Novel Production of Bovine Papillomavirus Pseudovirions in Tobacco Plants




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
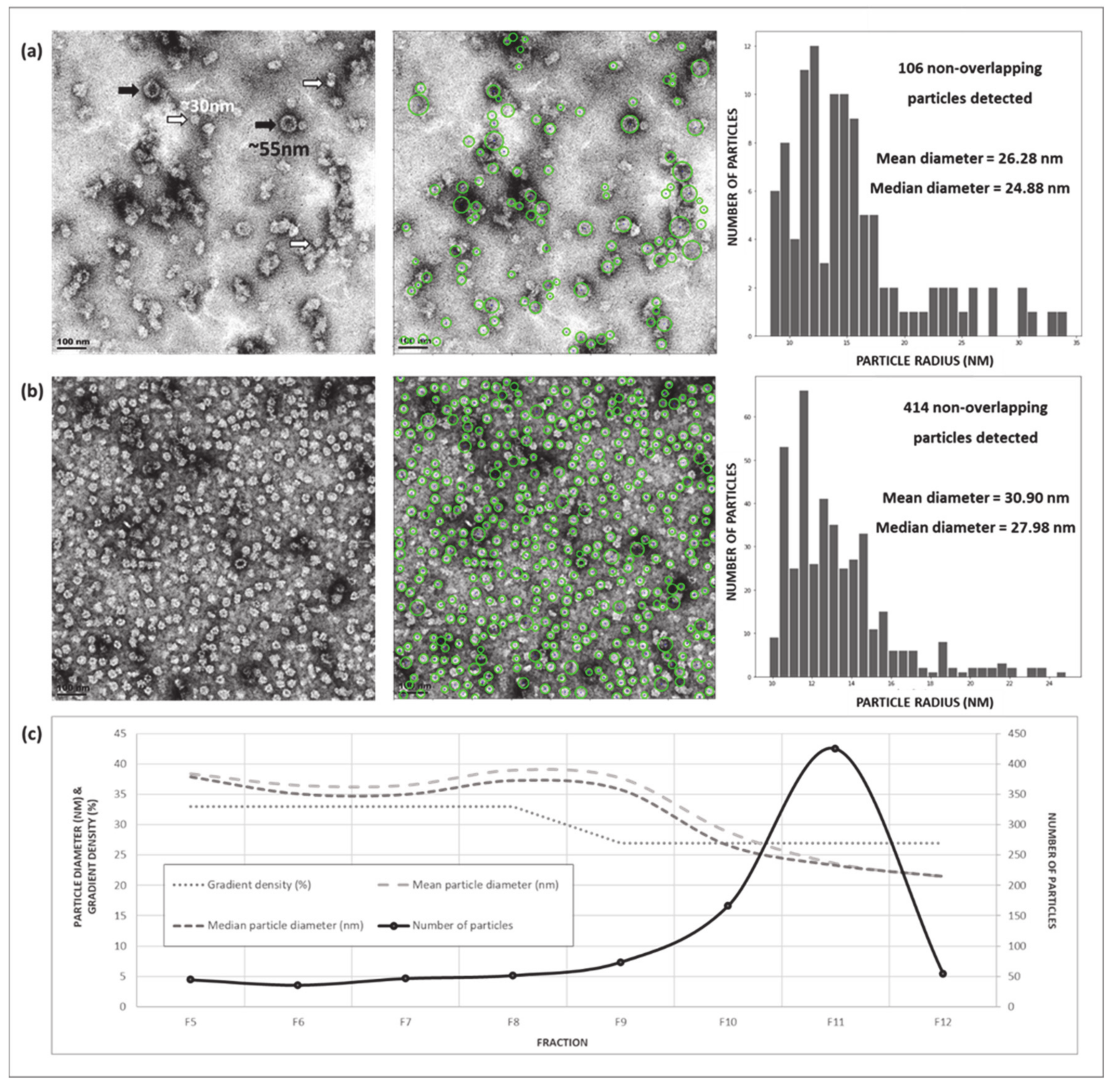
2. Results
2.1. Transient Expression of BPV1 VLPs in N. benthamiana
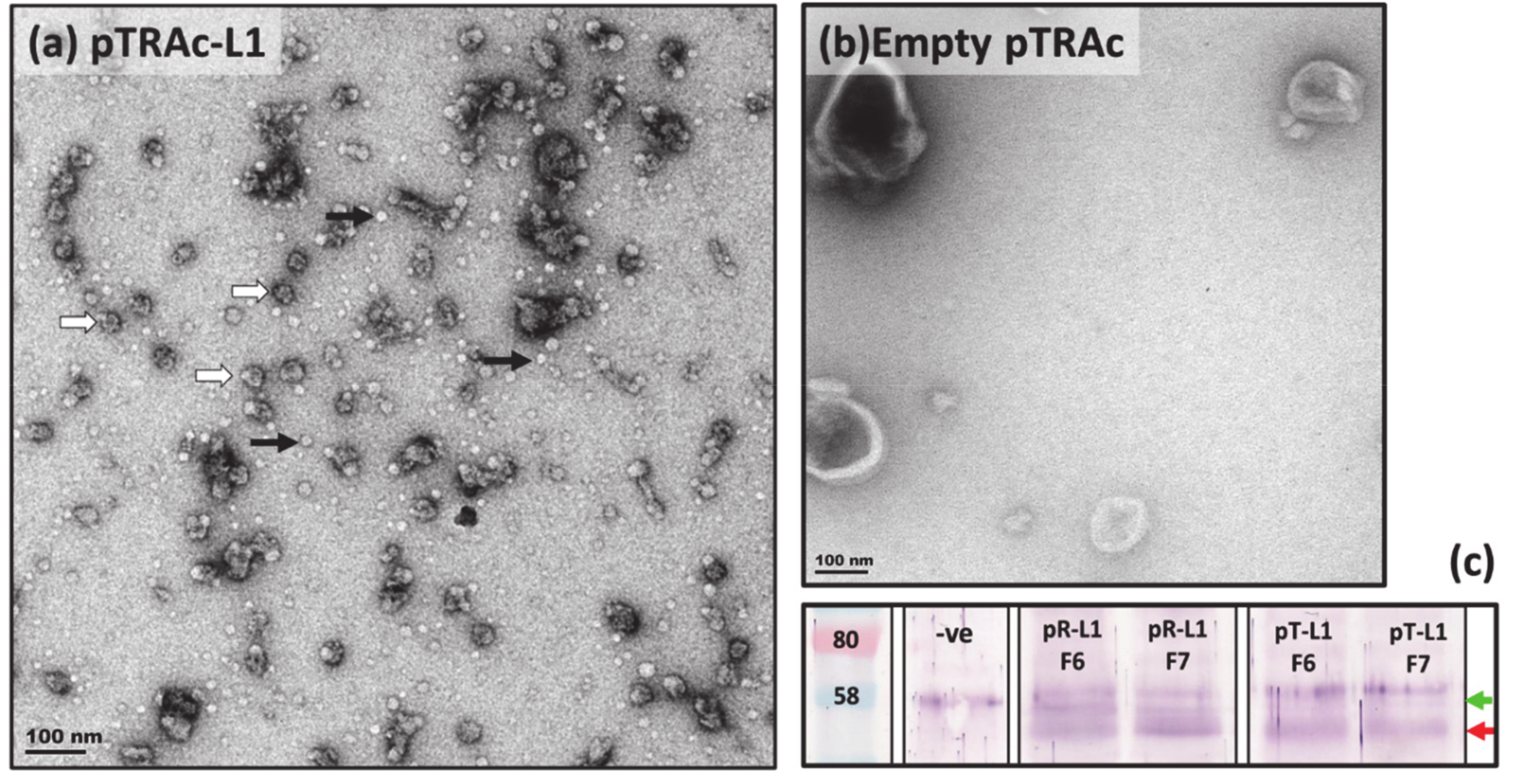
2.2. Expression and Purification of Plant-Produced BPV1 PsVs
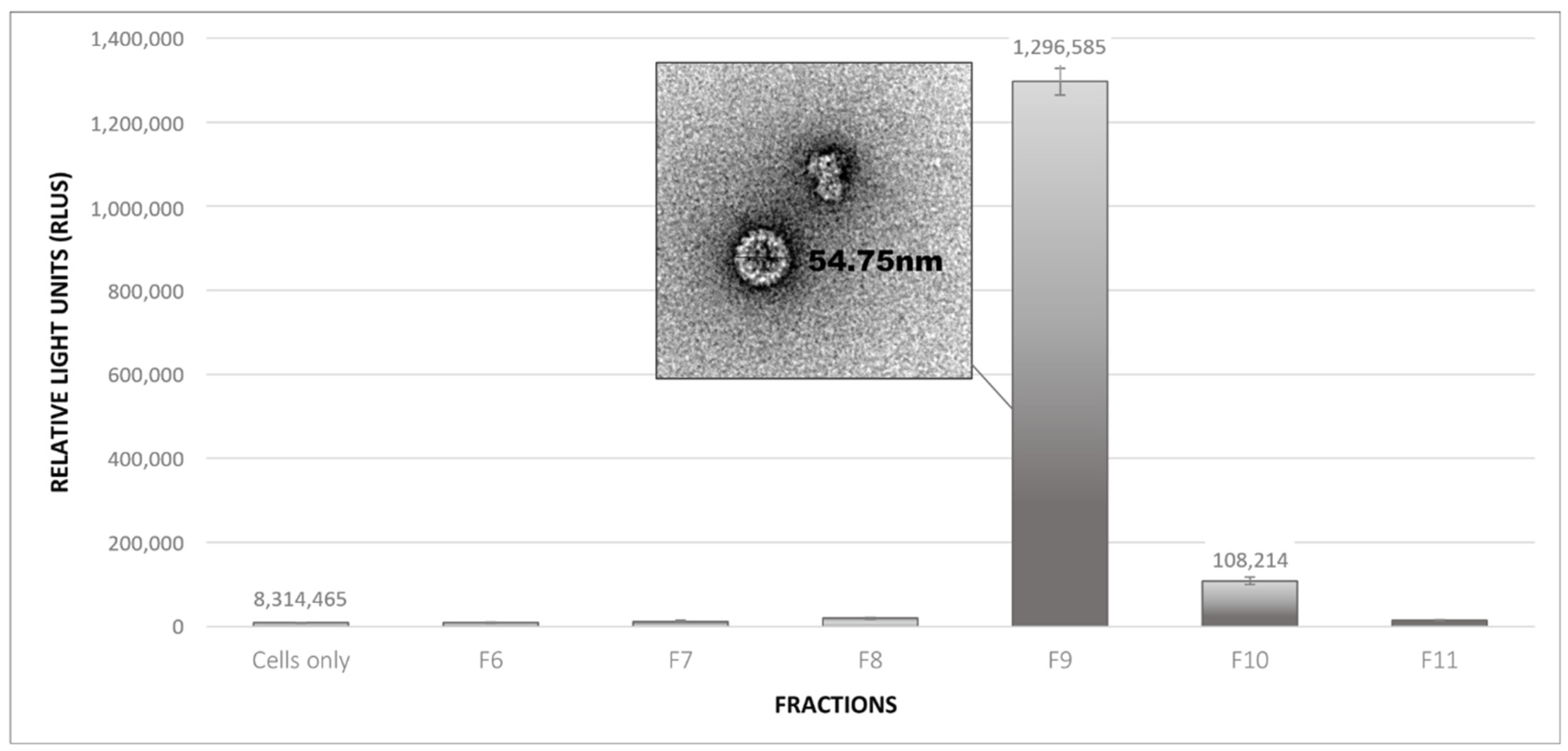
2.3. Infectivity Assays of Plant-Produced BPV PsVs
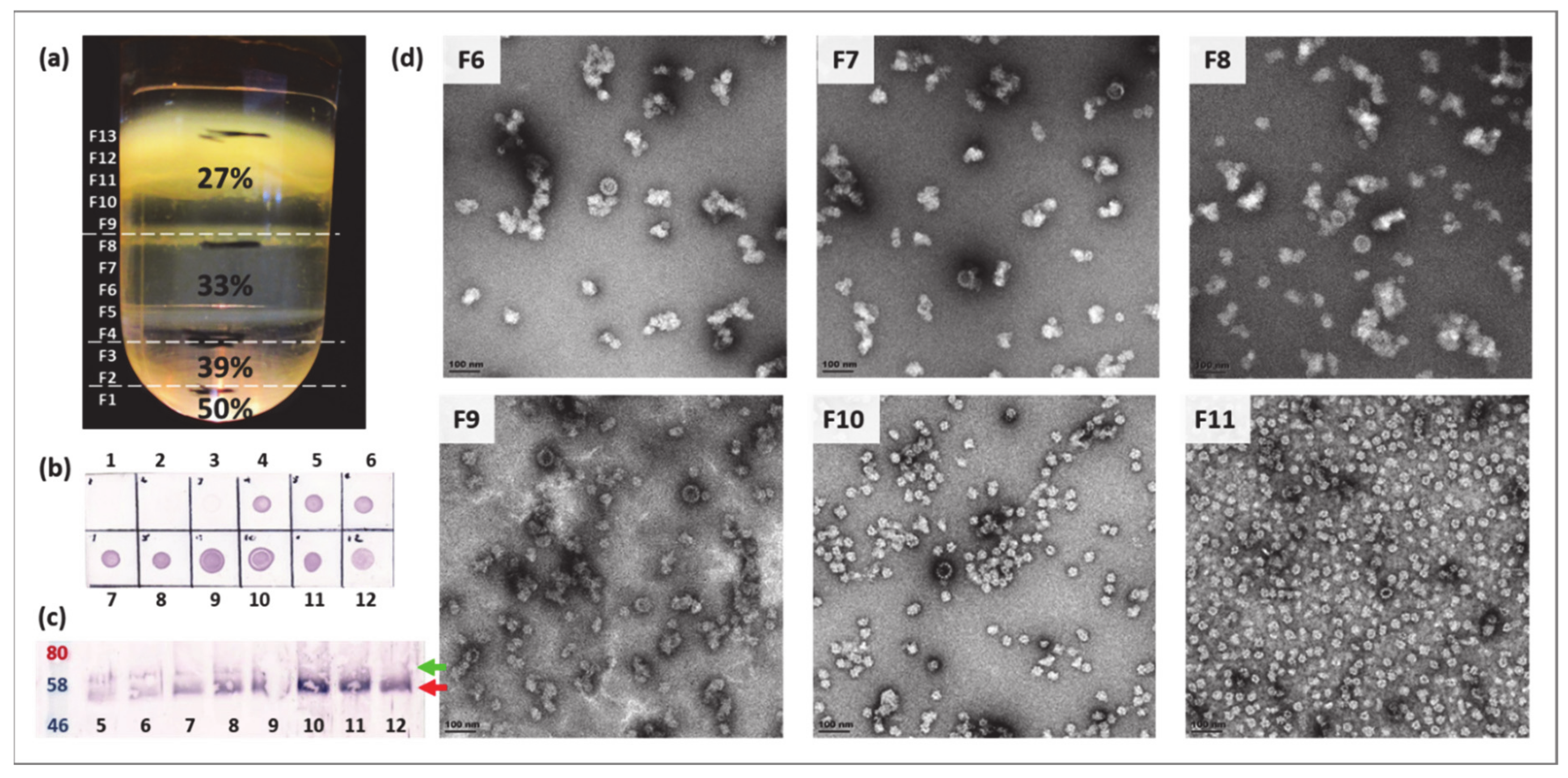
2.4. Purification and Expression Optimisation of BPV VLPs and PsVs
2.4.1. Effect of Biomass Freezing on Particle Stability
2.4.2. Optimisation of Density Gradient Ultracentrifugation
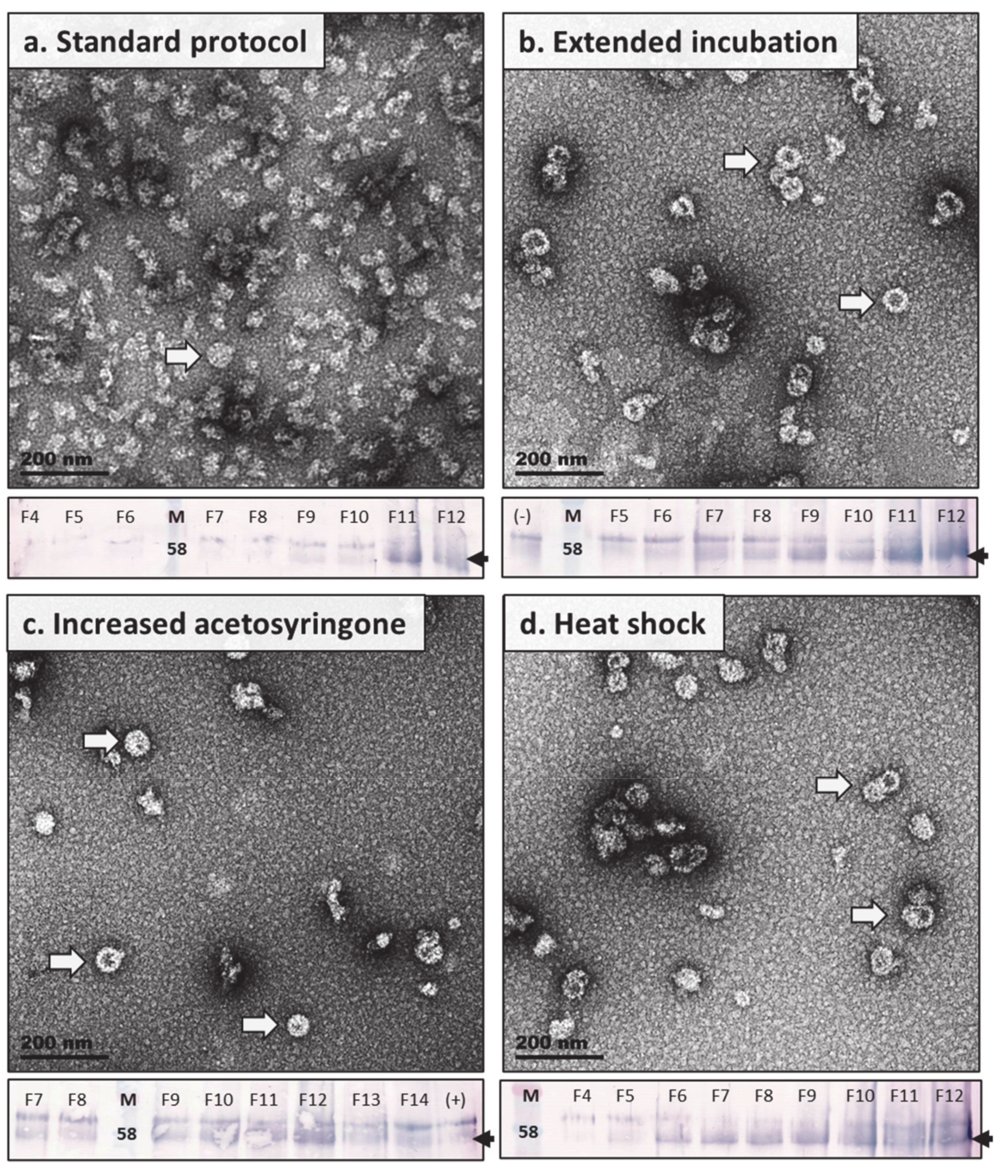
2.4.3. Expression Studies: Heat Shock Treatment, Increased Acetosyringone, and Extended in Planta Maturation
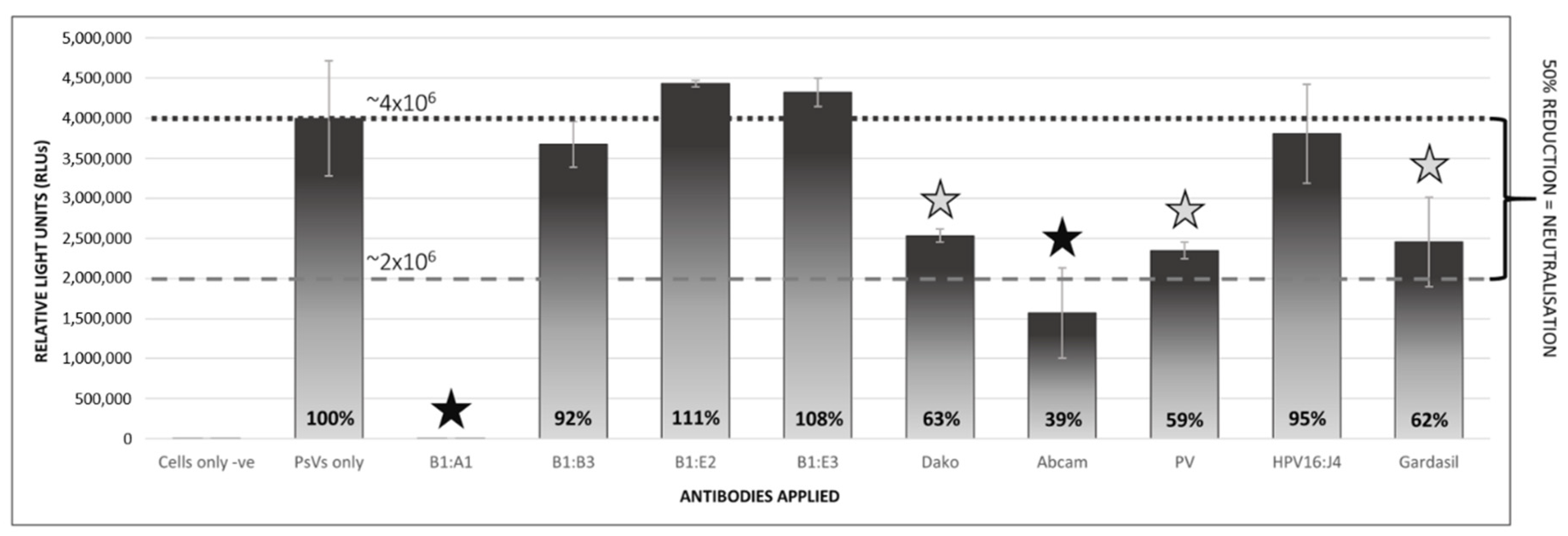
2.5. Pseudovirion-Based Neutralisation Assay of Plant-Produced BPV1 PsVs
3. Discussion
4. Limitations and Future Work
5. Conclusions
6. Materials and Methods
6.1. Construct Design and Generation of Recombinant Agrobacterium
6.2. Transient Expression of BPV1 VLPs in N. benthamiana
6.3. Transient Expression of BPV1 PsVs in N. benthamiana
6.4. Large-Scale Protein Purification of BPV1 VLPs and PsVs
6.5. SDS-PAGE and Western Blots
6.6. Particle Visualisation and Analysis
6.7. Optimisation of Protein Expression and Purification
6.8. Growth and Maintenance of HEK293TT Cell Culture
6.9. SEAP Assay for PsV Infection of Mammalian Cells
6.10. Neutralisation of Plant-Produced BPV1 Pseudovirions by BPV, HPV, and PV Antibodies
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campo, M.S.; Roden, R.B.S. Papillomavirus Prophylactic Vaccines: Established Successes, New Approaches. J. Virol. 2010, 84, 1214–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araldi, R.P.; Muro, S.; Assaf, R.; Carvalho, R.F.D.; Caldas, M.A.; Carvalho, R.D.; Souza, J.M.D.; Magnelli, R.F.; Grando, D.; Roperto, F.P.; et al. Papillomaviruses: A systematic review. Genet. Mol. Biol. 2017, 40, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocaneti, F.; Altamura, G.; Corteggio, A.; Velescu, E.; Roperto, F.; Borzacchiello, G. Bovine Papillomavirus: New Insights into an Old Disease. Transbound. Emerg. Dis. 2016, 63, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Savini, F.; Gallina, L.; Prosperi, A.; Puleio, R.; Lavazza, A.; Marco, P.D.; Tumino, S.; Moreno, A.; Lelli, D.; Guercio, A.; et al. Bovine Papillomavirus 1 Gets Out of the Flock: Detection in an Ovine Wart in Sicily. Pathogens 2020, 9, 429. [Google Scholar] [CrossRef]
- Roperto, S.; Russo, V.; Corrado, F.; De Falco, F.; Munday, J.S.; Roperto, F. Oral fibropapillomatosis and epidermal hyperplasia of the lip in newborn lambs associated with bovine Deltapapillomavirus. Sci. Rep. 2018, 8, 13310. [Google Scholar] [CrossRef]
- Munday, J.S. Papillomaviruses in felids. Vet. J. 2014, 199, 340–347. [Google Scholar] [CrossRef]
- Orbell, G.M.B.; Young, S.; Munday, J.S. Cutaneous Sarcoids in Captive African Lions Associated With Feline Sarcoid-Associated Papillomavirus Infection. Vet. Pathol. 2011, 48, 1176–1179. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.H.; van Dyk, E.; Nel, P.J.; Lane, E.; Van Wilpe, E.; Bengis, R.G.; de Klerk-Lorist, L.M.; van Heerden, J. Pathology and immunohistochemistry of papillomavirus-associated cutaneous lesions in Cape mountain zebra, giraffe, sable antelope and African buffalo in South Africa. J. S. Afr. Vet. Assoc. 2011, 82, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Van Dyk, E.; Bosman, A.-M.; Van Wilpe, E.; Williams, J.H.; Bengis, R.G.; Van Heerden, J.; Venter, E.H. Detection and characterisation of papillomavirus in skin lesions of giraffe and sable antelope in South Africa. J. S. Afr. Vet. Assoc. 2011, 82, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Silvestre, O.; Orzacchiello, G.B.; Ava, D.N.; Ovane, G.I.; Usso, V.R.; Ecchio, D.V.; Usilio, F.D.A.; Ault, E.A.G.; Ampo, M.S.C.; Aciello, O.P. Bovine Papillomavirus Type 1 DNA and E5 Oncoprotein Expression in Water Buffalo Fibropapillomas. Vet. Pathol. 2009, 46, 636–641. [Google Scholar] [CrossRef]
- Roperto, S.; Russo, V.; Ozkul, A.; Sepici-dincel, A.; Maiolino, P.; Borzacchiello, G.; Marcus, I.; Esposito, I.; Riccardi, M.G.; Roperto, F. Communication Bovine papillomavirus type 2 infects the urinary bladder of water buffalo (Bubalus bubalis) and plays a crucial role in bubaline urothelial carcinogenesis. J. Gen. Virol. 2013, 94, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Pangty, K.; Singh, S.; Goswami, R.; Saikumar, G.; Somvanshi, R. Detection of BPV-1 and -2 and Quantification of BPV-1 by Real-Time PCR in Cutaneous Warts in Cattle and Buffaloes. Transbound. Emerg. Dis. 2010, 57, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Campo, M.S. Animal models of papillomavirus pathogenesis. Virus Res. 2002, 89, 249–261. [Google Scholar] [CrossRef]
- Taylor, S.; Haldorson, G. A review of equine sarcoid. Equine Vet. Educ. 2013, 25, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Lunardi, M.; Alcântara, K.D.; Arellano, A.; Rodrigues, B. Bovine Papillomavirus Type 13 DNA in Equine Sarcoids. J. Clin. Microbiol. 2013, 51, 2167–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pangty, K.; Singh, S.; Pandey, A.B.; Somvanshi, R. Preliminary binary ethylenimine (BEI) inactivated bovine papillomavirus (BPV) vaccine trial against cutaneous warts in bull calves : A pathological assessment. Braz. J. Vet. Pathol. 2010, 3, 105–110. [Google Scholar]
- Terziev, G.; Roydev, R.; Kalkanov, I.; Borissov, I.; Dinev, I. Papillomatosis in heifers—Comparative studies on Surgical excision and autogenous vaccine therapies. Trakia J. Sci. 2015, 13, 274–279. [Google Scholar] [CrossRef]
- Kale, M.; Saltik, H.S.; Hasircioglu, S.; Yildirim, Y.; Yavru, S.; Mamak, N.; Atli, K. Treatment of Bovine papillomavirus-induced teat warts in a cow by using Podophyllin magistral formula and autologous vaccine applications together. Indian J. Anim. Res. 2019, 832–836. [Google Scholar] [CrossRef] [Green Version]
- Rothacker, C.C.; Boyle, A.G.; Levine, D.G. Autologous vaccination for the treatment of equine sarcoids: 18 cases (2009–2014). Can. Vet. J. 2014, 56, 709–714. [Google Scholar]
- Liu, F.; Ge, S.; Li, L.; Wu, X.; Liu, Z.; Wang, Z. Virus-like particles: Potential veterinary vaccine immunogens. Res. Vet. Sci. 2012, 93, 553–559. [Google Scholar] [CrossRef]
- Liu, F.; Wu, X.; Li, L.; Ge, S.; Liu, Z.; Wang, Z. Virus-like particles: Promising platforms with characteristics of DIVA for veterinary vaccine design. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Kirnbauer, R.; Chandrachud, L.M.; O’Neil, B.W.; Wagner, E.R.; Grindlay, G.J.; Armstrong, A.; McGarvie, G.M.; Schiller, J.T.; Lowy, D.R.; Campo, M.S. Virus-like particles of bovine papillomavirus type 4 in prophylactic and therapeutic immunization. Virology 1996, 219, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campo, M.S. Vaccination against papillomavirus in cattle. Clin. Dermatol. 1997, 15, 275–283. [Google Scholar] [CrossRef]
- Hartl, B.; Hainisch, E.K.; Shafti-Keramat, S.; Kirnbauer, R.; Corteggio, A.; Borzacchiello, G.; Tober, R.; Kainzbauer, C.; Pratscher, B.; Brandt, S. Inoculation of young horses with bovine papillomavirus type 1 virions leads to early infection of PBMCS prior to pseudo-sarcoid formation. J. Gen. Virol. 2011, 92, 2437–2445. [Google Scholar] [CrossRef]
- Kirnbauer, R.; Booy, F.; Cheng, N.; Lowy, D.R.; Schiller, J.T. Papillomavirus L1 major capsid protein self-assembles into virus-like particles that are highly immunogenic. Proc. Natl. Acad. Sci. USA 1992, 89, 12180–12184. [Google Scholar] [CrossRef] [Green Version]
- Shafti-Keramat, S.; Schellenbacher, C.; Handisurya, A.; Christensen, N.; Reininger, B.; Brandt, S.; Kirnbauer, R. Bovine papillomavirus type 1 (BPV1) and BPV2 are closely related serotypes. Virology 2009, 393, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Gaukroger, J.M.; Chandrachud, L.M.; O’Neil, B.W.; Grindlay, G.J.; Knowles, G.; Campo, M.S. Vaccination of cattle with bovine papillomavirus type 4 L2 elicits the production of virus-neutralizing antibodies. J. Gen. Virol. 1996, 77, 1577–1583. [Google Scholar] [CrossRef]
- Mariz, F.C.; Jesus, A.L.S.; Silva, M.A.R. The Challenges Inherent in the Control and Prevention of Bovine Papillomaviruses. Austin J. Genet. Genom. Res. 2016, 3, 1017. [Google Scholar]
- Bayer, L.; Gumpel, J.; Hause, G.; Muller, M.; Grunwald, T. Non-human papillomaviruses for gene delivery in vitro and in vivo. PLoS ONE 2018, 13, e0198996. [Google Scholar] [CrossRef]
- Kines, R.C.; Zarnitsyn, V.; Johnson, T.R.; Pang, Y.S.; Corbett, K.S.; Nicewonger, J.D.; Gangopadhyay, A.; Chen, M.; Liu, J. Vaccination with Human Papillomavirus Pseudovirus-Encapsidated Plasmids Targeted to Skin Using Microneedles. PLoS ONE 2015, 10, e0120797. [Google Scholar] [CrossRef]
- Cerqueira, C.; Thompson, C.D.; Day, P.M.; Pang, Y.-Y.S.; Lowy, D.R.; Schiller, J.T. Efficient Production of Papillomavirus Gene Delivery Vectors in Defined In Vitro Reactions. Mol. Ther. Methods Clin. Dev. 2017, 5, 165–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, C.B.; Pastrana, D.V.; Lowy, D.R.; Schiller, J.T.; Al, B.E.T. Efficient Intracellular Assembly of Papillomaviral Vectors. J. Virol. 2004, 78, 751–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, C.B.; Pastrana, D.V.; Lowy, D.R.; Schiller, J.T. Generation of HPV pseudovirions using transfection and their use in neutralization assays. Methods Mol. Med. 2005, 119, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Pastrana, D.V.; Buck, C.B.; Pang, Y.Y.S.; Thompson, C.D.; Castle, P.E.; FitzGerald, P.C.; Kjaer, S.K.; Lowy, D.R.; Schiller, J.T. Reactivity of human sera in a sensitive, high-throughput pseudovirus-based papillomavirus neutralization assay for HPV16 and HPV18. Virology 2004, 321, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Lin, Z.J.; Huang, S.J.; Li, J.; Liu, X.H.; Guo, M.; Zhang, J.; Xia, N.S.; Pan, H.R.; Wu, T.; et al. Correlation between ELISA and pseudovirion-based neutralisation assay for detecting antibodies against human papillomavirus acquired by natural infection or by vaccination. Hum. Vaccines Immunother. 2014, 10, 740–746. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.F.; Brownlow, M.; Brown, M.; Kowalski, R.; Esser, M.T.; Ruiz, W.; Barr, E.; Brown, D.R.; Bryan, J.T. Antibodies from women immunized with Gardasil® cross-neutralize HPV 45 pseudovirions. Hum. Vaccin. 2007, 3, 109–115. [Google Scholar] [CrossRef]
- Sehr, P.; Rubio, I.; Seitz, H.; Putzker, K.; Ribeiro-Müller, L.; Pawlita, M.; Müller, M. High-Throughput Pseudovirion-Based Neutralization Assay for Analysis of Natural and Vaccine-Induced Antibodies against Human Papillomaviruses. PLoS ONE 2013, 8, e75677. [Google Scholar] [CrossRef]
- Leung, T.F.; Liu, A.P.Y.; Lim, F.S.; Thollot, F.; Oh, H.M.L.; Lee, B.W.; Rombo, L.; Tan, N.C.; Rouzier, R.; De Simoni, S.; et al. Comparative immunogenicity and safety of human papillomavirus (HPV)-16/18 AS04-adjuvanted vaccine and 4vHPV vaccine administered according to two- or three-dose schedules in girls aged 9–14 years: Results to month 36 from a randomized trial. Vaccine 2018, 36, 98–106. [Google Scholar] [CrossRef]
- Yeager, M.D.; Aste-Amezaga, M.; Brown, D.R.; Martin, M.M.; Shah, M.J.; Cook, J.C.; Christensen, N.D.; Ackerson, C.; Lowe, R.S.; Smith, J.F.; et al. Neutralization of human papillomavirus (HPV) pseudovirions: A novel and efficient approach to detect and characterize HPV neutralizing antibodies. Virology 2000, 278, 570–577. [Google Scholar] [CrossRef] [Green Version]
- Sankaranarayanan, R.; Prabhu, P.R.; Pawlita, M.; Gheit, T.; Bhatla, N.; Muwonge, R.; Nene, B.M.; Esmy, P.O.; Joshi, S.; Poli, U.R.R.; et al. Immunogenicity and HPV infection after one, two, and three doses of quadrivalent HPV vaccine in girls in India: A multicentre prospective cohort study. Lancet Oncol. 2016, 17, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Dessy, F.J.; Giannini, S.L.; Bougelet, C.A.; Kemp, T.J.; David, M.P.M.; Poncelet, S.M.; Pinto, L.A.; Wettendorff, M.A. Correlation between direct ELISA, single epitope-based inhibition ELISA and pseudovirion-based neutralization assay for measuring anti-HPV-16 and anti-HPV-18 antibody response after vaccination with the AS04-adjuvanted HPV-16/18 cervical cancer vaccine. Hum. Vaccin. 2008, 4, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Wang, Y.; Chen, N.; Jiang, D.; Qiu, Y.; Wang, Y.; Yan, M.; Chen, J.; Zhang, H.; Liu, Y. A novel trivalent HPV 16/18/58 vaccine with anti-HPV 16 and 18 neutralizing antibody responses comparable to those induced by the Gardasil quadrivalent vaccine in rhesus macaque model. Papillomavirus Res. 2017, 3, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Folschweiller, N.; Teixeira, J.; Joshi, S.; Goldani, L.Z.; Supparatpinyo, K.; Basu, P.; Chotpitayasunondh, T.; Chetchotisakd, P.; Ruxrungtham, K.; Roteli-Martins, C.; et al. Immunogenicity and safety of the AS04-HPV-16/18 and HPV-6/11/16/18 human papillomavirus vaccines in asymptomatic young women living with HIV aged 15–25 years: A phase IV randomized comparative study. EClinicalMedicine 2020, 23, 100353. [Google Scholar] [CrossRef] [PubMed]
- Hainisch, E.K.; Abel-reichwald, H.; Shafti-keramat, S.; Pratscher, B.; Corteggio, A.; Borzacchiello, G.; Wetzig, M.; Jindra, C.; Tichy, A.; Kirnbauer, R.; et al. Potential of a BPV1 L1 VLP vaccine to prevent BPV1- or BPV2- induced pseudo-sarcoid formation and safety and immunogenicity of EcPV2 L1 VLPs in horse. J. Gen. Virol. 2017, 98, 230–241. [Google Scholar] [CrossRef]
- Dubey, K.K.; Luke, G.A.; Knox, C.; Kumar, P.; Pletschke, B.I.; Singh, P.K.; Shukla, P. Vaccine and antibody production in plants : Developments and computational tools. Brief. Funct. Genom. 2018, 17, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Rybicki, E.P. Plant-based vaccines against viruses. Virol. J. 2014, 11, 205. [Google Scholar] [CrossRef]
- Love, A.J.; Chapman, S.N.; Matic, S.; Noris, E.; Lomonossoff, G.P.; Taliansky, M. In planta production of a candidate vaccine against bovine papillomavirus type 1. Planta 2012, 236, 1305–1313. [Google Scholar] [CrossRef]
- Zahin, M.; Joh, J.; Khanal, S.; Husk, A.; Mason, H.; Warzecha, H.; Ghim, S.J.; Miller, D.M.; Matoba, N.; Jenson, A.B. Scalable production of HPV16 L1 protein and VLPs from tobacco leaves. PLoS ONE 2016, 11, e0160995. [Google Scholar] [CrossRef] [Green Version]
- Thuenemann, E.C.; Lenzi, P.; Love, A.J.; Taliansky, M.; Bécares, M.; Zuñiga, S.; Enjuanes, L.; Zahmanova, G.G.; Minkov, I.N.; Noris, E.; et al. The Use of Transient Expression Systems for the Rapid Production of Virus-like Par- ticles in Plants. Curr. Pharm. Des. 2013, 19, 5564–5573. [Google Scholar] [CrossRef]
- Lamprecht, R.L.; Kennedy, P.; Huddy, S.M.; Bethke, S.; Hendrikse, M.; Hitzeroth, I.I.; Rybicki, E.P. Production of Human papillomavirus pseudovirions in plants and their use in pseudovirion-based neutralisation assays in mammalian cells. Nat. Sci. Rep. 2016, 6, 20431. [Google Scholar] [CrossRef] [Green Version]
- Matić, S.; Masenga, V.; Poli, A.; Rinaldi, R.; Milne, R.G.; Vecchiati, M.; Noris, E. Comparative analysis of recombinant Human Papillomavirus 8L1 production in plants by a variety of expression systems and purification methods. Plant Biotechnol. J. 2012, 10, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Baker, T.S.; Newcomb, W.W.; Olson, N.H.; Cowsert, L.M.; Olson, C.; Brown, J.C. Structures of bovine and human papillomaviruses. Analysis by cryoelectron microscopy and three-dimensional image reconstruction. Biophys. J. 1991, 60, 1445–1456. [Google Scholar] [CrossRef] [Green Version]
- Belnap, D.M.; Olson, N.H.; Cladel, N.M.; Newcomb, W.W.; Brown, J.C.; Kreider, J.W.; Christensen, N.D.; Baker, T.S. Conserved features in papillomavirus and polyomavirus capsids. J. Mol. Biol. 1996, 259, 249–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broniarczyk, J.; Massimi, P.; Pim, D.; Marušic, B.; Myers, M.P.; Garcea, R.L.; Banks, L. Phosphorylation of human papillomavirus type 16 L2 contributes to efficient virus infectious entry. J. Virol. 2019, 93, e00128-19. [Google Scholar] [CrossRef] [Green Version]
- Sweke, R. Particle Counter, GitHub, 2017. Available online: https://github.com/CorrieGunter/particle_counter (accessed on 26 November 2020).
- Norkunas, K.; Harding, R.; Dale, J.; Dugdale, B. Improving agroinfiltration—Based transient gene expression in Nicotiana benthamiana. Plant Methods 2018, 14, 71. [Google Scholar] [CrossRef]
- Schiller, J.T.; Lowy, D.R. Immunogenicity Testing in Human Papillomavirus Virus-Like-Particle Vaccine Trials. J. Infect. Dis. 2009, 200, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Fleury, M.J.J.; Touzé, A.; de Sanjosé, S.; Bosch, F.X.; Klaustermeiyer, J.; Coursaget, P. Detection of Human Papillomavirus Type 31-Neutralizing Antibodies from Naturally Infected Patients by an Assay Based on Intracellular Assembly of Luciferase-Expressing Pseudovirions. Clin. Vaccine Immunol. 2008, 15, 172. [Google Scholar] [CrossRef] [Green Version]
- Jiang, R.T.; Schellenbacher, C.; Chackerian, B.; Roden, R.B.S. Progress and prospects for L2-based human papillomavirus vaccines. Expert Rev. Vaccines 2016, 15, 853–862. [Google Scholar] [CrossRef] [Green Version]
- Roden, R.B.S.; Day, P.M.; Bronzo, B.K.; Iv, W.H.Y.; Yang, Y.; Lowy, D.R.; Schiller, J.T. Positively Charged Termini of the L2 Minor Capsid Protein Are Necessary for Papillomavirus Infection. J. Virol. 2001, 75, 10493–10497. [Google Scholar] [CrossRef] [Green Version]
- Jarrett, W.F.H.; Smith, K.T.; Neil, B.W.O.; Gaukroger, M.; Chandrachud, L.M.; Al, J.E.T. Studies on Vaccination against Papillomaviruses: Prophylactic and Therapeutic Vaccination with Recombinant Structural Proteins. Virology 1991, 184, 33–42. [Google Scholar] [CrossRef]
- Buck, C.B.; Thompson, C.D.; Pang, Y.-Y.S.; Lowy, D.R.; Schiller, J.T. Maturation of Papillomavirus Capsids. J. Virol. 2005, 79, 2839–2846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, R.; Vasilev, N.; Twyman, R.M.; Schillberg, S. High-value products from plants: The challenges of process optimization. Curr. Opin. Biotechnol. 2015, 32, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Casini, G.L.; Graham, D.; Heine, D.; Garcea, R.L.; Wu, D.T. In vitro papillomavirus capsid assembly analyzed by light scattering. Virology 2004, 325, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Adams, A. Optimization and Characterisation of Plant Produced Human Papillomavirus Pseudovirions in Nicotiana Benthamiana. Master’s Thesis, University of Cape Town, Cape Town, South Africa, 2020. [Google Scholar]
- Zhao, K.-N.; Sun, X.-Y.; Frazer, I.H.; Zhou, J. DNA Packaging by L1 and L2 Capsid Proteins of Bovine Papillomavirus Type 1. Virology 1998, 243, 482–491. [Google Scholar] [CrossRef]
- Zhao, K.-N.; Hengst, K.; Liu, W.-J.; Liu, Y.H.; Liu, X.S.; McMillan, N.A.J.; Frazer, I.H. BPV1 E2 Protein Enhances Packaging of Full-Length Plasmid DNA in BPV1 Pseudovirions. Virology 2000, 272, 382–393. [Google Scholar] [CrossRef] [Green Version]
- Christensen, N.D.; Kreider, J.W. Monoclonal antibody neutralization of BPV-1. Virus Res. 1993, 28, 195–202. [Google Scholar] [CrossRef]
- Maclean, J.; Koekemoer, M.; Olivier, A.J.; Stewart, D.; Hitzeroth, I.I.; Rademacher, T.; Fischer, R.; Williamson, A.-L.; Rybicki, E.P. Optimization of human papillomavirus type 16 (HPV-16) L1 expression in plants: Comparison of the suitability of different HPV-16 L1 gene variants and different cell-compartment localization. J. Gen. Virol. 2007, 88, 1460–1469. [Google Scholar] [CrossRef]
- Shah, K.H.; Almaghrabi, B.; Bohlmann, H. Comparison of Expression Vectors for Transient Expression of Recombinant Proteins in Plants. Plant Mol. Biol. Report. 2013, 31, 1529–1538. [Google Scholar] [CrossRef] [Green Version]
- Reis, R.S.; Litholdo, C.G.; Bally, J.; Roberts, T.H.; Waterhouse, P.M. A conditional silencing suppression system for transient expression. Sci. Rep. 2018, 8, 9426. [Google Scholar] [CrossRef]
- Yanez, R.J.R.; Lamprecht, R.; Granadillo, M.; Torrens, I.; Rybicki, E.P.; Hitzeroth, I.I.; Biology, C.; Town, C.; Africa, S.; Biology, C.; et al. LALF 32-51-E7, a HPV-16 therapeutic vaccine candidate, forms protein body-like structures when expressed in Nicotiana benthamiana leaves. Plant Biotechnol. J. 2018, 16, 628–637. [Google Scholar] [CrossRef] [Green Version]
- Soboleski, M.R.; Oaks, J.; Halford, W.P. Green fluorescent protein is a quantitative reporter of gene expression in individual eukaryotic cells. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2005, 19, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Y.; Chye, M.-L. Use of GFP to investigate expression of plant-derived vaccines. Methods Mol. Biol. 2009, 515, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Leffel, S.M.; Mabon, S.A.; Stewart, C.N. Applications of Green Fluorescent Protein in Plants. BioTechniques 1997, 23, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, E.P. Plant-made vaccines for humans and animals. Plant Biotechnol. J. 2010, 8, 620–637. [Google Scholar] [CrossRef] [PubMed]
- NIAID Office of Cyber Infrastructure and Computational Biology. PaVE: Papilloma Virus Genome Database. 1999. Available online: http://pave.niaid.nih.gov (accessed on 26 November 2020).
- Regnard, G.L.; Halley-Stott, R.P.; Tanzer, F.L.; Hitzeroth, I.I.; Rybicki, E.P. High level protein expression in plants through the use of a novel autonomously replicating geminivirus shuttle vector: Protein expression in plants utilizing BeYDV. Plant Biotechnol. J. 2010, 8, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Christensen, N.D.; Dillner, J.; Eklund, C.; Carter, J.J.; Wipf, G.C.; Reed, C.A.; Cladel, N.M.; Galloway, D.A. Surface Conformational and Linear Epitopes on HPV-16 and HPV-18 L1 Virus-like Particles as Defined by Monoclonal Antibodies. Virology 1996, 184, 174–184. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietersen, I.; van Zyl, A.; Rybicki, E.; Hitzeroth, I. Novel Production of Bovine Papillomavirus Pseudovirions in Tobacco Plants. Pathogens 2020, 9, 996. https://doi.org/10.3390/pathogens9120996
Pietersen I, van Zyl A, Rybicki E, Hitzeroth I. Novel Production of Bovine Papillomavirus Pseudovirions in Tobacco Plants. Pathogens. 2020; 9(12):996. https://doi.org/10.3390/pathogens9120996
Chicago/Turabian StylePietersen, Inge, Albertha van Zyl, Edward Rybicki, and Inga Hitzeroth. 2020. "Novel Production of Bovine Papillomavirus Pseudovirions in Tobacco Plants" Pathogens 9, no. 12: 996. https://doi.org/10.3390/pathogens9120996