Molecular Characterization and Antimicrobial Susceptibility of C. jejuni Isolates from Italian Wild Bird Populations

 , ,
, ,
Abstract
:1. Introduction
2. Results
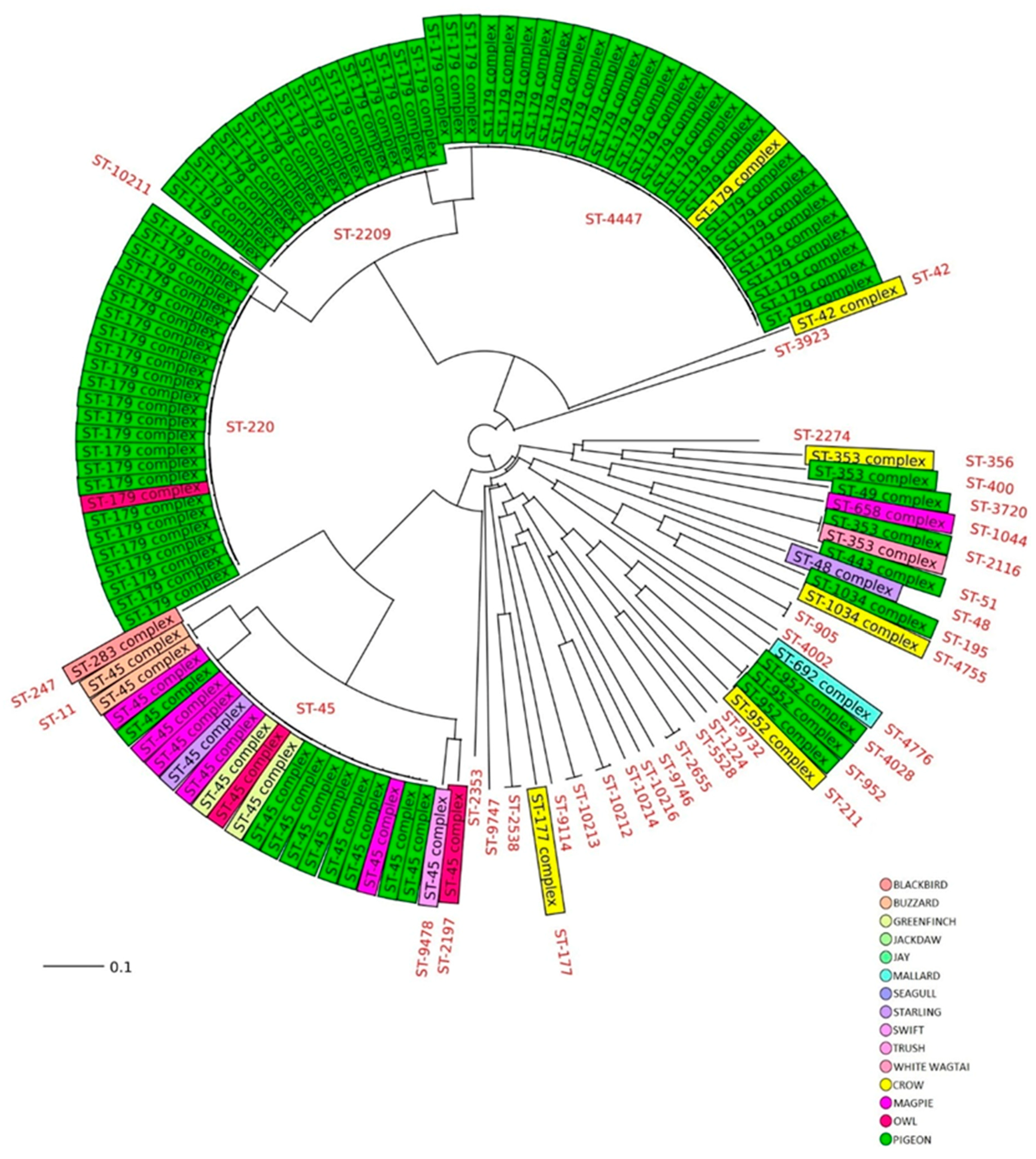
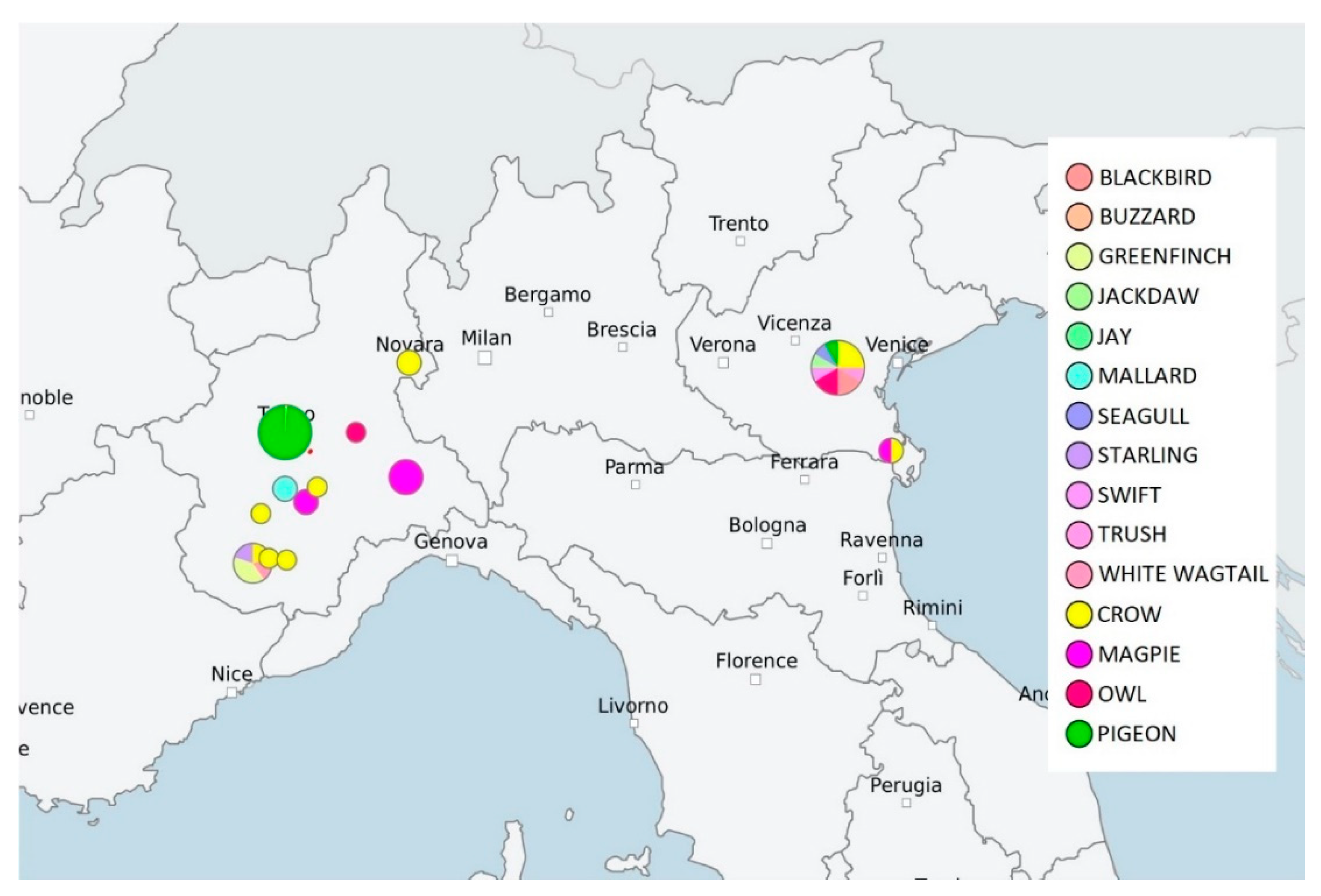
2.1. MLST and cgMLST
2.2. Antimicrobial Resistance Phenotypes
2.3. Detection of Resistance Genes, Mutations and Levels of Concordance Among the Two Type of Resistances
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Species Identification
4.2. Sequence Analysis and Identification of Antibiotic Resistance Genes
4.3. Antimicrobial Susceptibility
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Roberts, J.A.; Cumberland, P.; Sockett, P.N.; Wheeler, J.; Rodrigues, L.C.; Sethi, D.; Roderick, P.J.; on behalf of the IID Study Executive. The study of infectious intestinal disease in England: Socio-economic impact. Epidemiol. Infect. 2003, 130, 1–11. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and foodborne outbreaks in 2015. EFSA J. 2016, 14, 4634. [Google Scholar] [CrossRef]
- Doorduyn, Y.; Van Pelt, W.; Siezen, C.L.E.; Van Der Horst, F.; Van Duynhoven, Y.T.H.P.; Hoebee, B.; Janssen, R. Novel insight in the association between salmonellosis or campylobacteriosis and chronic illness, and the role of host genetics in susceptibility to these diseases. Epidemiol. Infect. 2007, 136, 1225–1234. [Google Scholar] [CrossRef]
- Haagsma, J.A.; Siersema, P.D.; De Wit, N.J.; Havelaar, A.H. Disease burden of post-infectious irritable bowel syndrome in The Netherlands. Epidemiol. Infect. 2010, 138, 1650–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, J.M.; Frost, J.A.; Bolton, F.J.; Wareing, D. Campylobacter Contamination of Raw Meat and Poultry at Retail Sale: Identification of Multiple Types and Comparison with Isolates from Human Infection. J. Food Prot. 2000, 63, 1654–1659. [Google Scholar] [CrossRef] [PubMed]
- Sahin, O.; Fitzgerald, C.; Stroika, S.; Zhao, S.; Sippy, R.; Kwan, P.; Plummer, P.J.; Han, J.; Yaeger, M.J.; Zhang, Q. Molecular Evidence for Zoonotic Transmission of an Emergent, Highly Pathogenic Campylobacter jejuni Clone in the United States. J. Clin. Microbiol. 2011, 50, 680–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skarp, C.; Hänninen, M.-L.; Rautelin, H. Campylobacteriosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, L.A.; Bennett, M.J.; Coffey, P.; Elliott, J.; Jones, T.R.; Jones, R.C.; Lahuerta-Marin, A.; Leatherbarrow, A.H.; McNiffe, K.; Norman, D.; et al. Molecular Epidemiology and Characterization of Campylobacter spp. Isolated from Wild Bird Populations in Northern England. Appl. Environ. Microbiol. 2009, 75, 3007–3015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, N.A.; Stevens, C.P.; Pearl, D.L.; Taboada, E.N.; Jardine, C.M. Generalizability and comparability of prevalence estimates in the wild bird literature: Methodological and epidemiological considerations. Anim. Health Res. Rev. 2020, 18, 1–7. [Google Scholar] [CrossRef]
- Hald, B.; Skov, M.N.; Nielsen, E.M.; Rahbek, C.; Madsen, J.J.; Wainø, M.; Chriél, M.; Nordentoft, S.; Baggesen, D.L.; Madsen, M. Campylobacter jejuni and Campylobacter coli in wild birds on Danish livestock farms. Acta Veter- Scand. 2016, 58, 11. [Google Scholar] [CrossRef] [Green Version]
- Agunos, A.; Waddell, L.A.; Léger, D.; Taboada, E.N. A Systematic Review Characterizing On-Farm Sources of Campylobacter spp. for Broiler Chickens. PLoS ONE 2014, 9, 104905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, C.R.; Neimann, H.; Wegener, H.C.; Tauxe, R.V.I. Epidemiology of Campylobacter jejuni in the United States and other industrialized nations. In Campylobacter; Nachamkin, I., Blaser, M.J., Eds.; ASM Press: Washington, DC, USA, 2000; pp. 121–138. [Google Scholar]
- Mohan, V.; Stevenson, M.A.; Marshall, J.; Fearnhead, P.; Holland, B.; Hotter, G.; French, N.P. Campylobacter jejunicolonization and population structure in urban populations of ducks and starlings in New Zealand. Microbiol. 2013, 2, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Waldenström, J.; Griekspoor, P. Ecology and host associations of Campylobacter in wild birds. In Campylobacter Ecology and Evolution; Sheppard, S., Ed.; Caister Academic Press: Norfolk, UK, 2014; pp. 265–284. [Google Scholar]
- Llarena, A.-K.; Haan, C.P.A.S.-D.; Rossi, M.; Hänninen, M.-L. Characterization of the Campylobacter jejuni Population in the Barnacle Geese Reservoir. Zoonoses Public Health 2014, 62, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Griekspoor, P.; Hansbro, P.M.; Waldenström, J.; Olsen, B. Campylobacter jejuni sequence types show remarkable spatial and temporal stability in Blackbirds. Infect. Ecol. Epidemiol. 2015, 5, 28383. [Google Scholar] [CrossRef] [PubMed]
- Griekspoor, P.; Colles, F.M.; McCarthy, N.; Hansbro, P.M.; Ashhurst-Smith, C.; Olsen, B.; Hasselquist, D.; Maiden, M.C.J.; Waldenström, J. Marked host specificity and lack of phylogeographic population structure of Campylobacter jejuni in wild birds. Mol. Ecol. 2013, 22, 1463–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colles, F.M.; McCarthy, N.; Howe, J.C.; Devereux, C.L.; Gosler, A.G.; Maiden, M.C.J. Dynamics of Campylobacter colonization of a natural host, Sturnus vulgaris (European Starling). Environ. Microbiol. 2009, 11, 258–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friis, C.; Wassenaar, T.M.; Javed, M.A.; Snipen, L.; Lagesen, K.; Hallin, P.F.; Newell, D.G.; Toszeghy, M.; Ridley, A.; Manning, G.; et al. Genomic Characterization of Campylobacter jejuni Strain M1. PLoS ONE 2010, 5, e12253. [Google Scholar] [CrossRef] [Green Version]
- Bang, D.D.; Scheutz, F.; Pedersen, K.; Handberg, K.; Madsen, M. PCR detection of seven virulence and toxin genes of Campylobacter jejuni and Campylobacter coli isolates from Danish pigs and cattle and cytolethal distending toxin production of the isolates. J. Appl. Microbiol. 2003, 94, 1003–1014. [Google Scholar] [CrossRef]
- Weis, A.; Miller, W.A.; Byrne, B.A.; Chouicha, N.; Boyce, W.M.; Townsend, A. Prevalence and Pathogenic Potential of Campylobacter Isolates from Free-Living, Human-Commensal American Crows. Appl. Environ. Microbiol. 2013, 80, 1639–1644. [Google Scholar] [CrossRef] [Green Version]
- Parkhill, J.; Wren, B.W.; Mungall, K.; Ketley, J.M.; Churcher, C.; Basham, D.; Chillingworth, T.; Davies, R.M.; Feltwell, T.; Holroyd, S.; et al. The genome sequence of the food-borne pathogen Campylobacter jejuni reveals hypervariable sequences. Nature 2000, 403, 665–668. [Google Scholar] [CrossRef] [Green Version]
- Cody, A.; McCarthy, N.; Van Rensburg, M.J.; Isinkaye, T.; Bentley, S.D.; Parkhill, J.; Dingle, K.; Bowler, I.C.J.W.; Jolley, K.; Maiden, M.C.J. Real-time genomic epidemiological evaluation of human Campylobacter isolates by use of whole-genome multilocus sequence typing. J. Clin. Microbiol. 2013, 51, 2526–2534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawiec, M.; Woźniak-Biel, A.; Bednarski, M.; Wieliczko, A. Antimicrobial Susceptibility and Genotypic Characteristic of Campylobacter spp. Isolates from Free-Living Birds in Poland. Vector-Borne Zoonotic Dis. 2017, 17, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Cody, A.; McCarthy, N.; Bray, J.; Wimalarathna, H.; Colles, F.M.; Van Rensburg, M.J.J.; Dingle, K.; Waldenström, J.; Maiden, M.C.J. Wild bird-associated C ampylobacter jejuni isolates are a consistent source of human disease, in Oxfordshire, United Kingdom. Environ. Microbiol. Rep. 2015, 7, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Di Giannatale, E.; Garofolo, G.; Alessiani, A.; Di Donato, G.; Candeloro, L.; Vencia, W.; Decastelli, L.; Marotta, F. Tracing Back Clinical Campylobacter jejuni in the Northwest of Italy and Assessing Their Potential Source. Front. Microbiol. 2016, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Llarena, A.-K.; Zhang, J.; Vehkala, M.; Valimaki, N.; Hakkinen, M.; Hänninen, M.-L.; Roasto, M.; Mäesaar, M.; Taboada, E.N.; Barker, D.; et al. Monomorphic genotypes within a generalist lineage of Campylobacter jejuni show signs of global dispersion. Microb. Genom. 2016, 2, e000088. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, S.K.; Dallas, J.F.; MacRae, M.; McCarthy, N.D.; Sproston, E.L.; Gormley, F.J.; Strachan, N.J.; Ogden, I.D.; Maiden, M.C.; Forbes, K.J. Campylobacter genotypes from food animals, environmental sources and clinical disease in Scotland 2005/2006. Int. J. Food Microbiol. 2009, 134, 96–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheppard, S.K.; Dallas, J.F.; Strachan, N.J.C.; Macrae, M.; McCarthy, N.; Wilson, D.J.; Gormley, F.J.; Falush, D.; Ogden, I.D.; Maiden, M.C.J.; et al. Campylobacter Genotyping to Determine the Source of Human Infection. Clin. Infect. Dis. 2009, 48, 1072–1078. [Google Scholar] [CrossRef] [Green Version]
- Cody, A.; Bray, J.; Jolley, K.; McCarthy, N.; Maiden, M.C.J. Core Genome Multilocus Sequence Typing Scheme for Stable, Comparative Analyses of Campylobacter jejuni and C. coli Human Disease Isolates. J. Clin. Microbiol. 2017, 55, 2086–2097. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Halkilahti, J.; Hänninen, M.-L.; Rossi, M. Refinement of Whole-Genome Multilocus Sequence Typing Analysis by Addressing Gene Paralogy. J. Clin. Microbiol. 2015, 53, 1765–1767. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ma, Z.B.; Zeng, Z.L.; Yang, X.W.; Huang, Y.; Liu, J.H. The role of wildlife (wild birds) in the global transmission of antimicrobial resistance genes. Zool. Res. 2017, 38, 55–80. [Google Scholar] [CrossRef] [Green Version]
- de Haan, C.P.A.; Kivisto, R.; Hakkinen, M.; Rautelin, H.; Hanninen, M.L. Decreasing Trend of Overlapping Multilocus Sequence Types between Human and Chicken Campylobacter jejuni Isolates over a Decade in Finland. Appl. Environ. Microbiol. 2010, 76, 5228–5236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovanen, S.M.; Kivistö, R.; Rossi, M.; Schott, T.; Kärkkäinen, U.-M.; Tuuminen, T.; Uksila, J.; Rautelin, H.; Hänninen, M.-L. Multilocus Sequence Typing (MLST) and Whole-Genome MLST of Campylobacter jejuni Isolates from Human Infections in Three Districts during a Seasonal Peak in Finland. J. Clin. Microbiol. 2014, 52, 4147–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weis, A.M.; Huang, B.C.; Storey, D.B.; Kong, N.; Chen, P.; Arabyan, N.; Gilpin, B.; Mason, C.; Townsend, A.K.; Smith, W.A.; et al. Large-Scale Release of Campylobacter Draft Genomes: Resources for Food Safety and Public Health from the 100K Pathogen Genome Project. Genome Announc. 2017, 5, e00925-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévesque, S.; Frost, E.; Arbeit, R.D.; Michaud, S. Multilocus Sequence Typing of Campylobacter jejuni Isolates from Humans, Chickens, Raw Milk, and Environmental Water in Quebec, Canada. J. Clin. Microbiol. 2008, 46, 3404–3411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liakopoulos, A.; Mevius, D.J.; Olsen, B.; Bonnedahl, J. The colistin resistance mcr-1 gene is going wild: Table 1. J. Antimicrob. Chemother. 2016, 71, 2335–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, N.A.; Pearl, D.L.; Taboada, E.N.; Reid-Smith, R.J.; Mulvey, M.R.; Janecko, N.; Mutschall, S.K.; Jardine, C.M. A repeated cross-sectional study of the epidemiology of Campylobacter and antimicrobial resistant Enterobacteriaceae in free-living Canada geese in Guelph, Ontario, Canada. Zoonoses Public Health 2018, 66, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjölund, M.; Bonnedahl, J.; Hernandez, J.; Bengtsson, S.; Cederbrant, G.; Pinhassi, J.; Kahlmeter, G.; Olsen, B. Dissemination of Multidrug-Resistant Bacteria into the Arctic. Emerg. Infect. Dis. 2008, 14, 70–72. [Google Scholar] [CrossRef] [PubMed]
- Webb, A.L.; Selinger, L.B.; Taboada, E.N.; Inglis, G.D. Subtype-Specific Selection for Resistance to Fluoroquinolones but Not to Tetracyclines Is Evident in Campylobacter jejuni Isolates from Beef Cattle in Confined Feeding Operations in Southern Alberta, Canada. Appl. Environ. Microbiol. 2018, 84, e02713-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksomaitiene, J.; Ramonaite, S.; Tamuleviciene, E.; Novoslavskij, A.; Alter, T.; Malakauskas, M. Overlap of Antibiotic Resistant Campylobacter jejuni MLST Genotypes Isolated from Humans, Broiler Products, Dairy Cattle and Wild Birds in Lithuania. Front. Microbiol. 2019, 10, 1377. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Torrens, Y.; Miró, E.; Guirado, P.; Llovet, T.; Muñoz, C.; Cerdà-Cuéllar, M.; Madrid, C.; Balsalobre, C.; Madrid, C. Population Structure, Antimicrobial Resistance, and Virulence-Associated Genes in Campylobacter jejuni Isolated From Three Ecological Niches: Gastroenteritis Patients, Broilers, and Wild Birds. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.E.; Giske, C.; Harbarth, S.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marotta, F.; Garofolo, G.; Di Marcantonio, L.; Di Serafino, G.; Neri, D.; Romantini, R.; Sacchini, L.; Alessiani, A.; Di Donato, G.; Nuvoloni, R.; et al. Antimicrobial resistance genotypes and phenotypes of Campylobacter jejuni isolated in Italy from humans, birds from wild and urban habitats, and poultry. PLoS ONE 2019, 14, e0223804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, H.; Shen, Z.; Wang, Y.; Deng, F.; Liu, D.; Naren, G.; Dai, L.; Su, C.-C.; Wang, B.; Wang, S.; et al. Emergence of a Potent Multidrug Efflux Pump Variant That Enhances Campylobacter Resistance to Multiple Antibiotics. mBio 2016, 7, e01543-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfredson, D.A.; Korolik, V. Isolation and expression of a novel molecular class D beta-lactamase, OXA-61, from Campylobacter jejuni. Antimicrob. Agents Chemother. 2005, 49, 2515–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Tyson, G.; Chen, Y.; Li, C.; Mukherjee, S.; Young, S.; Lam, C.; Folster, J.P.; Whichard, J.M.; McDermott, P.F. Whole-Genome Sequencing Analysis Accurately Predicts Antimicrobial Resistance Phenotypes in Campylobacter spp. Appl. Environ. Microbiol. 2015, 82, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.; Dvorkin, M.; Kulikov, A.; Lesin, V.M.; Nikolenko, S.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Boil. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carraro, L.; Marotta, F.; Janowicz, A.; Patavino, C.; Piccirillo, A. Draft Whole-Genome Sequences of 16 Campylobacter jejuni Isolates Obtained from Wild Birds. Microbiol. Resour. Announc. 2019, 8, e00359-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2016, 45, D566–D573. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Page, A.; Cummins, C.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resistance Phenotypes (No. Isolates) | Resistance Genotypes | MDR | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Family | Species | ST (No. Isolates) | CC | gen | stm | cip | Na | ery | tet | aad | blaOXA-61 | blaOXA-184 | gyrA | Tet(O) | cmeA,B,C,R | cipNatet | cipNateterystmCipEGG | ciperygen | Naerystm |
| Accipitridae | Common Buzzard (Buteo buteo) | 11 (2) | 45 | ||||||||||||||||
| Anatidae | Mallard (Anas platyrhynchos) | 4776 (1) | ST-692 complex | ||||||||||||||||
| 4002 (1) | na | ||||||||||||||||||
| Columbidae | Pigeon (Columba livia) | 10211 * (1) | na | ||||||||||||||||
| 10212 * (2) | na | 1 | |||||||||||||||||
| 10214 * (1) | na | ||||||||||||||||||
| 1956 (1) | ST-1034 complex | ||||||||||||||||||
| 2116 (1) | ST-353 complex | ||||||||||||||||||
| 220 (24) | ST-179 complex | 1 | |||||||||||||||||
| 2209 (17) | ST-179 complex | 1 | |||||||||||||||||
| 2274 (1) | na | ||||||||||||||||||
| 2665 (1) | na | ||||||||||||||||||
| 3720 (1) | ST-49 complex | ||||||||||||||||||
| 3923 (1) | na | ||||||||||||||||||
| 400 (1) | ST-353 complex | ||||||||||||||||||
| 4028 (1) | ST-952 complex | ||||||||||||||||||
| 4447 (29) | ST-179 complex | 2 | 2 | 3 | 3 | 3 | 1 | ||||||||||||
| 45 (9) | ST-45 complex | 1 | 1 | ||||||||||||||||
| 51 (1) | ST-443 complex | ||||||||||||||||||
| 5528 (1) | na | ||||||||||||||||||
| 905 (2) | na | ||||||||||||||||||
| 952 (2) | ST-952 complex | 2 | 2 | ||||||||||||||||
| Corvidae | Crow (Corvus sp.) | 10213 * (2) | na | ||||||||||||||||
| 10216 * (1) | na | ||||||||||||||||||
| 177 (1) | ST-177 complex | ||||||||||||||||||
| 2655 (1) | na | ||||||||||||||||||
| 9732 (1) | na | ||||||||||||||||||
| 4755 (1) | ST-1034 complex | ||||||||||||||||||
| 356 (1) | ST-353 complex | ||||||||||||||||||
| 2111 (1) | ST-952 complex | ||||||||||||||||||
| 4447 (1) | ST-179 complex | ||||||||||||||||||
| 9114 (1) | na | ||||||||||||||||||
| 42 (1) | ST-42 complex | ||||||||||||||||||
| Magpie (Pica pica) | 45 (5) | ST-45 complex | 1 | 1 | |||||||||||||||
| 1224 (1) | Na | ||||||||||||||||||
| 1044 (1) | ST-658 complex | ||||||||||||||||||
| Jackdaw (Corvus onedula) | 9746 (1) | na | |||||||||||||||||
| Eurasian jay (Cyanocitta) | 2538 (1) | na | |||||||||||||||||
| Strigidae | Eurasian Scops Owl (Otus scops) | 2197 (1) | ST-45 complex | ||||||||||||||||
| 220 (1) | ST-179 complex | ||||||||||||||||||
| 45 (1) | ST-45 complex | ||||||||||||||||||
| Fringillidae | Greenfinch (Chloris chloris) | 45 (2) | ST-45 complex | 2 | 1 | 1 | |||||||||||||
| Motacillidae | Whitewagtail (Motacilla alba) | 2116 (1) | ST-353 complex | ||||||||||||||||
| Sturnidae | European starling (Sturnus vulgaris) | 45 (1) | ST-45 complex | ||||||||||||||||
| 48 (1) | ST-48 complex | ||||||||||||||||||
| Turdidae | Blackbirds (Turdus merula) | 9747 (1) | na | ||||||||||||||||
| 267 (1) | ST-283 complex | ||||||||||||||||||
| Thrush (Psophocichla litsitsirupa) | 2538 (1) | na | |||||||||||||||||
| Laridae | Seagul (Larus argentatus) | 2353 (1) | na | ||||||||||||||||
| Apodidae | Swift (Apus apus) | 9478 (1) | ST-45 complex | ||||||||||||||||
| Antibiotic Class | Antibiotics | Genes a | % Resistances (Phenotypes) b | % Resistance (Genotypes) c | Concordance rate d |
|---|---|---|---|---|---|
| Aminoglycosides | Gentamicin (gen) | - | 2.5 | ||
| Streptomycin (stm) | aad | 6.7 | 28.6 | 23.3 | |
| Beta-lactams e | - | blaOXA-61, blaOXA-184 | - | 10, 86.7 | - |
| Fluoroquinolones/ Quinolones | Ciprofloxacin (cip)/ Nalidixic acid (Na) | gyrA | 10 | 33.3 | 30.0 |
| Macrolides | Erytromicin (ery) | erm(B), 23S rRNA | 4.2 | - | - |
| Tetracyclines | Tertracycline (tet) | Tet(O) | 12.5 | 53.3 | 23.4 |
| Multidrug CmeABC efflux system and cmeR | cmeA, cmeB, cmeC, cmeR | - | 100, 100, 99.2, 100 | - | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marotta, F.; Janowicz, A.; Di Marcantonio, L.; Ercole, C.; Di Donato, G.; Garofolo, G.; Di Giannatale, E. Molecular Characterization and Antimicrobial Susceptibility of C. jejuni Isolates from Italian Wild Bird Populations. Pathogens 2020, 9, 304. https://doi.org/10.3390/pathogens9040304
Marotta F, Janowicz A, Di Marcantonio L, Ercole C, Di Donato G, Garofolo G, Di Giannatale E. Molecular Characterization and Antimicrobial Susceptibility of C. jejuni Isolates from Italian Wild Bird Populations. Pathogens. 2020; 9(4):304. https://doi.org/10.3390/pathogens9040304
Chicago/Turabian StyleMarotta, Francesca, Anna Janowicz, Lisa Di Marcantonio, Claudia Ercole, Guido Di Donato, Giuliano Garofolo, and Elisabetta Di Giannatale. 2020. "Molecular Characterization and Antimicrobial Susceptibility of C. jejuni Isolates from Italian Wild Bird Populations" Pathogens 9, no. 4: 304. https://doi.org/10.3390/pathogens9040304