Distribution and Nucleotide Diversity of Yr15 in Wild Emmer Populations and Chinese Wheat Germplasm

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Distribution of Yr15 in Wild Emmer Populations
2.2. Characterization of Yr15 in Chinese Wheat Germplasm
2.3. Haplotype Diversity of WTK1
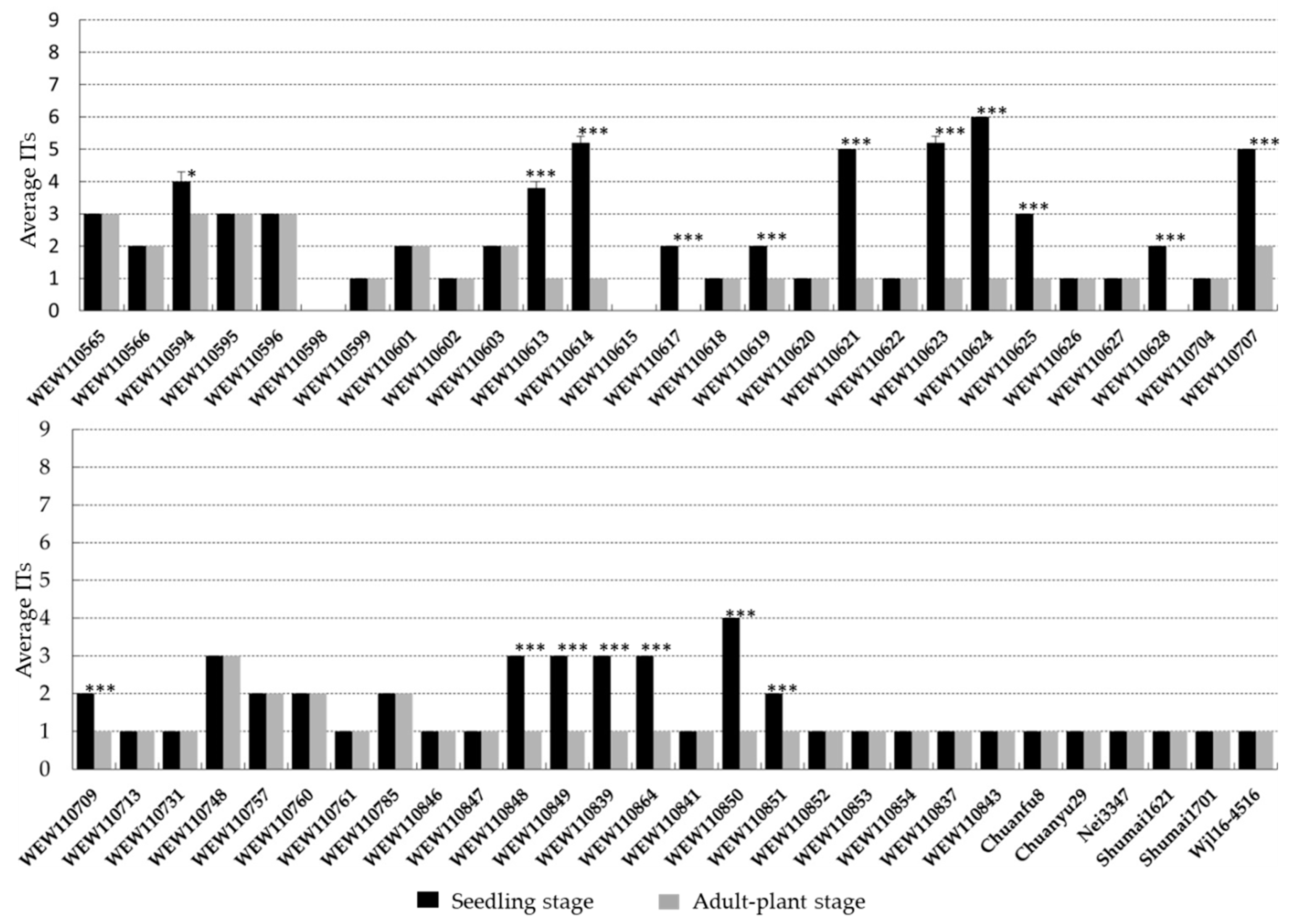
2.4. Stripe Rust Resistance in Wheat Accessions That Carry Yr15
3. Discussion
3.1. The Geographic Distibution of Yr15 in Wild Emmer Populations
3.2. Sequence Diversity of the WTK1
3.3. Stripe Rust Resistance Variation in Wild Emmer Accessions Carrying Yr15
3.4. The Potential Value of Yr15 Gene in Wheat Stripe Rust Resistance Breeding Programs in China
4. Materials and Methods
4.1. Plant Material
4.2. Analysis of Presence/absence Polymorphism in Yr15
4.3. Sequence Analysis of WTK1
4.4. Stripe Rust Resistance Response
5. Patents
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Chen, X.M. Epidemiology and control of stripe rust Puccinia striiformis f. sp tritici on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Yuan, C.L.; Wu, J.Z.; Yan, B.Q.; Hao, Q.Q.; Zhang, C.Z.; Lyu, B.; Ni, F.; Caplan, A.; Wu, J.J.; Fu, D.L. Remapping of the stripe rust resistance gene Yr10 in common wheat. Theor. Appl. Genet. 2018, 131, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- De Vallavieille-Pope, C.; Ali, S.; Leconte, M.; Enjalbert, J.; Delos, M.; Rouzet, J. Virulence dynamics and regional structuring of Puccinia striiformis f. sp.tritici in France between 1984 and 2009. Plant Dis. 2012, 96, 131–140. [Google Scholar] [PubMed] [Green Version]
- Dangl, J.L.; Jones, J.D. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K. Durable resistance of crops to disease: A Darwinian perspective. Annu. Rev. Phytopathol. 2015, 53, 513–539. [Google Scholar] [CrossRef]
- Fu, D.; Uauy, C.; Distelfeld, A.; Blechl, A.; Epstein, L.; Chen, X.M.; Sela, H.; Fahima, T.; Dubcovsky, J. A kinase-START gene confers temperature-dependent resistance to wheat stripe rust. Science 2009, 323, 1357–1360. [Google Scholar] [CrossRef] [Green Version]
- Krattinger, S.G.; Lagudah, E.S.; Spielmeyer, W.; Singh, R.P.; Huerta-Espino, J.; McFadden, H.; Bossolini, E.; Selter, L.L.; Keller, B. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 2009, 323, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Xing, L.P.; Hu, P.; Liu, J.Q.; Witek, K.; Zhou, S.; Xu, J.F.; Zhou, W.H.; Gao, L.; Huang, Z.P.; Zhang, R.Q.; et al. Pm21 from Haynaldia villosa encodes a CC-NBS-LRR protein conferring powdery mildew resistance in wheat. Mol. Plant 2018, 11, 874–878. [Google Scholar] [CrossRef] [Green Version]
- Sela, H.; Loutre, C.; Keller, B.; Schulman, A.; Nevo, E.; Korol, A.; Fahima, T. Rapid linkage disequilibrium decay in the Lr10 gene in wild emmer wheat (Triticum dicoccoides) populations. Theor. Appl. Genet. 2011, 122, 175–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sela, H.; Spiridon, L.N.; Ashkenazi, H.; Bhullar, N.K.; Brunner, S.; Petrescu, A.J.; Fahima, T.; Keller, B.; Jordan, T. Three-dimensional modeling and diversity analysis reveals distinct AVR recognition sites and evolutionary pathways in wild and domesticated wheat Pm3 R genes. Mol. Plant Microbe Interact. 2014, 27, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Raats, D.; Sela, H.; Klymiuk, V.; Lidzbarsky, G.; Feng, L.H.; Krugman, T.; Fahima, T. Evolution and adaptation of wild emmer wheat populations to biotic and abiotic stresses. Annu. Rev. Phytopathol. 2016, 54, 279–301. [Google Scholar] [CrossRef] [PubMed]
- Gou, J.Y.; Li, K.; Wu, K.T.; Wang, X.D.; Lin, H.Q.; Cantu, D.; Uauy, C.; Dobon-Alonso, A.; Midorikawa, T.; Inoue, K.; et al. Wheat stripe rust resistance protein WKS1 reduces the ability of the thylakoid-associated ascorbate peroxidase to detoxify reactive oxygen species. Plant Cell 2015, 27, 1755–1770. [Google Scholar] [CrossRef] [PubMed]
- Krattinger, S.G.; Kang, J.; Braunlich, S.; Boni, R.; Chauhan, H.; Setter, L.L.; Robinson, M.D.; Schmid, M.W.; Wiederhold, E.; Hensel, G.; et al. Abscisic acid is a substrate of the ABC transporter encoded by the durable wheat disease resistance gene Lr34. New Phytol. 2019, 223, 853–866. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Li, Q.P.; Wang, J.; Yan, Y.; Zhang, G.L.; Zhang, H.; Wu, J.; Chen, F.; Wang, X.; Kang, Z.S.; et al. YR36/WKS1-mediated phosphorylation of PsbO, an extrinsic member of photosystem II, inhibits photosynthesis and confers stripe rust resistance in wheat. Mol. Plant 2019, 12, 1639–1650. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Kang, Z.S. Stripe Rust. Springer Science+Business Media: Berlin, Germany, 2017; p. 409. [Google Scholar]
- Gerechter-Amitai, Z.K.; van Silfhout, C.H.; Grama, A.; Kleitman, F. Yr15–a new gene for resistance to Puccinia striiformis in Triticum dicoccoides sel. G-25. Euphytica 1989, 43, 187–190. [Google Scholar] [CrossRef]
- Yaniv, E.; Raats, D.; Ronin, Y.; Korol, A.B.; Grama, A.; Bariana, H.; Dubcovsky, J.; Schulman, A.H.; Fahima, T. Evaluation of marker-assisted selection for the stripe rust resistance gene Yr15, introgressed from wild emmer wheat. Mol. Breed. 2015, 35, 43. [Google Scholar] [CrossRef]
- Klymiuk, V.; Yaniv, E.; Huang, L.; Raats, D.; Fatiukha, A.; Chen, S.; Feng, L.; Frenkel, Z.; Krugman, T.; Lidzbarsky, G.; et al. Cloning of the wheat Yr15 resistance gene sheds light on the plant tandem kinase-pseudokinase family. Nat. Commun. 2018, 9, 3735. [Google Scholar] [CrossRef] [Green Version]
- Klymiuk, V.; Fatiukha, A.; Fahima, T. Wheat tandem kinases provide insights on disease-resistance gene flow and host-parasite co-evolution. Plant J. 2019, 98, 667–679. [Google Scholar] [CrossRef]
- Chen, S.S.; Rouse, M.; Zhang, W.J.; Zhang, X.Q.; Guo, Y.; Briggs, J.; Dubcovsky, J. Wheat gene Sr60 encodes a protein with two putative kinase domains that confers resistance to stem rust. New Phytol. 2020, 225, 948–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, P.; Guo, L.; Wang, Z.Z.; Li, B.B.; Li, J.; Li, Y.H.; Qiu, D.; Shi, W.Q.; Yang, L.J.; Wang, N.; et al. A rare gain of function mutation in a wheat tandem kinase confers resistance to powdery mildew. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.C.; Yang, Z.L.; You, F.M.; Kawahara, T.; Waines, J.G.; Dvorak, J. The structure of wild and domesticated emmer wheat populations, gene flow between them, and the site of emmer domestication. Theor. Appl. Genet. 2007, 114, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, H.; Brandolini, A.; Schafer-Pregl, R.; Salamini, F. AFLP analysis of a collection of tetraploid wheats indicates the origin of emmer and hard wheat domestication in southeast Turkey. Mol. Biol. Evol. 2002, 19, 1797–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nave, M.; Avni, R.; Cakir, E.; Portnoy, V.; Sela, H.; Pourkheirandish, M.; Ozkan, H.; Hale, I.; Komatsuda, T.; Dvorak, J.; et al. Wheat domestication in light of haplotype analyses of the Brittle rachis 1 genes (BTR1-A and BTR1-B). Plant Sci. 2019, 285, 193–199. [Google Scholar] [CrossRef]
- Huang, L.; Sela, H.; Feng, L.; Chen, Q.; Krugman, T.; Yan, J.; Dubcovsky, J.; Fahima, T. Distribution and haplotype diversity of WKS resistance genes in wild emmer wheat natural populations. Theor. Appl. Genet. 2016, 129, 921–934. [Google Scholar] [CrossRef] [Green Version]
- Kato, K.; Tanizoe, C.; Beiles, A.; Nevo, E. Geographical variation in heading traits in wild emmer wheat, Triticum dicoccoides. II. Variation in heading date and adaptation to diverse eco-geographical conditions. Hereditas 1998, 128, 33–39. [Google Scholar] [CrossRef]
- Huang, L.; Feng, L.H.; He, Y.; Tang, Z.Z.; He, J.S.; Sela, H.; Krugman, T.; Fahima, T.; Liu, D.C.; Wu, B.H. Variation in stripe rust resistance and morphological traits in wild emmer wheat populations. Agronomy 2019, 9, 44. [Google Scholar] [CrossRef] [Green Version]
- Nevo, E.; Beiles, A. Genetic diversity of wild emmer wheat in Israel and Turkey-structure, evolution, and application in breeding. Theor. Appl. Genet. 1989, 77, 421–455. [Google Scholar] [CrossRef]
- Haudry, A.; Cenci, A.; Ravel, C.; Bataillon, T.; Brunel, D.; Poncet, C.; Hochu, I.; Poirier, S.; Santoni, S.; David, J. Grinding up wheat: A massive loss of nucleotide diversity since domestication. Mol. Biol. Evol. 2007, 24, 1506–1517. [Google Scholar] [CrossRef] [Green Version]
- Bergelson, J.; Kreitman, M.; Stahl, E.A.; Tian, D. Evolutionary dynamics of plant R-genes. Science 2001, 292, 2281–2285. [Google Scholar] [CrossRef] [Green Version]
- Ersoz, E.S.; Wright, M.H.; Gonzalez-Martinez, S.C.; Langley, C.H.; Neale, D.B. Evolution of disease response genes in loblolly pine: Insights from candidate genes. PLoS ONE 2010, 5, e14234. [Google Scholar] [CrossRef] [Green Version]
- Krattinger, S.G.; Jordan, D.R.; Mace, E.S.; Raghavan, C.; Luo, M.C.; Keller, B.; Lagudah, E.S. Recent emergence of the wheat Lr34 multi-pathogen resistance: Insights from haplotype analysis in wheat, rice, sorghum and Aegilops tauschii. Theor. Appl. Genet. 2013, 126, 663–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klymiuk, V.; Fatiukha, A.; Raats, D.; Bocharova, V.; Huang, L.; Feng, L.H.; Jaiwar, S.; Pozniak, C.; Coaker, G.; Dubcovsky, J.; et al. Three previously characterized resistances to yellow rust are encoded by a single locus Wtk1. J. Exp. Bot. 2020, eraa020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurni, S.; Brunner, S.; Stirnweis, D.; Herren, G.; Peditto, D.; McIntosh, R.A.; Keller, B. The powdery mildew resistance gene Pm8 derived from rye is suppressed by its wheat ortholog Pm3. Plant J. 2014, 79, 904–913. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Dodds, P.; Pryor, T. Structure, function and evolution of plant disease resistance genes. Curr. Opin. Plant Biol. 2000, 3, 278–284. [Google Scholar] [CrossRef]
- Zeng, Q.D.; Han, D.J.; Wang, Q.L.; Yuan, F.P.; Wu, J.H.; Zhang, L.; Wang, X.J.; Huang, L.L.; Chen, X.M.; Kang, Z.S. Stripe rust resistance and genes in Chinese wheat cultivars and breeding lines. Euphytica 2014, 196, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Krugman, T.; Nevo, E.; Beharav, A.; Sela, H.; Fahima, T. The Institute of Evolution Wild Cereal Gene Bank at the University of Haifa. Isr. J. Plant Sci. 2018, 65, 129–146. [Google Scholar] [CrossRef]
- Zhou, Y.; Tang, H.; Cheng, M.P.; Dankwa, K.O.; Chen, Z.X.; Li, Z.Y.; Gao, S.; Liu, Y.X.; Jiang, Q.T.; Lan, X.J.; et al. Genome-wide association study for pre-harvest sprouting resistance in a large germplasm collection of Chinese wheat landraces. Front. Plant Sci. 2017, 8, 401. [Google Scholar] [CrossRef] [Green Version]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol. 1985, 5, 69–76. [Google Scholar] [CrossRef]
- Appels, R.; Eversole, K.; Feuillet, C.; Keller, B.; Rogers, J.; Stein, N.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; Poland, J.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, 7191. [Google Scholar]
- Avni, R.; Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Jordan, K.W. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 2017, 357, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.C.; Tang, X.R.; Wu, J.H.; Shen, C.; Dai, M.F.; Wang, Q.L.; Zeng, Q.D.; Kang, Z.S.; Wu, Y.F.; Han, D.J. Stripe rust resistance to a burgeoning Puccinia striiformis f. sp. tritici race CYR34 in current Chinese wheat cultivars for breeding and research. Euphytica 2019, 215, 68. [Google Scholar]
- Line, R.F.; Qayoum, A. Virulence, Aggressiveness, Evolution, and Distribution of Races of Puccinia striiformis (the Causes of Stripe Rust of Wheat) in North. America, 1968-1987; Technical Bulletin 1788; United State Departmentof Agriculture: Washington, DC, USA, 1992.




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Feng, L.; Jiang, Y.; Zhang, L.; Yan, J.; Zhao, G.; Wang, J.; Chen, G.; Wu, B.; Liu, D.; et al. Distribution and Nucleotide Diversity of Yr15 in Wild Emmer Populations and Chinese Wheat Germplasm. Pathogens 2020, 9, 212. https://doi.org/10.3390/pathogens9030212
He Y, Feng L, Jiang Y, Zhang L, Yan J, Zhao G, Wang J, Chen G, Wu B, Liu D, et al. Distribution and Nucleotide Diversity of Yr15 in Wild Emmer Populations and Chinese Wheat Germplasm. Pathogens. 2020; 9(3):212. https://doi.org/10.3390/pathogens9030212
Chicago/Turabian StyleHe, Yu, Lihua Feng, Yun Jiang, Lianquan Zhang, Jun Yan, Gang Zhao, Jirui Wang, Guoyue Chen, Bihua Wu, Dengcai Liu, and et al. 2020. "Distribution and Nucleotide Diversity of Yr15 in Wild Emmer Populations and Chinese Wheat Germplasm" Pathogens 9, no. 3: 212. https://doi.org/10.3390/pathogens9030212