Dissemination of Piscine orthoreovirus-1 (PRV-1) in Atlantic Salmon (Salmo salar) during the Early and Regenerating Phases of Infection

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. PRV-1 in Blood Cells
2.2. PRV-1 Load in Plasma
2.3. PRV-1 Load in the Heart
2.4. Histological Findings
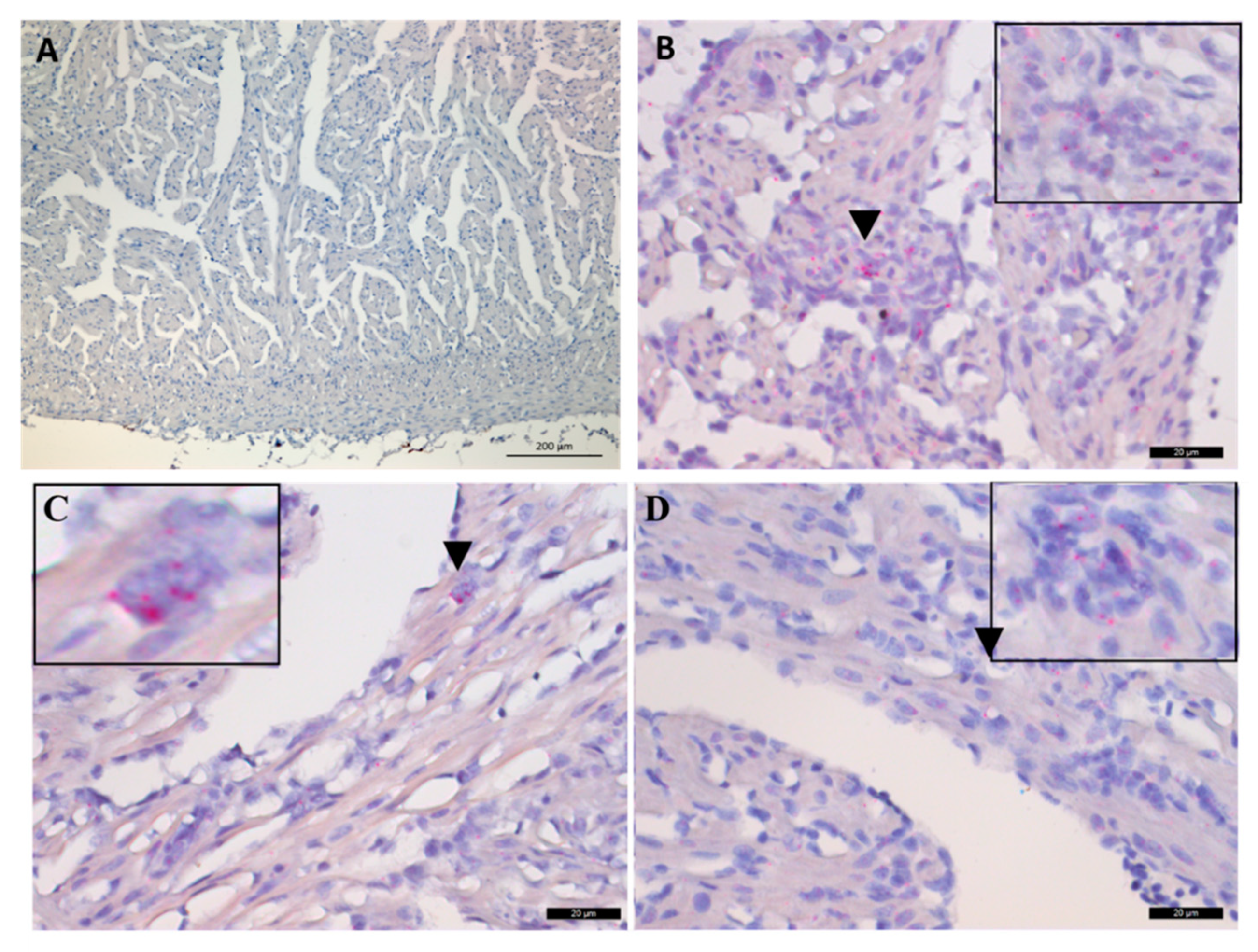
2.5. In Situ Hybridization of Arginase-2 in the Heart
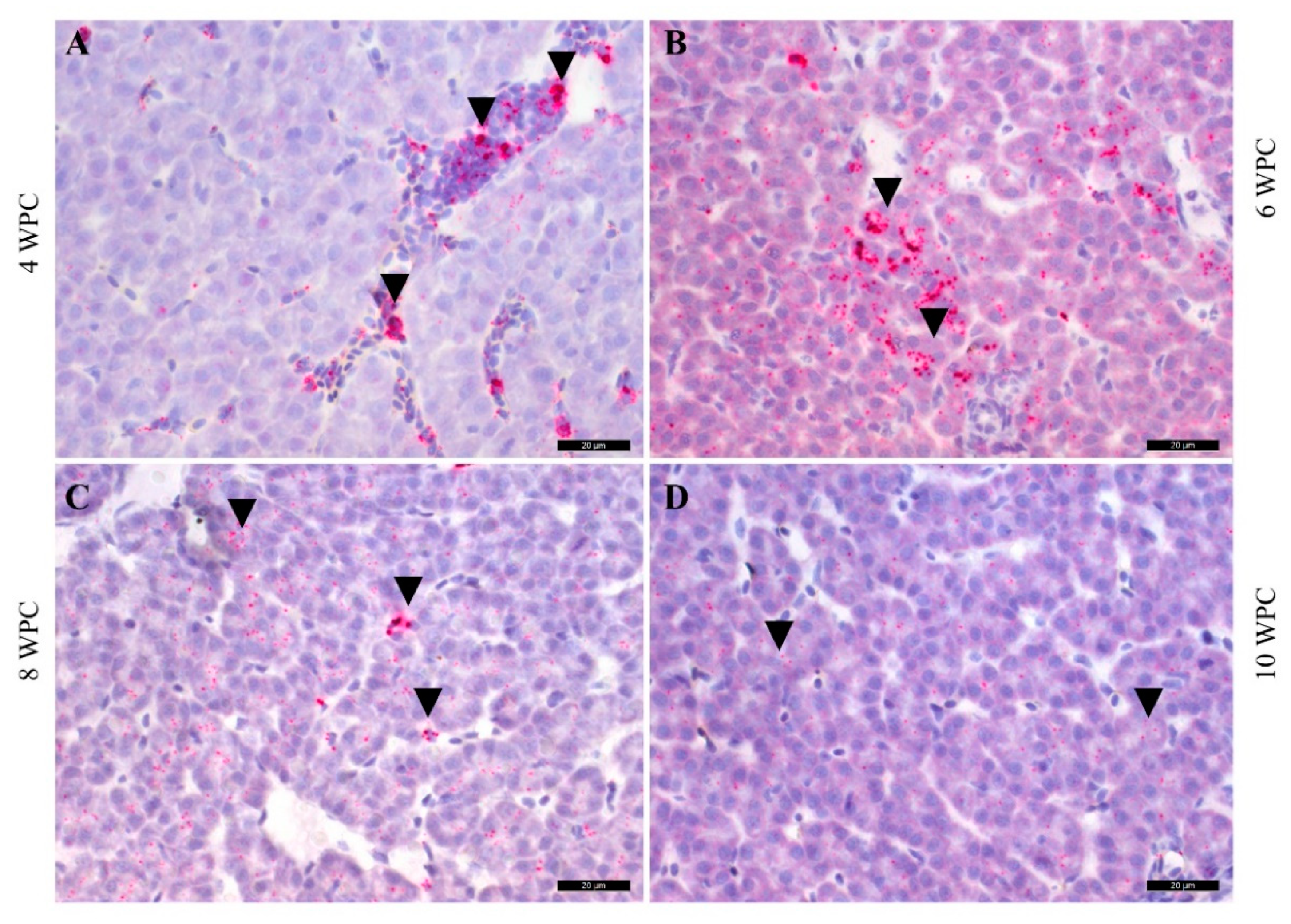
2.6. PRV-1 Load in the Liver
3. Discussion
4. Materials and Methods
4.1. Challenge Experiment
4.2. RNA Isolation and RT-qPCR
4.3. Western Blot
4.4. Flow Cytometry
4.5. Histology and Immunohistochemistry
4.6. In Situ Hybridization
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethics Approval
References
- Hjeltnes, B.; Bang-Jensen, B.; Bornø, G.; Haukaas, A.; Wlade, C.S. (Eds.) The Fish Health Report 2017; The Norwegian Veterinary Institute: Oslo, Norway, 2018. [Google Scholar]
- Wessel, O.; Braaen, S.; Alarcon, M.; Haatveit, H.; Roos, N.; Markussen, T.; Tengs, T.; Dahle, M.K.; Rimstad, E. Infection with purified piscine orthoreovirus demonstrates a causal relationship with heart and skeletal muscle inflammation in atlantic salmon. PLoS ONE 2017, 12, e0183781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finstad, O.W.; Dahle, M.K.; Lindholm, T.H.; Nyman, I.B.; Lovoll, M.; Wallace, C.; Olsen, C.M.; Storset, A.K.; Rimstad, E. Piscine orthoreovirus (prv) infects atlantic salmon erythrocytes. Vet. Res. 2014, 45, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garseth, Å.H.; Ekrem, T.; Biering, E. Phylogenetic evidence of long distance dispersal and transmission of piscine reovirus (prv) between farmed and wild atlantic salmon. PLoS ONE 2013, 8, e82202. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Lovoll, M.; Tengs, T.; Hornig, M.; Hutchison, S.; Hui, J.; Kongtorp, R.T.; Savji, N.; Bussetti, A.V.; Solovyov, A.; et al. Heart and skeletal muscle inflammation of farmed salmon is associated with infection with a novel reovirus. PLoS ONE 2010, 5, e11487. [Google Scholar] [CrossRef]
- Godoy, M.G.; Kibenge, M.J.; Wang, Y.; Suarez, R.; Leiva, C.; Vallejos, F.; Kibenge, F.S. First description of clinical presentation of piscine orthoreovirus (prv) infections in salmonid aquaculture in chile and identification of a second genotype (genotype ii) of prv. Virol. J. 2016, 13, 98. [Google Scholar] [CrossRef]
- Purcell, M.K.; Powers, R.L.; Evered, J.; Kerwin, J.; Meyers, T.R.; Stewart, B.; Winton, J.R. Molecular testing of adult pacific salmon and trout (oncorhynchus spp.) for several rna viruses demonstrates widespread distribution of piscine orthoreovirus in alaska and washington. J. Fish Dis. 2018, 41, 347–355. [Google Scholar] [CrossRef]
- Takano, T.; Nawata, A.; Sakai, T.; Matsuyama, T.; Ito, T.; Kurita, J.; Terashima, S.; Yasuike, M.; Nakamura, Y.; Fujiwara, A.; et al. Full-genome sequencing and confirmation of the causative agent of erythrocytic inclusion body syndrome in coho salmon identifies a new type of piscine orthoreovirus. PLoS ONE 2016, 11, e0165424. [Google Scholar] [CrossRef] [Green Version]
- Vendramin, N.; Dhamotharan, K.; Olsen, A.B.; Cuenca, A.; Teige, L.H.; Wessel, O.; Iburg, T.M.; Dahle, M.K.; Rimstad, E.; Olesen, N.J. Piscine orthoreovirus subtype 3 (prv-3) causes heart inflammation in rainbow trout (oncorhynchus mykiss). Vet. Res. 2019, 50, 14. [Google Scholar] [CrossRef] [Green Version]
- Kongtorp, R.T.; Kjerstad, A.; Taksdal, T.; Guttvik, A.; Falk, K. Heart and skeletal muscle inflammation in atlantic salmon, Salmo salar L.: A new infectious disease. J. Fish Dis. 2004, 27, 351–358. [Google Scholar] [CrossRef]
- Bjorgen, H.; Wessel, O.; Fjelldal, P.G.; Hansen, T.; Sveier, H.; Saebo, H.R.; Enger, K.B.; Monsen, E.; Kvellestad, A.; Rimstad, E.; et al. Piscine orthoreovirus (prv) in red and melanised foci in white muscle of atlantic salmon (salmo salar). Vet. Res. 2015, 46, 89. [Google Scholar] [CrossRef] [Green Version]
- Finstad, O.W.; Falk, K.; Lovoll, M.; Evensen, O.; Rimstad, E. Immunohistochemical detection of piscine reovirus (prv) in hearts of atlantic salmon coincide with the course of heart and skeletal muscle inflammation (hsmi). Vet. Res. 2012, 43, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haatveit, H.M.; Wessel, O.; Markussen, T.; Lund, M.; Thiede, B.; Nyman, I.B.; Braaen, S.; Dahle, M.K.; Rimstad, E. Viral protein kinetics of piscine orthoreovirus infection in atlantic salmon blood cells. Viruses 2017, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.S.; Bjorgen, H.; Dhamotharan, K.; Wessel, O.; Koppang, E.O.; Di Cicco, E.; Hansen, E.F.; Dahle, M.K.; Rimstad, E. Erythroid progenitor cells in atlantic salmon (salmo salar) may be persistently and productively infected with piscine orthoreovirus (prv). Viruses 2019, 11, 824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polinski, M.P.; Marty, G.D.; Snyman, H.N.; Garver, K.A. Piscine orthoreovirus demonstrates high infectivity but low virulence in atlantic salmon of pacific canada. Sci. Rep. 2019, 9, 3297. [Google Scholar] [CrossRef] [Green Version]
- Di Cicco, E.; Ferguson, H.W.; Kaukinen, K.H.; Schulze, A.D.; Li, S.; Tabata, A.; Günther, O.P.; Mordecai, G.; Suttle, C.A.; Miller, K.M. The same strain of piscine orthoreovirus (prv-1) is involved in the development of different, but related, diseases in atlantic and pacific salmon in british columbia. Facets 2018, 3, 599–641. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, N.; Takahashi, K.; Maita, M.; Rohovec, J.S.; Ikeda, Y. Erythrocytic inclusion body syndrome: Susceptibility of selected sizes of coho salmon and of several other species of salmonid fish. Fish Pathol. 1992, 27, 153–156. [Google Scholar] [CrossRef]
- Hauge, H.; Vendramin, N.; Taksdal, T.; Olsen, A.B.; Wessel, O.; Mikkelsen, S.S.; Alencar, A.L.F.; Olesen, N.J.; Dahle, M.K. Infection experiments with novel piscine orthoreovirus from rainbow trout (oncorhynchus mykiss) in salmonids. PLoS ONE 2017, 12, e0180293. [Google Scholar] [CrossRef] [Green Version]
- Bohle, H.; Bustos, P.; Leiva, L.; Grothusen, H.; Navas, E.; Sandoval, A.; Bustamante, F.; Montecinos, K.; Gaete, A.; Mancilla, M. First complete genome sequence of piscine orthoreovirus variant 3 infecting coho salmon (Oncorhynchus kisutch) farmed in southern chile. Genome Announc. 2018, 6, e00484-18. [Google Scholar] [CrossRef] [Green Version]
- Olsen, A.B.; Hjortaas, M.; Tengs, T.; Hellberg, H.; Johansen, R. First description of a new disease in rainbow trout (oncorhynchus mykiss (walbaum)) similar to heart and skeletal muscle inflammation (hsmi) and detection of a gene sequence related to piscine orthoreovirus (prv). PLoS ONE 2015, 10, e0131638. [Google Scholar] [CrossRef]
- Vendramin, N.; Alencar, A.L.F.; Iburg, T.M.; Dahle, M.K.; Wessel, O.; Olsen, A.B.; Rimstad, E.; Olesen, N.J. Piscine orthoreovirus infection in atlantic salmon (salmo salar) protects against subsequent challenge with infectious hematopoietic necrosis virus (ihnv). Vet. Res. 2018, 49, 30. [Google Scholar] [CrossRef] [Green Version]
- Dhamotharan, K.; Tengs, T.; Wessel, O.; Braaen, S.; Nyman, I.B.; Hansen, E.F.; Christiansen, D.H.; Dahle, M.K.; Rimstad, E.; Markussen, T. Evolution of the piscine orthoreovirus genome linked to emergence of heart and skeletal muscle inflammation in farmed atlantic salmon (salmo salar). Viruses 2019, 11, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiegertjes, G.F.; Wentzel, A.S.; Spaink, H.P.; Elks, P.M.; Fink, I.R. Polarization of immune responses in fish: The ‘macrophages first’ point of view. Mol. Immunol. 2016, 69, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Haatveit, H.M.; Nyman, I.B.; Markussen, T.; Wessel, O.; Dahle, M.K.; Rimstad, E. The non-structural protein muns of piscine orthoreovirus (prv) forms viral factory-like structures. Vet. Res. 2016, 47, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauge, H.; Dahle, M.; Moldal, T.; Thoen, E.; Gjevre, A.G.; Weli, S.; Alarcon, M.; Grove, S. Piscine orthoreovirus can infect and shed through the intestine in experimentally challenged atlantic salmon (Salmo salar L.). Vet. Res. 2016, 47, 57. [Google Scholar] [CrossRef] [PubMed]
- Witeska, M. Erythrocytes in teleost fishes: A review. Zool. Ecol. 2013, 23, 275–281. [Google Scholar] [CrossRef]
- Götting, M.; Nikinmaa, M.J. Transcriptomic analysis of young and old erythrocytes of fish. Front. Physiol. 2017, 8, 1046. [Google Scholar] [CrossRef] [Green Version]
- Garver, K.A.; Johnson, S.C.; Polinski, M.P.; Bradshaw, J.C.; Marty, G.D.; Snyman, H.N.; Morrison, D.B.; Richard, J. Piscine orthoreovirus from western north america is transmissible to atlantic salmon and sockeye salmon but fails to cause heart and skeletal muscle inflammation. PLoS ONE 2016, 11, e0146229. [Google Scholar] [CrossRef] [Green Version]
- Wessel, O.; Olsen, C.M.; Rimstad, E.; Dahle, M.K. Piscine orthoreovirus (prv) replicates in atlantic salmon (Salmo salar L.) erythrocytes ex vivo. Vet. Res. 2015, 46, 26. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, C.K.; Li, Z.; George, C.X.; Samuel, C.E. Protein kinase pkr and rna adenosine deaminase adar1: New roles for old players as modulators of the interferon response. Curr. Opin. Immunol. 2011, 23, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.A.; Schmechel, S.C.; Williams, B.R.G.; Silverman, R.H.; Schiff, L.A. Involvement of the interferon-regulated antiviral proteins PKR and RNASE L in reovirus-induced shutoff of cellular translation. J. Virol. 2005, 79, 2240–2250. [Google Scholar] [CrossRef] [Green Version]
- Lanoie, D.; Boudreault, S.; Bisaillon, M.; Lemay, G. How many mammalian reovirus proteins are involved in the control of the interferon response? Pathogens 2019, 8, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikalsen, A.B.; Haugland, O.; Rode, M.; Solbakk, I.T.; Evensen, O. Atlantic salmon reovirus infection causes a cd8 t cell myocarditis in atlantic salmon (Salmo salar L.). PLoS ONE 2012, 7, e37269. [Google Scholar] [CrossRef] [PubMed]
- Teige, L.H.; Lund, M.; Haatveit, H.M.; Røsæg, M.V.; Wessel, Ø.; Dahle, M.K.; Storset, A.K. A bead based multiplex immunoassay detects piscine orthoreovirus specific antibodies in atlantic salmon (Salmo salar). Fish Shellfish Immunol. 2017, 63, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart regeneration in zebrafish. Science 2002, 298, 2188–2190. [Google Scholar] [CrossRef]
- Lund, M.; Krudtaa Dahle, M.; Timmerhaus, G.; Alarcon, M.; Powell, M.; Aspehaug, V.; Rimstad, E.; Jorgensen, S.M. Hypoxia tolerance and responses to hypoxic stress during heart and skeletal muscle inflammation in atlantic salmon (Salmo salar). PLoS ONE 2017, 12, e0181109. [Google Scholar] [CrossRef]
- Forlenza, M.; Fink, I.R.; Raes, G.; Wiegertjes, G.F. Heterogeneity of macrophage activation in fish. Dev. Comp. Immunol. 2011, 35, 1246–1255. [Google Scholar] [CrossRef]
- Caldwell, R.B.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.W. Arginase: An old enzyme with new tricks. Trends Pharmacol. Sci. 2015, 36, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Wessel, O.; Haugland, O.; Rode, M.; Fredriksen, B.N.; Dahle, M.K.; Rimstad, E. Inactivated piscine orthoreovirus vaccine protects against heart and skeletal muscle inflammation in atlantic salmon. J. Fish Dis. 2018, 41, 1411–1419. [Google Scholar] [CrossRef]
- Wessel, O.; Nyman, I.B.; Markussen, T.; Dahle, M.K.; Rimstad, E. Piscine orthoreovirus (prv) o3 protein binds dsrna. Virus Res. 2015, 198, 22–29. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhamotharan, K.; Bjørgen, H.; Malik, M.S.; Nyman, I.B.; Markussen, T.; Dahle, M.K.; Koppang, E.O.; Wessel, Ø.; Rimstad, E. Dissemination of Piscine orthoreovirus-1 (PRV-1) in Atlantic Salmon (Salmo salar) during the Early and Regenerating Phases of Infection. Pathogens 2020, 9, 143. https://doi.org/10.3390/pathogens9020143
Dhamotharan K, Bjørgen H, Malik MS, Nyman IB, Markussen T, Dahle MK, Koppang EO, Wessel Ø, Rimstad E. Dissemination of Piscine orthoreovirus-1 (PRV-1) in Atlantic Salmon (Salmo salar) during the Early and Regenerating Phases of Infection. Pathogens. 2020; 9(2):143. https://doi.org/10.3390/pathogens9020143
Chicago/Turabian StyleDhamotharan, Kannimuthu, Håvard Bjørgen, Muhammad Salman Malik, Ingvild B. Nyman, Turhan Markussen, Maria K. Dahle, Erling Olaf Koppang, Øystein Wessel, and Espen Rimstad. 2020. "Dissemination of Piscine orthoreovirus-1 (PRV-1) in Atlantic Salmon (Salmo salar) during the Early and Regenerating Phases of Infection" Pathogens 9, no. 2: 143. https://doi.org/10.3390/pathogens9020143