Legionella pneumophila and Protozoan Hosts: Implications for the Control of Hospital and Potable Water Systems


Abstract
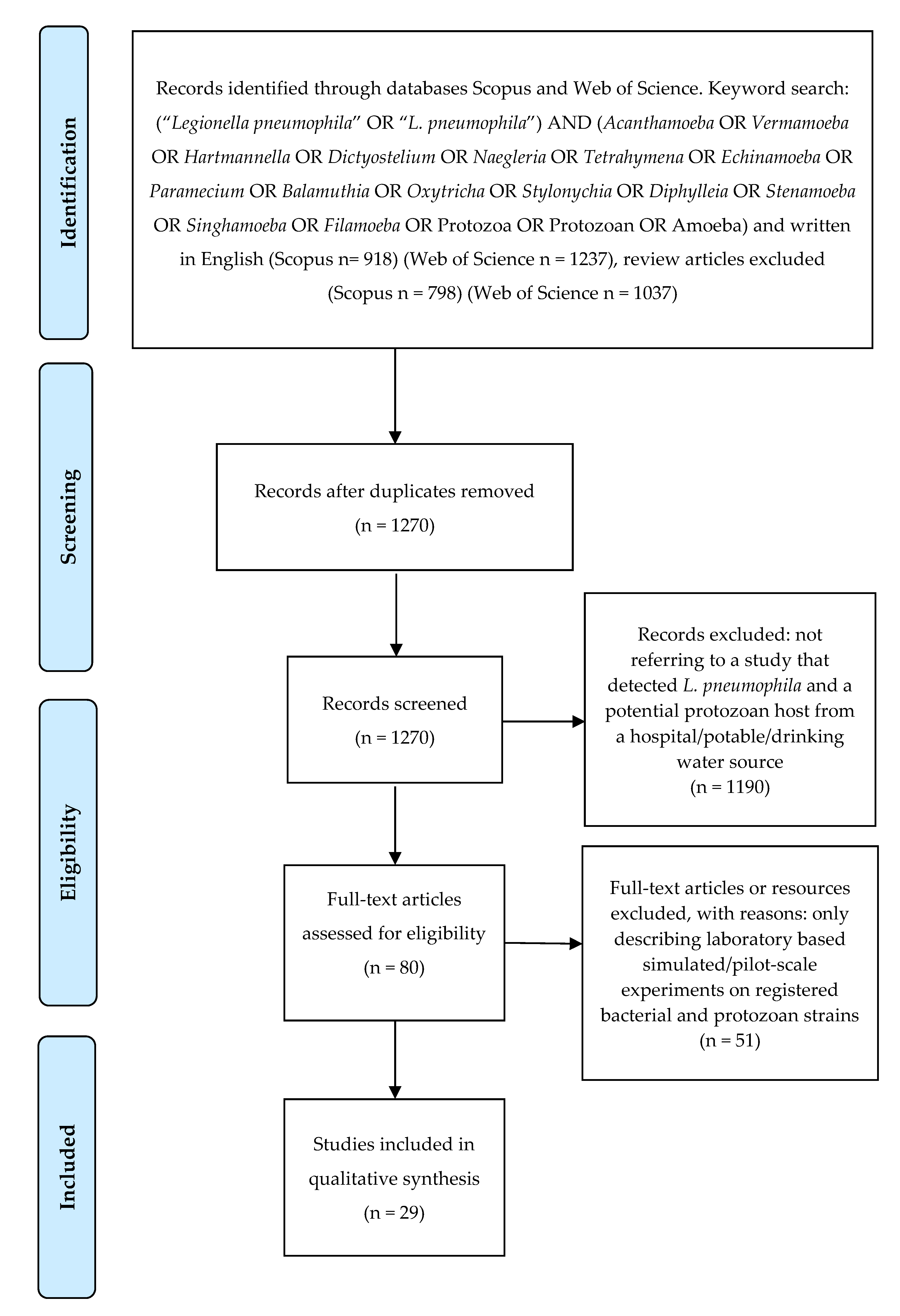
:1. Introduction
2. Results
3. Discussion
3.1. Implications for the Control of L. pneumophila
3.2. Protozoan Host Control Strategies
3.3. Detection Methods
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cunha, B.A.; Burillo, A.; Bouza, E. Legionnaires’ disease. Lancet 2016, 387, 376–385. [Google Scholar] [CrossRef]
- The European Legionnaires’ Disease Surveillance Network; The European Centre for Disease Prevention and Control. Surveillance Atlas of Infectious Diseases. Available online: https://atlas.ecdc.europa.eu/public/index.aspx (accessed on 4 October 2019).
- Smith, P.; Moore, M.; Alexander, N.; Hicks, L.; O’Loughlin, R. Surveillance for travel-associated Legionnaires disease-United States, 2005–2006. Mmwr-Morb Mortal Wkly. Rep. 2007, 56, 1261–1263. [Google Scholar]
- National Notifiable Diseases Surveillance System; Centers for Disease Control and Prevention. National Notifiable Infectious Diseases: Weekly Tables; National Notifiable Diseases Surveillance System; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018.
- Phin, N.; Parry-Ford, F.; Harrison, T.; Stagg, H.R.; Zhang, N.; Kumar, K.; Lortholary, O.; Zumla, A.; Abubakar, I. Epidemiology and clinical management of Legionnaires’ disease. Lancet Infect. Dis. 2014, 14, 1011–1021. [Google Scholar] [CrossRef]
- O’Mahony, M.C.; Stanwell-Smith, R.E.; Tillett, H.E.; Harper, D.; Hutchison, J.G.; Farrell, I.D.; Hutchinson, D.N.; Lee, J.V.; Dennis, P.J.; Duggal, H.V.; et al. The Stafford outbreak of Legionnaires’ disease. Epidemiol. Infect. 1990, 104, 361–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soda, E.A. Vital Signs: Health care–associated Legionnaires’ disease surveillance data from 20 states and a large metropolitan area-United States, 2015. Mmwr-Morb Mortal Wkly. Rep. 2017, 66, 584–589. [Google Scholar] [CrossRef]
- Mercante, J.W.; Winchell, J.M. Current and emerging Legionella diagnostics for laboratory and outbreak investigations. Clin. Microbiol. Rev. 2015, 28, 95–133. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, N.; Higa, F.; Aoki, Y.; Kikuchi, T.; Seki, M.; Tateda, K.; Maki, N.; Uchino, K.; Ogasawara, K.; Kiyota, H.; et al. Distribution of Legionella species and serogroups in patients with culture-confirmed Legionella pneumonia. J. Infect. Chemother 2020. [Google Scholar] [CrossRef]
- Bartram, J.; Chartier, Y.; Lee, J.V.; Pond, K.; Surman-Lee, S. Legionella and the Prevention of Legionellosis; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- NNDSS Annual Report Working Group. Australia’s notifiable disease status, 2015: Annual report of the national notifiable diseases surveillance system. Commun. Dis. Intel. (2018) 2019, 43. [Google Scholar] [CrossRef]
- Graham, F.F.; White, P.S.; Harte, D.J.; Kingham, S.P. Changing epidemiological trends of legionellosis in New Zealand, 1979–2009. Epidemiol. Infect. 2012, 140, 1481–1496. [Google Scholar] [CrossRef] [Green Version]
- Benedict, K.M. Surveillance for waterborne disease outbreaks associated with drinking water—United States, 2013–2014. Mmwr.-Morb. Mortal Wkly. Rep. 2017, 66, 1216. [Google Scholar] [CrossRef]
- Abdel-Nour, M.; Duncan, C.; Low, D.E.; Guyard, C. Biofilms: The stronghold of Legionella pneumophila. Int. J. Mol. Sci 2013, 14, 21660. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Lin, Y.E.; Stout, J.E.; Hwang, C.C.; Vidic, R.D.; Yu, V.L. Effect of flow regimes on the presence of Legionella within the biofilm of a model plumbing system. J. Appl. Microbiol. 2006, 101, 437–442. [Google Scholar] [CrossRef]
- Prussin, A.J., 2nd; Schwake, D.O.; Marr, L.C. Ten questions concerning the aerosolization and transmission of Legionella in the built environment. Build. Environ. 2017, 123, 684–695. [Google Scholar] [CrossRef]
- Ashbolt, N.J. Environmental (Saprozoic) Pathogens of Engineered Water Systems: Understanding Their Ecology for Risk Assessment and Management. Pathogens 2015, 4, 390. [Google Scholar] [CrossRef] [Green Version]
- Falkinham, J.O.; Pruden, A.; Edwards, M. Opportunistic premise plumbing pathogens: Increasingly important pathogens in drinking water. Pathogens 2015, 4, 373. [Google Scholar] [CrossRef] [Green Version]
- Boamah, D.K.; Zhou, G.; Ensminger, A.W.; O’Connor, T.J. From many hosts, one accidental pathogen: The diverse protozoan hosts of Legionella. Front. Cell Infect. Microbiol. 2017, 7, 477. [Google Scholar] [CrossRef] [Green Version]
- Newton, H.J.; Ang, D.K.; van Driel, I.R.; Hartland, E.L. Molecular pathogenesis of infections caused by Legionella pneumophila. Clin. Microbiol. Rev. 2010, 23, 274–298. [Google Scholar] [CrossRef] [Green Version]
- Oliva, G.; Sahr, T.; Buchrieser, C. The Life Cycle of L. pneumophila: Cellular Differentiation Is Linked to Virulence and Metabolism. Front. Cell Infect. Microbiol. 2018, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Bouyer, S.; Imbert, C.; Rodier, M.H.; Hechard, Y. Long-term survival of Legionella pneumophila associated with Acanthamoeba castellanii vesicles. Environ. Microbiol. 2007, 9, 1341–1344. [Google Scholar] [CrossRef]
- Koubar, M.; Rodier, M.H.; Garduno, R.A.; Frere, J. Passage through Tetrahymena tropicalis enhances the resistance to stress and the infectivity of Legionella pneumophila. Fems Microbiol. Lett. 2011, 325, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Jjemba, P.K.; Johnson, W.; Bukhari, Z.; LeChevallier, M.W. Occurrence and control of Legionella in recycled water systems. Pathogens 2015, 4, 470. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.R.; Anderson, J.E.; Mueller, S.A.; Gaines, W.A.; Kendall, A.M. Literature review--efficacy of various disinfectants against Legionella in water systems. Water Res. 2002, 36, 4433–4444. [Google Scholar] [CrossRef]
- Buse, H.Y.; Donohue, M.J.; Ashbolt, N.J. Hartmannella vermiformis inhibition of Legionella pneumophila cultivability. Microb. Ecol. 2013, 66, 715–726. [Google Scholar] [CrossRef]
- Casini, B.; Baggiani, A.; Totaro, M.; Mansi, A.; Costa, A.L.; Aquino, F.; Miccoli, M.; Valentini, P.; Bruschi, F.; Lopalco, P.L.; et al. Detection of viable but non-culturable Legionella in hospital water network following monochloramine disinfection. J. Hosp. Infect. 2018, 98, 46–52. [Google Scholar] [CrossRef]
- Thomas, J.M.; Ashbolt, N.J. Do free-living amoebae in treated drinking water systems present an emerging health risk? Environ. Sci. Technol. 2011, 45, 860–869. [Google Scholar] [CrossRef]
- Yu, V.L.; Plouffe, J.F.; Pastoris, M.C.; Stout, J.E.; Schousboe, M.; Widmer, A.; Summersgill, J.; File, T.; Heath, C.M.; Paterson, D.L.; et al. Distribution of Legionella species and serogroups isolated by culture in patients with sporadic community-acquired legionellosis: An international collaborative survey. J. Infect. Dis. 2002, 186, 127–128. [Google Scholar] [CrossRef] [Green Version]
- Beaute, J.; Zucs, P.; de Jong, B.; European Legionnaires’ Disease Surveillance Network. Legionnaires disease in Europe, 2009–2010. Euro Surveill. 2013, 18, 20417. [Google Scholar] [CrossRef] [Green Version]
- Fields, B.S.; Sanden, G.N.; Barbaree, J.M.; Morrill, W.E.; Wadowsky, R.M.; White, E.H.; Feeley, J.C. Intracellular multiplication of Legionella pneumophila in amoebae isolated from hospital hot water tanks. Curr. Microbiol. 1989, 18, 131–137. [Google Scholar] [CrossRef]
- Nahapetian, K.; Challemel, O.; Beurtin, D.; Dubrou, S.; Gounon, P.; Squinazi, F. The intracellular multiplication of Legionella pneumophila in protozoa from hospital plumbing systems. Res. Microbiol. 1991, 142, 677–685. [Google Scholar] [CrossRef]
- Garcia, A.; Goni, P.; Cieloszyk, J.; Fernandez, M.T.; Calvo-Begueria, L.; Rubio, E.; Fillat, M.F.; Peleato, M.L.; Clavel, A. Identification of free-living amoebae and amoeba-associated bacteria from reservoirs and water treatment plants by molecular techniques. Environ. Sci. Technol. 2013, 47, 3132–3140. [Google Scholar] [CrossRef]
- Moreno, Y.; Moreno-Mesonero, L.; Garcia-Hernandez, J. DVC-FISH to identify potentially pathogenic Legionella inside free-living amoebae from water sources. Environ. Res. 2019, 176, 108521. [Google Scholar] [CrossRef]
- Breiman, R.F.; Fields, B.S.; Sanden, G.N.; Volmer, L.; Meier, A.; Spika, J.S. Association of shower use with Legionnaires’ disease-possible role of ameobas. JAMA-J. Am. Med. Assoc. 1990, 263, 2924–2926. [Google Scholar] [CrossRef]
- Bezanson, G.; Burbridge, S.; Haldane, D.; Yoell, C.; Marrie, T. Diverse populations of Legionella pneumophila present in the water of geographically clustered institutions served by the same water reservoir. J. Clin. Microbiol. 1992, 30, 570–576. [Google Scholar] [CrossRef] [Green Version]
- Patterson, W.J.; Hay, J.; Seal, D.V.; McLuckie, J.C. Colonization of transplant unit water supplies with Legionella and protozoa: Precautions required to reduce the risk of legionellosis. J. Hosp. Infect. 1997, 37, 7–17. [Google Scholar] [CrossRef]
- Steinert, M.; Ockert, G.; Luck, C.; Hacker, J. Regrowth of Legionella pneumophila in a heat-disinfected plumbing system. Zent. Bakteriol. 1998, 288, 331–342. [Google Scholar] [CrossRef]
- Thomas, V.; Herrera-Rimann, K.; Blanc, D.S.; Greub, G. Biodiversity of amoebae and amoeba-resisting bacteria in a hospital water network. Appl. Environ. Microbiol. 2006, 72, 2428–2438. [Google Scholar] [CrossRef] [Green Version]
- Lagana, P.; Caruso, G.; Piccione, D.; Gioffre, M.E.; Pino, R.; Delia, S. Legionella spp., amoebae and not-fermenting Gram negative bacteria in an Italian university hospital water system. Ann. Agric. Environ. Med. 2014, 21, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Trnkova, K.; Kotrbancova, M.; Spalekova, M.; Fulova, M.; Boledovicova, J.; Vesteg, M. MALDI-TOF MS analysis as a useful tool for an identification of Legionella pneumophila, a facultatively pathogenic bacterium interacting with free-living amoebae: A case study from water supply system of hospitals in Bratislava (Slovakia). Exp. Parasitol 2018, 184, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Muchesa, P.; Leifels, M.; Jurzik, L.; Barnard, T.G.; Bartie, C. Detection of amoeba-associated Legionella pneumophila in hospital water networks of Johannesburg. S. Afr J. Infect. D 2018, 33, 72–75. [Google Scholar] [CrossRef]
- Spagnolo, A.M.; Sartini, M.; Cave, D.D.; Casini, B.; Tuvo, B.; Cristina, M.L. Evaluation of microbiological and free-living protozoa contamination in dental unit waterlines. Int. J. Environ. Res. Public Health 2019, 16, 2468. [Google Scholar] [CrossRef] [Green Version]
- Valster, R.M.; Wullings, B.A.; Bakker, G.; Smidt, H.; van der Kooij, D. Free-living protozoa in two unchlorinated drinking water supplies, identified by phylogenic analysis of 18S rRNA gene sequences. Appl. Environ. Microbiol. 2009, 75, 4736–4746. [Google Scholar] [CrossRef] [Green Version]
- Valster, R.M.; Wullings, B.A.; van der Kooij, D. Detection of protozoan hosts for Legionella pneumophila in engineered water systems by using a biofilm batch test. Appl. Environ. Microbiol. 2010, 76, 7144–7153. [Google Scholar] [CrossRef] [Green Version]
- Valster, R.M.; Wullings, B.A.; van den Berg, R.; van der Kooij, D. Relationships between free-living protozoa, cultivable Legionella spp., and water quality characteristics in three drinking water supplies in the Caribbean. Appl. Environ. Microbiol. 2011, 77, 7321–7328. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Edwards, M.; Falkinham, J.O., 3rd; Pruden, A. Molecular survey of the occurrence of Legionella spp., Mycobacterium spp., Pseudomonas aeruginosa, and amoeba hosts in two chloraminated drinking water distribution systems. Appl. Environ. Microbiol. 2012, 78, 6285–6294. [Google Scholar] [CrossRef] [Green Version]
- Ji, W.T.; Hsu, B.M.; Chang, T.Y.; Hsu, T.K.; Kao, P.M.; Huang, K.H.; Tsai, S.F.; Huang, Y.L.; Fan, C.W. Surveillance and evaluation of the infection risk of free-living amoebae and Legionella in different aquatic environments. Sci. Total Environ. 2014, 499, 212–219. [Google Scholar] [CrossRef]
- Lu, J.; Struewing, I.; Yelton, S.; Ashbolt, N. Molecular survey of occurrence and quantity of Legionella spp., Mycobacterium spp., Pseudomonas aeruginosa and amoeba hosts in municipal drinking water storage tank sediments. J. Appl. Microbiol. 2015, 119, 278–288. [Google Scholar] [CrossRef]
- Lu, J.; Struewing, I.; Vereen, E.; Kirby, A.E.; Levy, K.; Moe, C.; Ashbolt, N. Molecular Detection of Legionella spp. and their associations with Mycobacterium spp., Pseudomonas aeruginosa and amoeba hosts in a drinking water distribution system. J. Appl. Microbiol. 2016, 120, 509–521. [Google Scholar] [CrossRef]
- Lienard, J.; Croxatto, A.; Gervaix, A.; Levi, Y.; Loret, J.F.; Posfay-Barbe, K.M.; Greub, G. Prevalence and diversity of Chlamydiales and other amoeba-resisting bacteria in domestic drinking water systems. New Microbes New Infect. 2017, 15, 107–116. [Google Scholar] [CrossRef]
- Qin, K.; Struewing, I.; Domingo, J.S.; Lytle, D.; Lu, J. Opportunistic pathogens and microbial communities and their associations with sediment physical parameters in drinking water storage tank sediments. Pathogens 2017, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Shen, Y.; Hu, C.; Xing, X.; Zhao, D. Sulfadiazine/ciprofloxacin promote opportunistic pathogens occurrence in bulk water of drinking water distribution systems. Environ. Pollut. 2018, 234, 71–78. [Google Scholar] [CrossRef]
- Wang, H.; Hu, C.; Zhang, S.; Liu, L.; Xing, X. Effects of O3/Cl2 disinfection on corrosion and opportunistic pathogens growth in drinking water distribution systems. J. Environ. Sci. (China) 2018, 73, 38–46. [Google Scholar] [CrossRef]
- Van der Kooij, D.; Veenendaal, H.R.; Italiaander, R.; van der Mark, E.J.; Dignum, M. Primary colonizing Betaproteobacteriales play a key role in the growth of Legionella pneumophila in biofilms on surfaces exposed to drinking water treated by slow sand filtration. Appl. Environ. Microbiol. 2018, 84, e01732–e01818. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, S.; Tang, W.; Yang, Y.; Zhao, J.; Xia, S.; Zhang, W.; Wang, H. Influence of secondary water supply systems on microbial community structure and opportunistic pathogen gene markers. Water Res. 2018, 136, 160–168. [Google Scholar] [CrossRef]
- Wang, H.; Xu, J.; Tang, W.; Li, H.; Xia, S.; Zhao, J.; Zhang, W.; Yang, Y. Removal efficacy of opportunistic pathogens and bacterial community dynamics in two drinking water treatment trains. Small 2019, 15, 1804436. [Google Scholar] [CrossRef]
- Dai, D.J.; Rhoads, W.J.; Katner, A.; Strom, L.; Edwards, M.A.; Pruden, A.; Pieper, K.J. Molecular survey of Legionella and Naegleria fowleri in private well water and premise plumbing following the 2016 Louisiana flood. Environ. Sci.-Wat. Res. 2019, 5, 1464–1477. [Google Scholar] [CrossRef]
- Lau, H.Y.; Ashbolt, N.J. The role of biofilms and protozoa in Legionella pathogenesis: Implications for drinking water. J. Appl. Microbiol. 2009, 107, 368–378. [Google Scholar] [CrossRef]
- Lin, Y.S.; Stout, J.E.; Yu, V.L.; Vidic, R.D. Disinfection of water distribution systems for Legionella. Semin. Respir. Infect. 1998, 13, 147–159. [Google Scholar]
- Whiley, H.; Bentham, R.; Brown, M.H. Legionella persistence in manufactured water systems: Pasteurization potentially selecting for thermal tolerance. Front. Microbiol. 2017, 8, 1330. [Google Scholar] [CrossRef]
- Fields, B.S.; Benson, R.F.; Besser, R.E. Legionella and Legionnaires’ disease: 25 years of investigation. Clin. Microbiol. Rev. 2002, 15, 506–526. [Google Scholar] [CrossRef] [Green Version]
- Dobrowsky, P.H.; Khan, S.; Cloete, T.E.; Khan, W. Molecular detection of Acanthamoeba spp., Naegleria fowleri and Vermamoeba (Hartmannella) vermiformis as vectors for Legionella spp. in untreated and solar pasteurized harvested rainwater. Parasit. Vectors 2016, 9, 539. [Google Scholar] [CrossRef] [Green Version]
- Rhoads, W.J.; Ji, P.; Pruden, A.; Edwards, M.A. Water heater temperature set point and water use patterns influence Legionella pneumophila and associated microorganisms at the tap. Microbiome 2015, 3, 67. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Guidelines for Drinking-Water Quality; WHO: Geneva, Switzerland, 2004. [Google Scholar]
- Muraca, P.; Stout, J.E.; Yu, V.L. Comparative assessment of chlorine, heat, ozone, and UV light for killing Legionella pneumophila within a model plumbing system. Appl. Environ. Microbiol. 1987, 53, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Kilvington, S.; Price, J. Survival of Legionella pneumophila within cysts of Acanthamoeba polyphaga following chlorine exposure. J. Appl. Bacteriol. 1990, 68, 519–525. [Google Scholar] [CrossRef]
- Kool, J.L.; Carpenter, J.C.; Fields, B.S. Effect of monochloramine disinfection of municipal drinking water on risk of nosocomial Legionnaires’ disease. Lancet 1999, 353, 272–277. [Google Scholar] [CrossRef]
- Dupuy, M.; Mazoua, S.; Berne, F.; Bodet, C.; Garrec, N.; Herbelin, P.; Menard-Szczebara, F.; Oberti, S.; Rodier, M.H.; Soreau, S.; et al. Efficiency of water disinfectants against Legionella pneumophila and Acanthamoeba. Water Res. 2011, 45, 1087–1094. [Google Scholar] [CrossRef]
- Donlan, R.M.; Forster, T.; Murga, R.; Brown, E.; Lucas, C.; Carpenter, J.; Fields, B. Legionella pneumophila associated with the protozoan Hartmannella vermiformis in a model multi-species biofilm has reduced susceptibility to disinfectants. Biofouling 2005, 21, 1–7. [Google Scholar] [CrossRef]
- Walker, J.T.; Mackerness, C.W.; Mallon, D.; Makin, T.; Williets, T.; Keevil, C.W. Control of Legionella pneumophila in a hospital water system by chlorine dioxide. J. Ind. Microbiol. 1995, 15, 384–390. [Google Scholar] [CrossRef]
- Schwartz, T.; Hoffmann, S.; Obst, U. Formation of natural biofilms during chlorine dioxide and UV disinfection in a public drinking water distribution system. J. Appl. Microbiol. 2003, 95, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Cervero-Arago, S.; Sommer, R.; Araujo, R.M. Effect of UV irradiation (253.7 nm) on free Legionella and Legionella associated with its amoebae hosts. Water Res. 2014, 67, 299–309. [Google Scholar] [CrossRef]
- Garcia, M.T.; Jones, S.; Pelaz, C.; Millar, R.D.; Abu Kwaik, Y. Acanthamoeba polyphaga resuscitates viable non-culturable Legionella pneumophila after disinfection. Environ. Microbiol. 2007, 9, 1267–1277. [Google Scholar] [CrossRef]
- Loret, J.F.; Robert, S.; Thomas, V.; Cooper, A.J.; McCoy, W.F.; Levi, Y. Comparison of disinfectants for biofilm, protozoa and Legionella control. J. Water Health 2005, 3, 423–433. [Google Scholar] [CrossRef]
- Thomas, V.; Bouchez, T.; Nicolas, V.; Robert, S.; Loret, J.F.; Levi, Y. Amoebae in domestic water systems: Resistance to disinfection treatments and implication in Legionella persistence. J. Appl. Microbiol. 2004, 97, 950–963. [Google Scholar] [CrossRef]
- Critchley, M.; Bentham, R. The efficacy of biocides and other chemical additives in cooling water systems in the control of amoebae. J. Appl. Microbiol. 2009, 106, 784–789. [Google Scholar] [CrossRef]
- Kuchta, J.M.; Navratil, J.S.; Shepherd, M.E.; Wadowsky, R.M.; Dowling, J.N.; States, S.J.; Yee, R.B. Impact of chlorine and heat on the survival of Hartmannella vermiformis and subsequent growth of Legionella pneumophila. Appl. Environ. Microbiol. 1993, 59, 4096–4100. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, M.; Berne, F.; Herbelin, P.; Binet, M.; Berthelot, N.; Rodier, M.H.; Soreau, S.; Hechard, Y. Sensitivity of free-living amoeba trophozoites and cysts to water disinfectants. Int. J. Hyg. Environ. Health 2014, 217, 335–339. [Google Scholar] [CrossRef]
- Mogoa, E.; Bodet, C.; Legube, B.; Hechard, Y. Acanthamoeba castellanii: Cellular changes induced by chlorination. Exp. Parasitol. 2010, 126, 97–102. [Google Scholar] [CrossRef]
- Storey, M.V.; Winiecka-Krusnell, J.; Ashbolt, N.J.; Stenstrom, T.A. The efficacy of heat and chlorine treatment against thermotolerant Acanthamoebae and Legionellae. Scand. J. Infect. Dis. 2004, 36, 656–662. [Google Scholar] [CrossRef]
- Loret, J.; Jousset, M.; Robert, S.; Anselme, C.; Saucedo, G.; Ribas, F.; Martinez, L.; Catalan, V. Elimination of free-living amoebae by drinking water treatment processes. Eur. J. Water Qual. 2008, 39, 37–50. [Google Scholar] [CrossRef]
- Fouque, E.; Hechard, Y.; Hartemann, P.; Humeau, P.; Trouilhe, M.C. Sensitivity of Vermamoeba (Hartmannella) vermiformis cysts to conventional disinfectants and protease. J. Water Health 2015, 13, 302–310. [Google Scholar] [CrossRef]
- Cursons, R.T.; Brown, T.J.; Keys, E.A. Effect of disinfectants on pathogenic free-living amoebae: In axenic conditions. Appl. Environ. Microbiol. 1980, 40, 62–66. [Google Scholar] [CrossRef] [Green Version]
- De Jonckheere, J.; van de Voorde, H. Differences in destruction of cysts of pathogenic and nonpathogenic Naegleria and Acanthamoeba by chlorine. Appl. Environ. Microbiol. 1976, 31, 294–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, H.C.; Wylie, J.; Dejean, G.; Kaksonen, A.H.; Sutton, D.; Braun, K.; Puzon, G.J. Reduced efficiency of chlorine disinfection of Naegleria fowleri in a drinking water distribution biofilm. Environ. Sci. Technol. 2015, 49, 11125–11131. [Google Scholar] [CrossRef] [PubMed]
- Mogoa, E.; Bodet, C.; Morel, F.; Rodier, M.H.; Legube, B.; Hechard, Y. Cellular response of the amoeba Acanthamoeba castellanii to chlorine, chlorine dioxide, and monochloramine treatments. Appl. Environ. Microbiol. 2011, 77, 4974–4980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goudot, S.; Herbelin, P.; Mathieu, L.; Soreau, S.; Banas, S.; Jorand, F.P. Biocidal efficacy of monochloramine against planktonic and biofilm-associated Naegleria fowleri cells. J. Appl. Microbiol. 2014, 116, 1055–1065. [Google Scholar] [CrossRef]
- Chang, S.L. Resistance of pathogenic Naegleria to some common physical and chemical agents. Appl. Environ. Microbiol. 1978, 35, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Coulon, C.; Collignon, A.; McDonnell, G.; Thomas, V. Resistance of Acanthamoeba cysts to disinfection treatments used in health care settings. J. Clin. Microbiol. 2010, 48, 2689–2697. [Google Scholar] [CrossRef] [Green Version]
- Aksozek, A.; McClellan, K.; Howard, K.; Niederkorn, J.Y.; Alizadeh, H. Resistance of Acanthamoeba castellanii cysts to physical, chemical, and radiological conditions. J. Parasitol. 2002, 88, 621–623. [Google Scholar] [CrossRef]
- Sarkar, P.; Gerba, C.P. Inactivation of Naegleria fowleri by chlorine and ultraviolet light. J. Am. Water Work Assoc. 2012, 104, E173–E180. [Google Scholar] [CrossRef]
- Langmark, J.; Storey, M.V.; Ashbolt, N.J.; Stenstrom, T.A. The effects of UV disinfection on distribution pipe biofilm growth and pathogen incidence within the greater Stockholm area, Sweden. Water Res. 2007, 41, 3327–3336. [Google Scholar] [CrossRef]
- Dietersdorfer, E.; Cervero-Arago, S.; Sommer, R.; Kirschner, A.K.; Walochnik, J. Optimized methods for Legionella pneumophila release from its Acanthamoeba hosts. BMC Microbiol. 2016, 16, 74. [Google Scholar] [CrossRef] [Green Version]
- Berk, S.G.; Ting, R.S.; Turner, G.W.; Ashburn, R.J. Production of respirable vesicles containing live Legionella pneumophila cells by two Acanthamoeba spp. Appl. Environ. Microbiol. 1998, 64, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berk, S.G.; Faulkner, G.; Garduno, E.; Joy, M.C.; Ortiz-Jimenez, M.A.; Garduno, R.A. Packaging of live Legionella pneumophila into pellets expelled by Tetrahymena spp. does not require bacterial replication and depends on a Dot/Icm-mediated survival mechanism. Appl. Environ. Microbiol. 2008, 74, 2187–2199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shadrach, W.S.; Rydzewski, K.; Laube, U.; Holland, G.; Ozel, M.; Kiderlen, A.F.; Flieger, A. Balamuthia mandrillaris, free-living ameba and opportunistic agent of encephalitis, is a potential host for Legionella pneumophila bacteria. Appl. Environ. Microbiol. 2005, 71, 2244–2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, J.M.; Rupper, A.; Cardelli, J.A.; Isberg, R.R. Intracellular growth of Legionella pneumophila in Dictyostelium discoideum, a system for genetic analysis of host-pathogen interactions. Infect. Immun. 2000, 68, 2939–2947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newsome, A.L.; Baker, R.L.; Miller, R.D.; Arnold, R.R. Interactions between Naegleria fowleri and Legionella pneumophila. Infect. Immun. 1985, 50, 449–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Nakao, R.; Fujishima, M.; Tachibana, M.; Shimizu, T.; Watarai, M. Ciliate Paramecium is a natural reservoir of Legionella pneumophila. Sci. Rep. 2016, 6, 24322. [Google Scholar] [CrossRef] [PubMed]
- Kao, P.M.; Tung, M.C.; Hsu, B.M.; Hsu, S.Y.; Huang, J.T.; Liu, J.H.; Huang, Y.L. Differential Legionella spp. survival between intracellular and extracellular forms in thermal spring environments. Environ. Sci. Pollut. Res. Int. 2013, 20, 3098–3106. [Google Scholar] [CrossRef]
- International Organization for Standardization. ISO/TS12869:2019 Water Quality-Detection and Quantification of LEGIONELLA spp. and/or Legionella pneumophila by Concentration and Genic amplification by Quantitative Polymerase Chain Reaction (qPCR); International Organization for Standardization: Geneva, Switzerland, 2019. [Google Scholar]
- Qvarnstrom, Y.; Visvesvara, G.S.; Sriram, R.; da Silva, A.J. Multiplex real-time PCR assay for simultaneous detection of Acanthamoeba spp., Balamuthia mandrillaris, and Naegleria fowleri. J. Clin. Microbiol. 2006, 44, 3589–3595. [Google Scholar] [CrossRef] [Green Version]
- Whiley, H.; Taylor, M.; Bentham, R. Detection of Legionella species in potting mixes using fluorescent in situ hybridisation (FISH). J. Microbiol. Methods 2011, 86, 304–309. [Google Scholar] [CrossRef]
- Health Protection Agency. Isolation and Identification of Acanthamoeba Species; Health Protection Agency: London, UK, 2004.
- Standards Australia. AS5132:2017 Waters-Examination for Legionella spp. Includeing Legionella Pneumophila-Using Concentration; Standards Australia: Sydney, Australia, 2017. [Google Scholar]
- Nocker, A.; Cheung, C.Y.; Camper, A.K. Comparison of propidium monoazide with ethidium monoazide for differentiation of live vs. dead bacteria by selective removal of DNA from dead cells. J. Microbiol. Methods 2006, 67, 310–320. [Google Scholar] [CrossRef]
- Yanez, M.A.; Nocker, A.; Soria-Soria, E.; Murtula, R.; Martinez, L.; Catalan, V. Quantification of viable Legionella pneumophila cells using propidium monoazide combined with quantitative PCR. J. Microbiol. Methods 2011, 85, 124–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brescia, C.C.; Griffin, S.M.; Ware, M.W.; Varughese, E.A.; Egorov, A.I.; Villegas, E.N. Cryptosporidium propidium monoazide-PCR, a molecular biology-based technique for genotyping of viable Cryptosporidium oocysts. Appl. Environ. Microbiol. 2009, 75, 6856–6863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fittipaldi, M.; Pino Rodriguez, N.J.; Adrados, B.; Agusti, G.; Penuela, G.; Morato, J.; Codony, F. Discrimination of viable Acanthamoeba castellani trophozoites and cysts by propidium monoazide real-time polymerase chain reaction. J. Eukaryot. Microbiol. 2011, 58, 359–364. [Google Scholar] [CrossRef]
- Dietersdorfer, E.; Kirschner, A.; Schrammel, B.; Ohradanova-Repic, A.; Stockinger, H.; Sommer, R.; Walochnik, J.; Cervero-Arago, S. Starved viable but non-culturable (VBNC) Legionella strains can infect and replicate in amoebae and human macrophages. Water Res. 2018, 141, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, A.K.T. Determination of viable Legionellae in engineered water systems: Do we find what we are looking for? Water Res. 2016, 93, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Cervero-Arago, S.; Rodriguez-Martinez, S.; Canals, O.; Salvado, H.; Araujo, R.M. Effect of thermal treatment on free-living amoeba inactivation. J. Appl. Microbiol. 2014, 116, 728–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohr, U.; Weber, S.; Michel, R.; Selenka, F.; Wilhelm, M. Comparison of free-living amoebae in hot water systems of hospitals with isolates from moist sanitary areas by identifying genera and determining temperature tolerance. Appl. Environ. Microbiol. 1998, 64, 1822–1824. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-S.R.; Sproul, O.J.; Rubin, A.J. Inactivation of Naegleria gruberi cysts by chlorine dioxide. Water Res. 1985, 19, 783–789. [Google Scholar] [CrossRef]


{kind=link}
| Isolation Source (Temperature at Time of Sampling) | Water Treatment Method | L. pneumophila | Potential Protozoan Host | Comments | Country of Origin (Sampling Site) | Reference | ||
|---|---|---|---|---|---|---|---|---|
| Identification Method | Serogroup Sequence-Type | Genus/Species | Identification Method | |||||
| Hospital Settings | ||||||||
| Hot (45–52 °C) water tanks | - | Culturing, co-culture assay and serological identification | SG1 | Hartmannella cantabrigiensis Vermamoeba vermiformis Echinamoeba exudans | Culturing, light and transmission electron microscopy | Nosocomial legionellosis investigation | USA | [31] |
| Potable water sites (39–40 °C) | - | Culturing and monoclonal antibody based serotyping | SG1 | Acanthamoeba hatchetti Hartmannella cantabrigiensis Vermamoeba vermiformis Vahlkampfia Filamoeba nolandi Comandonia operculata Paravahlkampfia ustiana | Culturing and light microscopy | Nosocomial legionellosis investigation Thermal treatment (70 °C) and chlorination (1.5–2.0 mg/L) controlled the bacteria for 6 months but not amoebae. The treatment reduced incidence of legionellosis | South Dakota, USA | [35] |
| Cooling tower, humidifier, hot water tank and supply | - | Culturing and co-culture assay | - | Vermamoeba vermiformis Naegleria | Culturing, light and transmission electron microscopy | - | Paris, France | [32] |
| Hot (39–60 °C) and cold water supply | - | Culturing (ODR: 1 × 103–9.7 × 104 CFU/L), direct fluorescent antibody and monoclonal antibody based serotyping | SG1 SG5 | Hartmannella (Hartmannellidae/limax amoebae) | Culturing and light microscopy | Post nosocomial outbreak surveillance | Halifax, Nova Scotia, Canada | [36] |
| Organ transplant unit hot (mean 56.2 °C) and cold water (mean 16.6 °C) supplies | - | Culturing and serological assay | SG1 | Acanthamoeba Hartmannella Echinamoeba Vahlkampfia Tetrahymena Vannella | Culturing and light microscopy | Population density of amoebae was greater in hot water supplies than cold water supplies Along amoebae other diverse eukaryotic microbes were detected as well | UK | [37] |
| Water supplies | Thermal treatment (60 and 70 °C) | Culturing (Legionella ODR: 2.89–6.74 × 105 CFU/L), co-culture, latex agglutination, indirect and immunofluorescence assays, and PFGE | SG1 SG2 | Acanthamoeba Vahlkampfia Mayorella | Culturing and light microscopy | Thermal treatment (70 °C) only controlled bacterial contamination for 3 months SG1 is more thermotolerant than SG2 at 60 °C | Germany | [38] |
| Water network system (mean 56 °C) | - | Amoebae co-culture assay, PCR and sequencing | - | Vermamoeba vermiformis | Culturing, PCR and sequencing | Detection of thermotolerant Vermamoeba vermiformis | Lausanne, Switzerland | [39] |
| Water distribution system (18.9–32.6 °C) | Chlorine dioxide treatment Thermal treatment (<50 °C) | Culturing (ODR: L. pneumophila SG1: 1 × 102–3.5 × 104 CFU/L and L. pneumophila SG2-14: 1 × 102–4 × 104 CFU/L) and latex agglutination assay | SG1 SG2-14 | Acanthamoeba Hartmannella | Culturing and light microscopy | - | Messina, Italy | [40] |
| Tap water | Chloramine (1.93 ± 1.04 mg/L) treatment | Culturing (protocol: ISO 11731-2:2004, LOD: 1 CFU/100 mL, ODR: 100–1.4 × 105 ± 1.3 × 105 CFU/L), qPCR (LOD: 5 GU, LOQ: 25 GU, Legionella ODR: 100–109 gu/L) and EMA-qPCR | ST269 | Acanthamoeba polyphaga | Culturing, light microscopy and PCR | Italy | [27] | |
| Cold (14.9 °C) and warm (45.1 °C) potable water | Thermal treatment, chlorination (hypochlorates, chloramine), bacterial filters and chlorine dioxide treatment | Culturing (protocols: ISO 11731:1998 and ISO 11731-2:2004, LOD: 1 CFU/100 mL, ODR: 0–3 × 103 CFU/100 mL) and MALDI-TOF MS | - | Acanthamoeba Vermamoeba vermiformis | Culturing and light microscopy | - | Bratislava, Slovakia | [41] |
| Cold water system (20–27.3 °C) | Chlorine contents 0.01–0.32 mg/L | qPCR (protocol: ISO/TS 12869:2012, LOD: 5 GU, LOQ: 25 GU, ODR: 2.7–3.8 × 102 gu/L) | - | Acanthamoeba Vermamoeba vermiformis | Culturing and light microscopy | Johannesburg, South Africa | [42] | |
| Dental unit waterlines | H2O2 treatment (occasionally) | Heterotrophic plate counts, culturing (protocol: ISO 11731-2:2004, LOD: 1 CFU/100 mL, ODR: 0–2700 CFU/L) and agglutination test | - | Vermamoeba vermiformis | Culturing, light microscopy, PCR and sequencing | Italy | [43] | |
| Potable Water System | ||||||||
| Unchlorinated water supplies (9.5–13.5 °C) | - | qPCR | - | Acanthamoeba Acanthamoeba polyphaga Vermamoeba vermiformis | qPCR (LOD: 1 cell/reaction), T-RFLP, cloning and sequencing | Along amoebae other diverse eukaryotic microbes were detected as well | Netherlands | [44] |
| Ground water supplies (5–39 °C) | Aeration, lime stone, granular activated carbon slow sand and rapid sand filtration, ozonisation and pellet softening | Culturing, biofilm batch test and qPCR | - | Acanthamoeba Vermamoeba vermiformis | 18S rDNA sequencing, PCR, T-RFLP and sequencing | Along amoebae other eukaryotic microbes were detected as well | Netherlands | [45] |
| Water supplies (mean 30 °C) | Reverse osmosis, distillation (82%), chlorination (<0.005–0.2 mg/L), dolomite, limestone and granular activated carbon filtration, fluoride addition (0.3–0.7 mg/L), UV treatment (7.5–35.99 mJ/cm2) | Culturing (LOD: 250 CFU/L, Legionella ODR: 2.5 × 102–2.5 × 105 CFU/L) and latex agglutination assay | - | Acanthamoeba Vermamoeba vermiformis Echinamoeba exundans Echinamoeba thermarum Neoparamoeba | qPCR (LOD: 2 C/L, ODR: Acanthamoeba < 2–56 C/L and V. vermiformis < 2–1670 C/L) | - | Caribbean islands, Leeward Antilles | [46] |
| Water distribution systems (mean 37.3 ± 8.4 °C) | Chloramine treatment (Chlorine contents 1.8 mg/L), flocculation, sedimentation, and dual-medium filtration | Culturing, qPCR (LOQ: 1–10 copies/reaction, maximum ODR: 13.7 ± 5.1 gc/mL) and T-RFLP | - | Acanthamoeba Vermamoeba vermiformis | qPCR (LOQ: 1–10 copies/reaction, maximum ODR: Acanthamoeba 6.8 ± 2.9 gc/mL and V. vermiformis 7.1 × 104 ± 4.4 × 103 gc/mL) | High concentration of chloramine is unable to disinfect water | Southwest Virginia, USA | [47] |
| Water treatment plant (7–21 °C) | - | Multiplex PCR | - | Vermamoeba vermiformis | Culturing, light microscopy, PCR and sequencing | Amoebae were frequently detected at 17 °C | Aragon, Spain | [33] |
| Water treatment facility (25 ± 3.4–28.2 ± 1.1 °C) | - | PCR (Legionella ODR: 1.2 × 104–2.4 × 105 gc/L) and sequencing | - | Acanthamoeba Vermamoeba vermiformis Naegleria | Culturing, PCR, qPCR (ODR: Acanthamoeba 2.1 × 102–7.7 × 102 gc/L and Naegleria 7.6 × 102–9.4 × 102 gc/L) and sequencing | - | Kaoping River, Taiwan | [48] |
| Sediments of municipal water storage tank (2.2–28.9 °C) | Chlorination (<4 mg/L) | qPCR (LOD: 2 CE/reaction, Legionella ODR: 51 ± 114–7.98 × 104 ± 2.49 × 104 CE/g), cloning and sequencing | SG1 | Acanthamoeba Vermamoeba vermiformis | qPCR (LOD: 2 CE/reaction, ODR: Acanthamoeba 22 ± 50–391 ± 243 CE/g and V. vermiformis 17 ± 23 CE/g), cloning and sequencing | - | Northeast, East Coast, Midwest, South and West Coast, USA | [49] |
| Water distribution system | - | qPCR (LOD: 2 CE/reaction, Legionella ODR: 2 ± 4–391 ± 17 CE/L), cloning and sequencing | - | Acanthamoeba Acanthamoeba castellanii Vermamoeba vermiformis | qPCR (LOD: 2 CE/reaction, ODR: Acanthamoeba 1 ± 2–16 ± 2 * CE/L and V. vermiformis 1 ± 1–9 ± 11 * CE/L), cloning and sequencing | - | USA | [50] |
| Domestic water systems (mean 20.6 ± 3.8 °C) | - | Culturing, co-culture assay, PCR and sequencing | - | Vermamoeba vermiformis | Culturing, light microscopy, PCR and sequencing | - | Geneva, Lausanne and Sion, Switzerland | [51] |
| Sediments of water storage tank | - | qPCR (ODR: 25 ± 51–300 ± 38 gn/g) and NGS | - | Acanthamoeba Vermamoeba vermiformis | qPCR (ODR: Acanthamoeba 3–7 gn/g, V. vermiformis 99 ± 43–120 ± 60 gn/g) and NGS | - | Ohio, West Virginia and Texas, USA | [52] |
| Potable water | Polyaluminium chloride coagulation, sedimentation, sand and biologically activated carbon filtration and chlorination | qPCR (LOQ: 1–10 copy/reaction, minimum ODR: 3.5 log(gc)/mL) | - | Acanthamoeba Vermamoeba vermiformis | qPCR (LOD: 1–10 copy/reaction, minimum ODR: 2 log(gc)/mL for V. vermiformis and 4 log(gc)/mL for Acanthamoeba) and sequencing | Antibiotics (sulfadiazine and ciprofloxacin) promote growth of both bacterium and amoebae | Northern China | [53] |
| Potable water | Polyaluminium chloride coagulation, sedimentation, sand and biologically activated carbon filtration, chlorination and ozonisation | qPCR (LOQ: 1–10 copies/reaction, minimum ODR ≈ 1 log(gc)/g) | - | Acanthamoeba Naegleria | qPCR (LOQ: 1–10 copies/reaction, minimum ODR: ≈ 0.5 log(gc)/g for Naegleria and ≈ 1 log(gc)/g for Acanthamoeba) | Combined chlorination and ozonisation are effective than chlorination only | Northern China | [54] |
| Potable water | Coagulation, ozonisation, pellet softening, granular activated carbon filtration, rapid and slow sand filtration | Heterotrophic plate counts, culturing (protocol: NEN 6275, LOD: 1 log(CFU)/cm2) epifluorescence microscopy, bioluminescence assay, PCR and sequencing | - | Vermamoeba vermiformis | qPCR (ODR: 0.7–384 CE/cm2) | - | Netherlands | [55] |
| Residential secondary water supply systems (13.9 ± 4.0–17.4 ± 2.9 °C) | Chloramine treatment (Chlorine contents 0.19–0.89 mg/L) | qPCR (LOQ: 10 copies/reaction, maximum ODR: ≈ 102 gc/mL) and sequencing | - | Acanthamoeba Vermamoeba vermiformis | qPCR (LOQ: 10 copies/reaction, ODR: 101–103 gc/mL for both Acanthamoeba and V. vermiformis) and sequencing | - | Shanghai, China | [56] |
| Water treatment facility | Coagulation, sedimentation, chlorination, ozonisation, granular activated carbon and sand filtration | qPCR (LOQ: 10 copies/reaction, minimum ODR: 102 log(gc)/mL) and sequencing | - | Vermamoeba vermiformis | qPCR (LOQ: 10 copies/reaction) and sequencing | Sand filtration after granular activated carbon treatment improves water quality | Southeast China | [57] |
| Water from private wells after flood | - | Culturing (protocol: ISO 11731, LOD: 1 CFU/100 mL) and qPCR (LOQ: 9.5 gc/mL, maximum ODR: 52.4 gc/mL) | - | Naegleria fowleri | qPCR (ODR: 11–610 gc/mL) | - | Louisiana, USA | [58] |
| Potable water | - | Culturing and DVC-FISH | - | Acanthamoeba Vermamoeba vermiformis | Culturing and PCR | - | Valencia, Spain | [34] |
| Hospital Settings | Potable Water System |
|---|---|
| Phylum: Amoebozoa Class: Tubulinea Genera: Vermamoeba, Echinamoeba, Hartmannella, Filamoeba Class: Discosea Genera: Acanthamoeba, Comandonia, Mayorella, Vannella Class: Heterolobosea Genera: Vahlkampfia, Paravahlkampfia Phylum: Percolozoa Class: Heterolobosea Genus: Naegleria Phylum: Ciliophora Class: Oligohymenophorea Genus: Tetrahymena | Phylum: Amoebozoa Class: Tubulinea Genera: Vermamoeba, Echinamoeba Class: Discosea Genera: Acanthamoeba, Neoparamoeba Phylum: Percolozoa Class: Heterolobosea Genus: Naegleria |
| Organisms | Disinfectant Dose | |||||
|---|---|---|---|---|---|---|
| Temperature (°C) | Chlorine (mg-min/L) | Monochloramine (mg-min/L) | Chlorine Dioxide (mg-min/L) | Ozone (mg-min/L) | UV Rays (mJ/cm2) | |
| Legionella pneumophila studies | ||||||
| Legionella pneumophila1 | 70 °C [35,38] | 6 mg/L/6 h (5 log reduction) [66] | 17 (3 log reduction) [69] | 0.4 (3 log reduction) [69] | 1–2 mg/L/5 h (5 log reduction) [66] | 30 (5 log reduction) 2 [66] |
| Legionella pneumophila–potential host protozoans coculture studies | ||||||
| Legionella pneumophila Acanthamoeba coculture | 93 °C 3 [63] | >50 mg/L [67] | 23 (3 log reduction) [69] | 2.8 (3 log reduction) [69] | - | 10.8 (4 log reduction) [73] |
| Legionella pneumophila Vermamoeba coculture | 58 °C [64] | - | - | - | - | - |
| Potential host protozoans studies | ||||||
| Acanthamoeba (trophozoite) | 65 °C/10 min (inactivation) [90] | 28 (2 log reduction) [79] | 40 (2 log reduction) [69] | >5 (2 log reduction) [79] | 10 (3 log reduction) [82] | 72.2 (3 log reduction) [73] |
| Acanthamoeba (cyst) | 80 °C/10 min [81] | 3500 (4 log reduction) [82] | 352 (2 log reduction) [79] | 80 (4 log reduction) [82] | 15 (4 log reduction) [82] | 800 [91] |
| Vermamoeba (trophozoite) | 60 °C/5 min (4 log reduction) [113] | 2–4 mg/L/30 min inactivation) [78] | - | - | - | 26 (3 log reduction) [73] |
| Vermamoeba (cyst) | 60 °C/5 min (2 log reduction) [113] | 15 mg/L/10 min (inactivation) [83] | - | - | - | 76.2 (3 log reduction) [73] |
| Hartmannella (trophozoite) | 53 °C [114] | 15 (2 log reduction) [79] | 12 2 log reduction) [79] | 5 (2 log reduction) [79] | - | - |
| Hartmannella (cyst) | - | 156 (2 log reduction) [79] | 34 (2 log reduction) [79] | 1 (2 log reduction) [79] | - | - |
| Naegleria (trophozoite) | 55 °C/15 min [89] | 5 (2 log reduction) [79] | 4–17 (2 log reduction) [88] | 1 (2 log reduction) [79] | 6.75 mg/L 30 min (3 log reduction) [84] | 24 (4 log reduction) [92] |
| Naegleria (cyst) | 65 °C/3 min [89] | 29 (2 log reduction) [79] | 13 (2 log reduction) [79] | 5.5 (2 log reduction) [115] | - | 121 (4 log reduction) [92] |
| Vahlkampfia (trophozoite) | - | 1 mg/L (inactivation) [77] | - | - | - | - |
| Vahlkampfia (cyst) | - | 2 mg/L/2 h (3 log reduction) [77] | - | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nisar, M.A.; Ross, K.E.; Brown, M.H.; Bentham, R.; Whiley, H. Legionella pneumophila and Protozoan Hosts: Implications for the Control of Hospital and Potable Water Systems. Pathogens 2020, 9, 286. https://doi.org/10.3390/pathogens9040286
Nisar MA, Ross KE, Brown MH, Bentham R, Whiley H. Legionella pneumophila and Protozoan Hosts: Implications for the Control of Hospital and Potable Water Systems. Pathogens. 2020; 9(4):286. https://doi.org/10.3390/pathogens9040286
Chicago/Turabian StyleNisar, Muhammad Atif, Kirstin E. Ross, Melissa H. Brown, Richard Bentham, and Harriet Whiley. 2020. "Legionella pneumophila and Protozoan Hosts: Implications for the Control of Hospital and Potable Water Systems" Pathogens 9, no. 4: 286. https://doi.org/10.3390/pathogens9040286