) | 10/197 (5.1) * | 47/451 (10.4) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malta, F.C.; Varella, R.B.; Guimarães, M.A.A.M.; Miagostovich, M.P.; Fumian, T.M. Human Bocavirus in Brazil: Molecular Epidemiology, Viral Load and Co-Infections. Pathogens 2020, 9, 645. https://doi.org/10.3390/pathogens9080645
Malta FC, Varella RB, Guimarães MAAM, Miagostovich MP, Fumian TM. Human Bocavirus in Brazil: Molecular Epidemiology, Viral Load and Co-Infections. Pathogens. 2020; 9(8):645. https://doi.org/10.3390/pathogens9080645
Chicago/Turabian StyleMalta, Fábio Correia, Rafael Brandão Varella, Maria Angelica Arpon Marandino Guimarães, Marize Pereira Miagostovich, and Tulio Machado Fumian. 2020. "Human Bocavirus in Brazil: Molecular Epidemiology, Viral Load and Co-Infections" Pathogens 9, no. 8: 645. https://doi.org/10.3390/pathogens9080645