Biological and Genomic Characterization of a Novel Jumbo Bacteriophage, vB_VhaM_pir03 with Broad Host Lytic Activity against Vibrio harveyi




Abstract
:1. Introduction
2. Results
2.1. Isolation and Morphology of vB_VhaM_pir03
2.2. Host Range and Efficiency of Plating (EOP) of vB_VhaM_pir03 against Multiple Antibiotic Resistant Strains
2.3. Adsorption Time and One-Step Growth of vB_VhaM_pir03
2.4. In Vitro Cell Lysis
2.5. Whole Genome Sequencing and Assembly
2.6. Genomic Features of vB_VhaM_pir03
2.7. Genomic Arrangement and Functional Annotations of vB_VhaM_pir03
2.7.1. Phage Structural Proteins
2.7.2. DNA Replication, Repair, and Recombination
2.7.3. Nucleotide Metabolism and Transcription
2.7.4. Miscellaneous Proteins
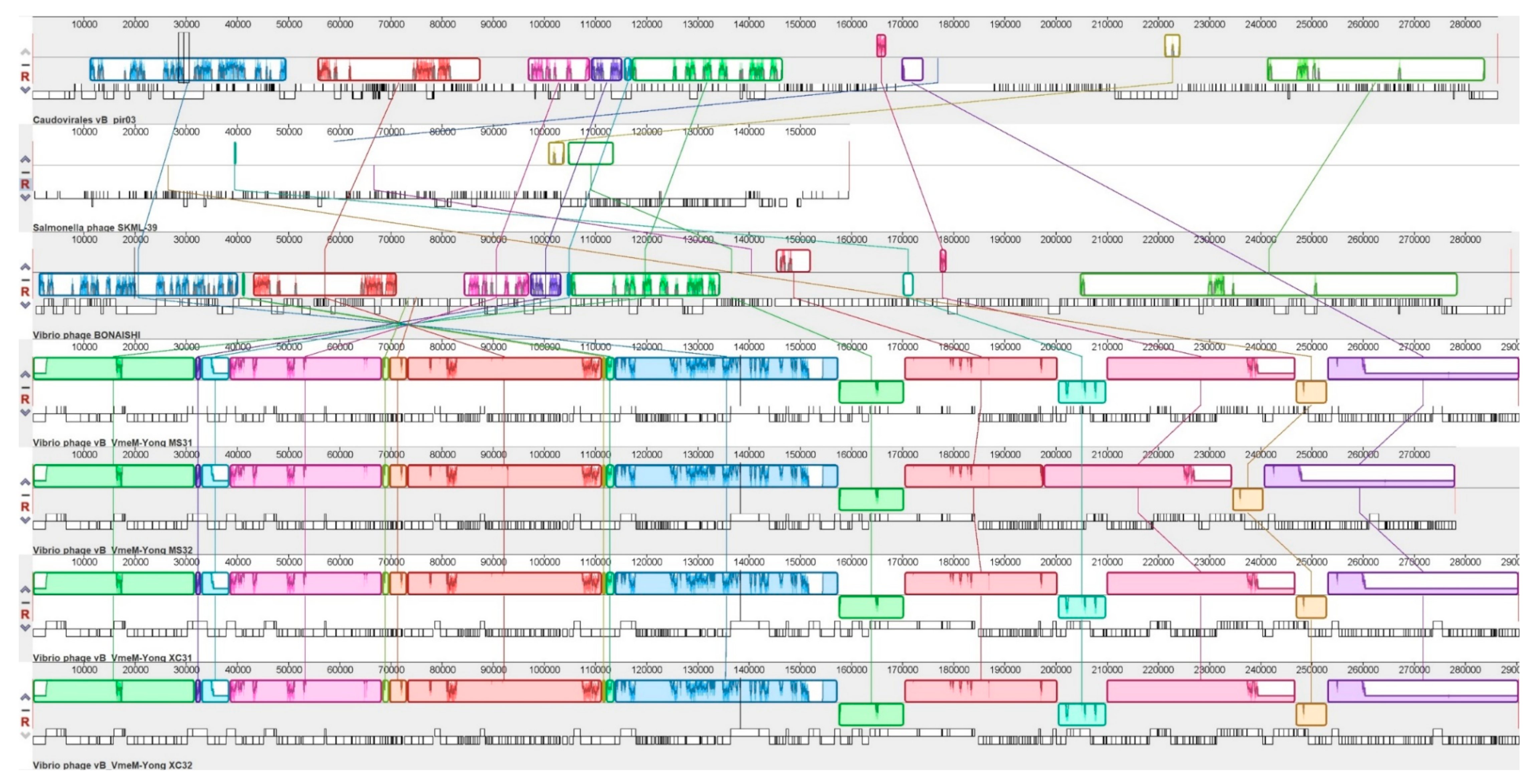
2.8. Genomic Synteny of vB_VhaM_pir03 with Other Similar Phages
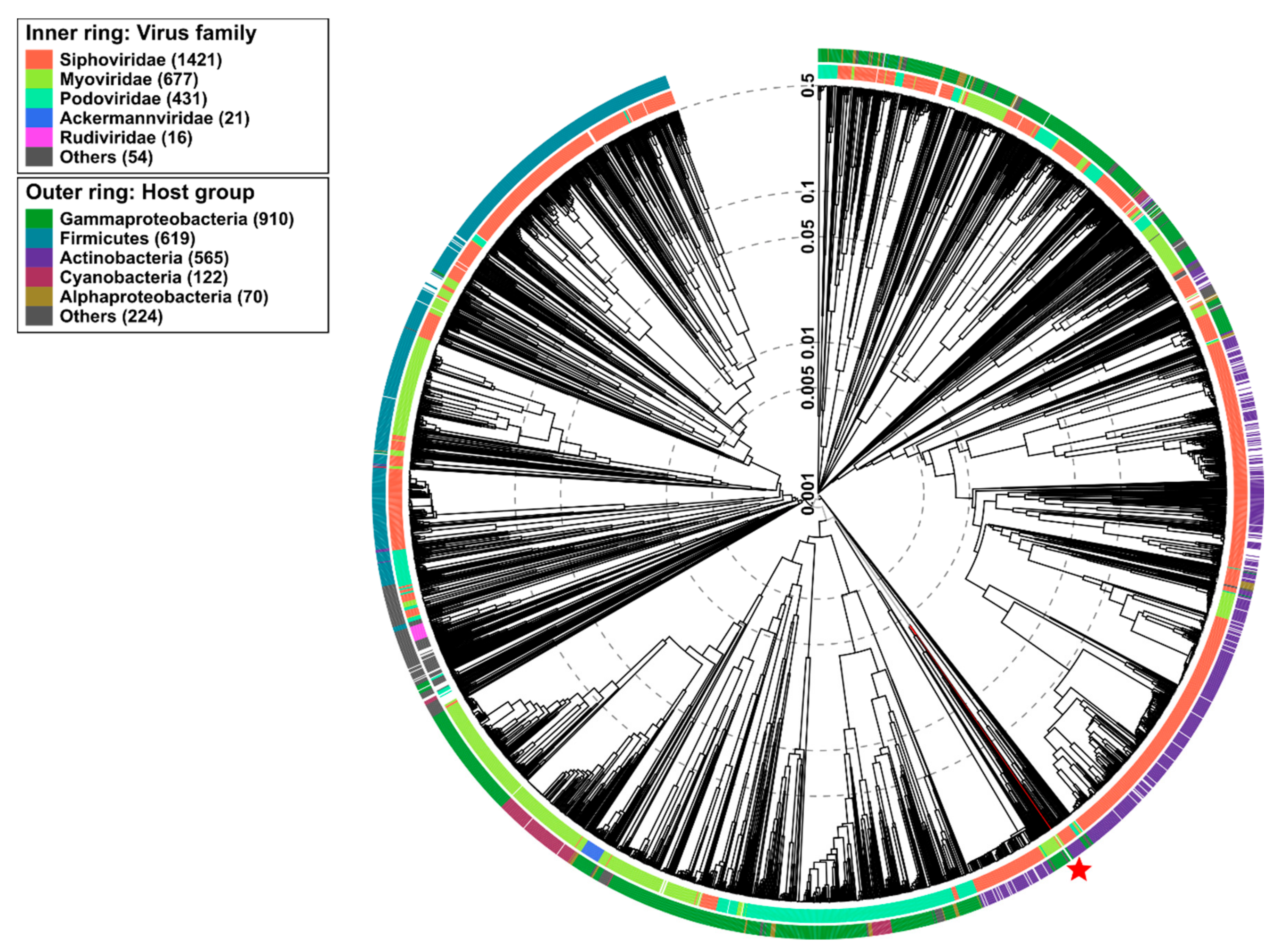
2.9. Phylogenetic Analysis
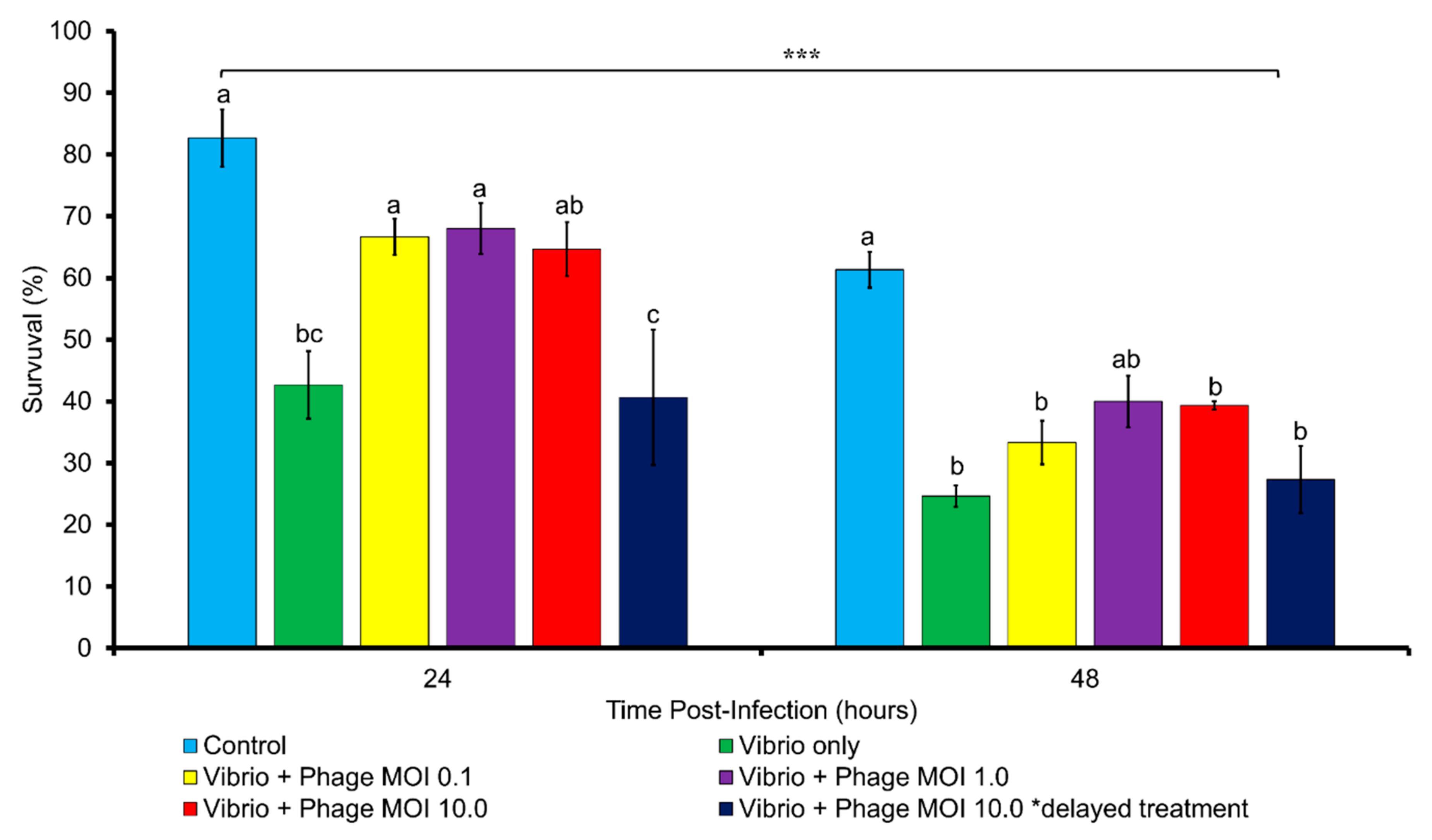
2.10. In Vivo Phage Therapy Trial with Artemia nauplii
3. Discussion
4. Conclusions and Future Directions
5. Materials and Methods
5.1. Bacterial Strains Used in This Study
5.2. Antibiotic Susceptibility Testing
5.3. Isolation and Purification of Bacteriophages
5.4. Transmission Electron Microscopy
5.5. Host Range Test
5.6. Efficiency of Plating (EOP)
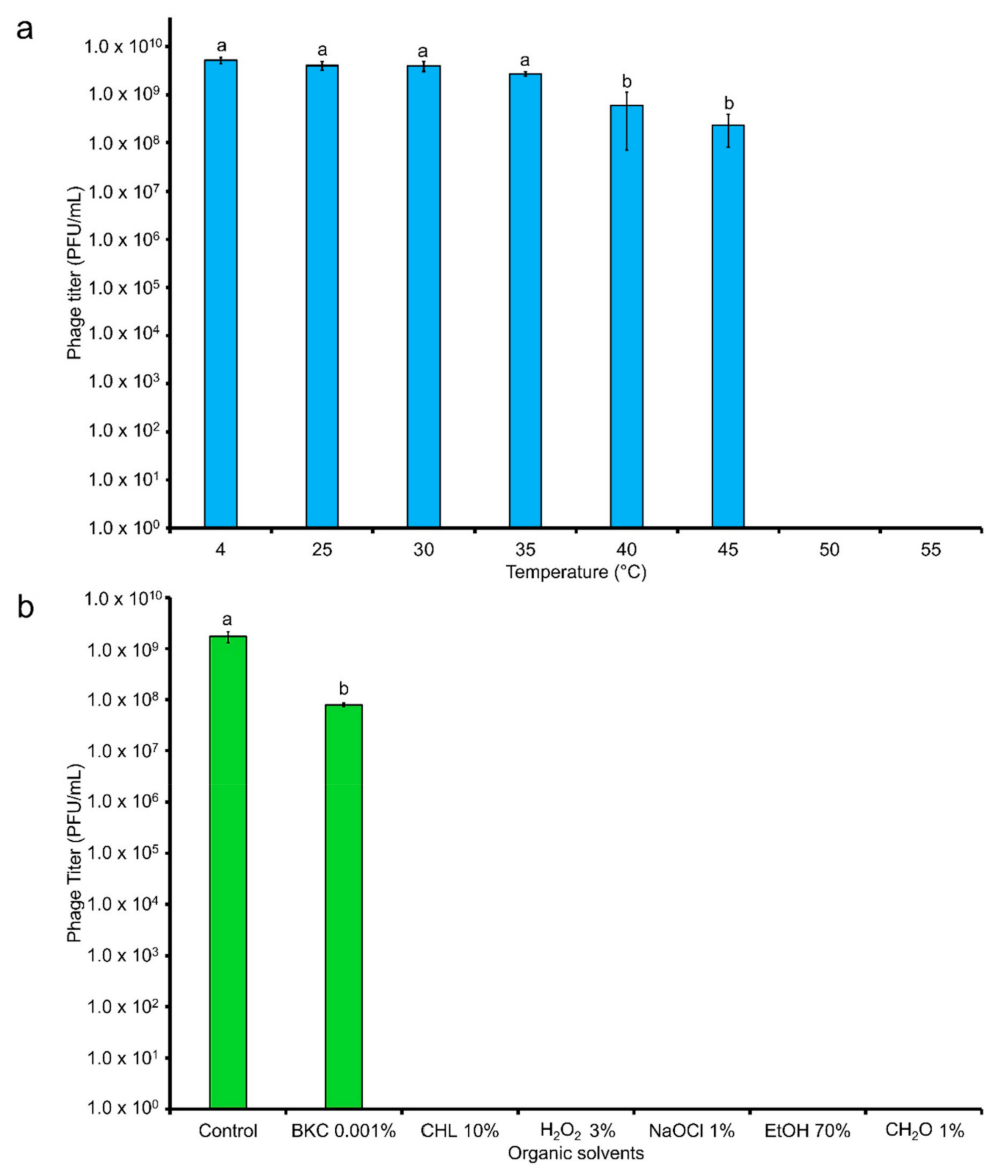
5.7. Stability of Phage in Different Temperatures and Organic Solvents
5.8. Adsorption Time and One-Step Growth
5.9. In Vitro Cell Lysis
5.10. DNA Extraction and Purification
5.11. Genomic Analysis
5.12. Genome Alignment and Phylogenetic Analysis of vB_VhaM_pir03
5.13. In Vivo Phage Therapy Trial Using Artemia nauplii
5.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pridgeon, J.W.; Klesius, P.H. Major bacterial diseases in aquaculture and their vaccine development. Cab Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Wang, W. Bacterial diseases of crabs: A review. J. Invertebr. Pathol. 2011, 106, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Cardinaud, M.; Barbou, A.; Capitaine, C.; Bidault, A.; Dujon, A.M.; Moraga, D.; Paillard, C. Vibrio harveyi adheres to and penetrates tissues of the European abalone Haliotis tuberculata within the first hours of contact. Appl. Environ. Microbiol. 2014, 80, 6328–6333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darshanee Ruwandeepika, H.A.; Sanjeewa Prasad Jayaweera, T.; Paban Bhowmick, P.; Karunasagar, I.; Bossier, P.; Defoirdt, T. Pathogenesis, virulence factors and virulence regulation of Vibrios belonging to the Harveyi clade. Rev. Aquac. 2012, 4, 59–74. [Google Scholar] [CrossRef]
- Chrisolite, B.; Thiyagarajan, S.; Alavandi, S.V.; Abhilash, E.C.; Kalaimani, N.; Vijayan, K.K.; Santiago, T.C. Distribution of luminescent Vibrio harveyi and their bacteriophages in a commercial shrimp hatchery in South India. Aquaculture 2008, 275, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Haldar, S.; Maharajan, A.; Chatterjee, S.; Hunter, S.A.; Chowdhury, N.; Hinenoya, A.; Asakura, M.; Yamasaki, S. Identification of Vibrio harveyi as a causative bacterium for a tail rot disease of sea bream Sparus aurata from research hatchery in Malta. Microbiol. Res. 2010, 165, 639–648. [Google Scholar] [CrossRef]
- Pujalte, M.; Sitjà-Bobadilla, A.; Álvarez-Pellitero, P.; Garay, E. Carriage of potentially fish-pathogenic bacteria in Sparus aurata cultured in Mediterranean fish farms. Dis. Aquat. Org. 2003, 54, 119–126. [Google Scholar] [CrossRef]
- Pujalte, M.J.; Sitjà-Bobadilla, A.; Macián, M.C.; Belloch, C.; Álvarez-Pellitero, P.; Pérez-Sánchez, J.; Uruburu, F.; Garay, E. Virulence and molecular typing of Vibrio harveyi strains isolated from cultured dentex, gilthead sea bream and European sea bass. Syst. Appl. Microbiol. 2003, 26, 284–292. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Stockley, L.; Rangdale, R.; Martinez-Urtaza, J. Environmental occurrence and clinical impact of Vibrio vulnificus and Vibrio parahaemolyticus: A European perspective. Environ. Microbiol. Rep. 2010, 2, 7–18. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Trinanes, J.A.; Taylor, N.G.H.; Hartnell, R.; Siitonen, A.; Martinez-Urtaza, J. Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Chang. 2013, 3, 73–77. [Google Scholar] [CrossRef]
- Elston, R.; Hasegawa, H.; Humphrey, K.; Polyak, I.; Häse, C. Re-emergence of Vibrio tubiashii in bivalve shellfish aquaculture: Severity, environmental drivers, geographic extent and management. Dis. Aquat. Org. 2008, 82, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.; Marques, A.; Nunes, M.L. Impact of climate change in Mediterranean aquaculture. Rev. Aquac. 2012, 4, 163–177. [Google Scholar] [CrossRef]
- Lulijwa, R.; Rupia, E.J.; Alfaro, A.C. Antibiotic use in aquaculture, policies and regulation, health and environmental risks: A review of the top 15 major producers. Rev. Aquac. 2020, 12, 640–663. [Google Scholar] [CrossRef]
- Cabello, F.C.; Godfrey, H.P.; Tomova, A.; Ivanova, L.; Dölz, H.; Millanao, A.; Buschmann, A.H. Antimicrobial use in aquaculture re-examined: Its relevance to antimicrobial resistance and to animal and human health. Environ. Microbiol. 2013, 15, 1917–1942. [Google Scholar] [CrossRef] [PubMed]
- Husevag, B.; Lunestad, B.T.; Johannessen, P.J.; Samuelsen, O.B. Simultaneous occurrence of Vibrio salmonicida and antibiotic-resistant bacteria in sediments at abandoned aquaculture sites. J. Fish Dis. 1991, 14, 631–640. [Google Scholar] [CrossRef]
- Jang, H.M.; Kim, Y.B.; Choi, S.; Lee, Y.; Shin, S.G.; Unno, T.; Kim, Y.M. Prevalence of antibiotic resistance genes from effluent of coastal aquaculture, South Korea. Environ. Pollut. 2018, 233, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Karunasagar, I.; Pai, R.; Malathi, G.R.; Karunasagar, I. Mass mortality of Penaeus monodon larvae due to antibiotic-resistant Vibrio harveyi infection. Aquaculture 1994, 128, 203–209. [Google Scholar] [CrossRef]
- Miranda, C.D.; Tello, A.; Keen, P.L. Mechanisms of antimicrobial resistance in finfish aquaculture environments. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Smith, P. Antimicrobial resistance in aquaculture. Rev. Sci. Tech. 2008, 27, 243–264. [Google Scholar] [CrossRef]
- Doss, J.; Culbertson, K.; Hahn, D.; Camacho, J.; Barekzi, N. A review of phage therapy against bacterial pathogens of aquatic and terrestrial organisms. Viruses 2017, 9, 50. [Google Scholar] [CrossRef] [Green Version]
- Gon Choudhury, T.; Tharabenahalli Nagaraju, V.; Gita, S.; Paria, A.; Parhi, J. Advances in Bacteriophage Research for Bacterial Disease Control in Aquaculture. Rev. Fish. Sci. Aquac. 2017, 25, 113–125. [Google Scholar] [CrossRef]
- Nakai, T. Application of bacteriophages for control of infectious diseases in aquaculture. Bacteriophages Control Food Waterborne Pathog. 2010, 257–272. [Google Scholar] [CrossRef]
- Jun, J.W.; Han, J.E.; Tang, K.F.J.; Lightner, D.V.; Kim, J.; Seo, S.W.; Park, S.C. Potential application of bacteriophage pVp-1: Agent combating Vibrio parahaemolyticus strains associated with acute hepatopancreatic necrosis disease (AHPND) in shrimp. Aquaculture 2016, 457, 100–103. [Google Scholar] [CrossRef] [Green Version]
- Kalatzis, P.G.; Bastías, R.; Kokkari, C.; Katharios, P. Isolation and characterization of two lytic bacteriophages, φst2 and φgrn1; Phage therapy application for biological control of Vibrio alginolyticus in aquaculture live feeds. PLoS ONE 2016, 11, e151101. [Google Scholar] [CrossRef] [Green Version]
- Katharios, P.; Kalatzis, P.G.; Kokkari, C.; Sarropoulou, E.; Middelboe, M. Isolation and characterization of a N4-like lytic bacteriophage infecting Vibrio splendidus, a pathogen of fish and bivalves. PLoS ONE 2017, 12, e190083. [Google Scholar] [CrossRef] [Green Version]
- Quiroz-Guzmán, E.; Peña-Rodriguez, A.; Vázquez-Juárez, R.; Barajas-Sandoval, D.R.; Balcázar, J.L.; Martínez-Díaz, S.F. Bacteriophage cocktails as an environmentally-friendly approach to prevent Vibrio parahaemolyticus and Vibrio harveyi infections in brine shrimp (Artemia franciscana) production. Aquaculture 2018, 492, 273–279. [Google Scholar] [CrossRef]
- Stenholm, A.R.; Dalsgaard, I.; Middelboe, M. Isolation and characterization of bacteriophages infecting the fish pathogen Flavobacterium psychrophilum. Appl. Environ. Microbiol. 2008, 74, 4070–4078. [Google Scholar] [CrossRef] [Green Version]
- Hyman, P. Phages for phage therapy: Isolation, characterization, and host range breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Jacquemot, L.; Bettarel, Y.; Monjol, J.; Corre, E.; Halary, S.; Desnues, C.; Bouvier, T.; Ferrier-Pagès, C.; Baudoux, A.C. Therapeutic potential of a new jumbo phage that infects Vibrio coralliilyticus, a widespread coral pathogen. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Shneider, M.M.; Buth, S.A.; Ho, B.T.; Basler, M.; Mekalanos, J.J.; Leiman, P.G. PAAR-repeat proteins sharpen and diversify the type VI secretion system spike. Nature 2013, 500, 350–353. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Shen, N.; Pei, J.; Grishin, N.V. Double-stranded DNA bacteriophage prohead protease is homologous to herpesvirus protease. Protein Sci. 2004, 13, 2260–2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, J.R.; Geiduschek, E.P. Site-specific DNA binding by the bacteriophage SPOl-encoded type HI DNA-binding protein. EMBO J. 1985, 4, 1345–1349. [Google Scholar] [CrossRef] [PubMed]
- Rubio Gomez, M.A.; Ibba, M. Aminoacyl-tRNA Synthetases. RNA 2020. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syn. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Abedon, S.T.; García, P.; Mullany, P.; Aminov, R. Editorial: Phage therapy: Past, present and future. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Custus (EN)—ACD Pharma. Available online: https://acdpharma.com/en/bacteriophage-products/custus-en/ (accessed on 17 July 2020).
- Abedon, S.T. Information phage therapy research should report. Pharmaceuticals 2017, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Casey, E.; van Sinderen, D.; Mahony, J. In Vitro Characteristics of Phages to Guide “Real Life” Phage Therapy Suitability. Viruses 2018, 10, 163. [Google Scholar] [CrossRef] [Green Version]
- Hyman, P.; Abedon, S.T. Smaller Fleas: Viruses of Microorganisms. Scientifica 2012, 2012, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Seguritan, V.; Feng, I.W.; Rohwer, F.; Swift, M.; Segall, A.M. Genome Sequences of Two Closely Related Vibrio parahaemolyticus Phages, VP16T and VP16C. J. Bacteriol. 2003, 185, 6434–6447. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.; Gram, L.; Middelboe, M. Vibriophages and their interactions with the fish pathogen Vibrio anguillarum. Appl. Environ. Microbiol. 2014, 80, 3128–3140. [Google Scholar] [CrossRef] [Green Version]
- Hua, J.; Huet, A.; Lopez, C.A.; Toropova, K.; Pope, W.H.; Duda, R.L.; Hendrix, R.W.; Conway, J.F. Capsids and genomes of jumbo-sized bacteriophages reveal the evolutionary reach of the HK97 fold. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.P.; Gong, T.; Jost, G.; Liu, W.H.; Ye, D.Z.; Luo, Z.H. Isolation and characterization of five lytic bacteriophages infecting a Vibrio strain closely related to Vibrio owensii. FEMS Microbiol. Lett. 2013, 348, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.G.; Jun, J.W.; Giri, S.S.; Yun, S.; Kim, H.J.; Kim, S.W.; Kang, J.W.; Han, S.J.; Jeong, D.; Park, S.C. Isolation and characterisation of pVa-21, a giant bacteriophage with anti-biofilm potential against Vibrio alginolyticus. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, H.; Radford, D.; Kropinski, A.M.; Lim, L.T.; Balamurugan, S. Thermal-stability and reconstitution ability of Listeria phages P100 and A511. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- De Paepe, M.; Taddei, F. Viruses’ life history: Towards a mechanistic basis of a trade-off between survival and reproduction among phages. PLoS Biol. 2006, 4, 1248–1256. [Google Scholar] [CrossRef] [Green Version]
- Purohit, P.K.; Inamdar, M.M.; Grayson, P.D.; Squires, T.M.; Kondev, J.; Phillips, R. Forces during bacteriophage DNA packaging and ejection. Biophys. J. 2005, 88, 851–866. [Google Scholar] [CrossRef] [Green Version]
- Mäntynen, S.; Sundberg, L.-R.; Oksanen, H.M.; Poranen, M.M. Half a Century of Research on Membrane-Containing Bacteriophages: Bringing New Concepts to Modern Virology. Viruses 2019, 11, 76. [Google Scholar] [CrossRef] [Green Version]
- Jurczak-Kurek, A.; Gąsior, T.; Nejman-Faleńczyk, B.; Bloch, S.; Dydecka, A.; Topka, G.; Necel, A.; Jakubowska-Deredas, M.; Narajczyk, M.; Richert, M.; et al. Biodiversity of bacteriophages: Morphological and biological properties of a large group of phages isolated from urban sewage. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Quiberoni, A.; Suárez, V.B.; Suárez, S.; Reinheimer, J.A. Inactivation of Lactobacillus helveticus Bacteriophages by Thermal and Chemical Treatments. J. Food Prot. 1999, 62, 894–898. [Google Scholar] [CrossRef]
- Higuera, G.; Bastías, R.; Tsertsvadze, G.; Romero, J.; Espejo, R.T. Recently discovered Vibrio anguillarum phages can protect against experimentally induced vibriosis in Atlantic salmon, Salmo salar. Aquaculture 2013, 392–395, 128–133. [Google Scholar] [CrossRef]
- Lal, T.M.; Sano, M.; Ransangan, J. Genome characterization of a novel vibriophage VpKK5 (Siphoviridae) specific to fish pathogenic strain of Vibrio parahaemolyticus. J. Basic Microbiol. 2016, 56, 872–888. [Google Scholar] [CrossRef] [PubMed]
- Mateus, L.; Costa, L.; Silva, Y.J.; Pereira, C.; Cunha, A.; Almeida, A. Efficiency of phage cocktails in the inactivation of Vibrio in aquaculture. Aquaculture 2014, 424–425, 167–173. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, I.N. Bacteriophage adsorption rate and optimal lysis time. Genetics 2008, 180, 471–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Menéndez, E.; Arroyo-López, F.N.; Martínez, B.; García, P.; Garrido-Fernández, A.; Rodríguez, A. Optimizing propagation of Staphylococcus aureus infecting bacteriophage vB_sauM-phiIPLA-RODI on Staphylococcus xylosus using response surface methodology. Viruses 2018, 10, 153. [Google Scholar] [CrossRef] [Green Version]
- Imam, M.; Alrashid, B.; Patel, F.; Dowah, A.S.A.; Brown, N.; Millard, A.; Clokie, M.R.J.; Galyov, E.E. vB_PaeM_MIJ3, a Novel Jumbo Phage Infecting Pseudomonas aeruginosa, Possesses Unusual Genomic Features. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yuan, S.; Liu, Q.; Mai, G.; Yang, J.; Deng, D.; Zhang, B.; Liu, C.; Ma, Y. In Vitro design and evaluation of phage cocktails against Aeromonas salmonicida. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Hatfull, G.F.; Hendrix, R.W. Bacteriophages and their genomes. Curr. Opin. Virol. 2011, 1, 298–303. [Google Scholar] [CrossRef] [Green Version]
- Comeau, A.M.; Bertrand, C.; Letarov, A.; Tétart, F.; Krisch, H.M. Modular architecture of the T4 phage superfamily: A conserved core genome and a plastic periphery. Virology 2007, 362, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Gao, M. Jumbo bacteriophages: An overview. Front. Microbiol. 2017, 8, 403. [Google Scholar] [CrossRef] [Green Version]
- Leskinen, K.; Blasdel, B.G.; Lavigne, R.; Skurnik, M. RNA-sequencing reveals the progression of Phage-Host interactions between φR1-37 and Yersinia enterocolitica. Viruses 2016, 8, 111. [Google Scholar] [CrossRef] [Green Version]
- Röske, K.; Foecking, M.F.; Yooseph, S.; Glass, J.I.; Calcutt, M.J.; Wise, K.S. A versatile palindromic amphipathic repeat coding sequence horizontally distributed among diverse bacterial and eucaryotic microbes. BMC Genom. 2010, 11, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moniruzzaman, M.; Wurch, L.L.; Alexander, H.; Dyhrman, S.T.; Gobler, C.J.; Wilhelm, S.W. Virus-host relationships of marine single-celled eukaryotes resolved from metatranscriptomics. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yakovchuk, P.; Protozanova, E.; Frank-Kamenetskii, M.D. Base-stacking and base-pairing contributions into thermal stability of the DNA double helix. Nucleic Acids Res. 2006, 34, 564–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, S.; Wolin, S.L. Bacterial Y RNAs: Gates, Tethers and tRNA Mimics. Mircrobiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Hendrix, R.W. Jumbo Bacteriophages. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2009; Volume 328, pp. 229–240. [Google Scholar]
- Prasad Bhattacharyya, S.; Basaveswara Rao, V. A novel terminase activity associated with the DNA packaging protein gp17 of bacteriophage T4. Virology 1993, 196, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Garneau, J.R.; Depardieu, F.; Fortier, L.C.; Bikard, D.; Monot, M. PhageTerm: A tool for fast and accurate determination of phage termini and packaging mechanism using next-generation sequencing data. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.M.; Liu, D.; Ogura, Y.; Hayashi, T.; Urbanczyk, H.; Tsai, I.J. Tracing genomic divergence of Vibrio bacteria in the Harveyi clade. J. Bacteriol. 2018, 200. [Google Scholar] [CrossRef] [Green Version]
- Urbanczyk, H.; Ogura, Y.; Hayashi, T. Taxonomic revision of Harveyi clade bacteria (family Vibrionaceae) based on analysis of whole genome sequences. Int. J. Syst. Evol. Microbiol. 2013, 63, 2742–2751. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Tanaka, S.; Koga, T.; Kawata, T. A Broad-Host-Range Vibriophage, KVP40, Isolated from Sea Water. Microbiol. Immunol. 1992, 36, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Holmfeldt, K.; Middelboe, M.; Nybroe, O.; Riemann, L. Large variabilities in host strain susceptibility and phage host range govern interactions between lytic marine phages and their Flavobacterium hosts. Appl. Environ. Microbiol. 2007, 73, 6730–6739. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, A.; Fujisawa, M.; Hamasaki, R.; Kawasaki, T.; Fujie, M.; Yamada, T. Biocontrol of Ralstonia solanacearum by treatment with lytic bacteriophages. Appl. Environ. Microbiol. 2011, 77, 4155–4162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, G.; Askora, A.; Blanc-Mathieu, R.; Kawasaki, T.; Li, Y.; Nakano, M.; Ogata, H.; Yamada, T. Xanthomonas citri jumbo phage XacN1 exhibits a wide host range and high complement of tRNA genes. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.; Ward, S.; Hyman, P. More is better: Selecting for broad host range bacteriophages. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, F.L.; Iida, T.; Swings, J. Biodiversity of Vibrios. Microbiol. Mol. Biol. Rev. 2004, 68, 403–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. infections. Nat. Rev. Dis. Primers 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Jo, Y.; Hwang, Y.J.; Hong, H.W.; Hong, S.S.; Park, K.; Myung, H. Phageantibiotic synergy via delayed lysis. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowska, K. Phage therapy: What factors shape phage pharmacokinetics and bioavailability? Systematic and critical review. Med. Res. Rev. 2019, 39, 2000–2025. [Google Scholar] [CrossRef] [Green Version]
- Middelboe, M.; Hagström, A.; Blackburn, N.; Sinn, B.; Fischer, U.; Borch, N.H.; Pinhassi, J.; Simu, K.; Lorenz, M.G. Effects of bacteriophages on the population dynamics of four strains of pelagic marine bacteria. Microb. Ecol. 2001, 42, 395–406. [Google Scholar] [CrossRef]
- Cenens, W.; Makumi, A.; Mebrhatu, M.T.; Lavigne, R.; Aertsen, A. Phage–host interactions during pseudolysogeny. Bacteriophage 2013, 3, e25029. [Google Scholar] [CrossRef] [Green Version]
- Olszak, T.; Danis-Wlodarczyk, K.; Arabski, M.; Gula, G.; Maciejewska, B.; Wasik, S.; Lood, C.; Higgins, G.; Harvey, B.J.; Lavigne, R.; et al. Pseudomonas aeruginosa PA5oct jumbo phage impacts planktonic and biofilm population and reduces its host virulence. Viruses 2019, 11, 1089. [Google Scholar] [CrossRef] [Green Version]
- Siringan, P.; Connerton, P.L.; Cummings, N.J.; Connerton, I.F. Alternative bacteriophage life cycles: The carrier state of Campylobacter jejuni. Open Biol. 2014, 4, 130200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Díaz, S.F.; Hipólito-Morales, A. Efficacy of phage therapy to prevent mortality during the vibriosis of brine shrimp. Aquaculture 2013, 400–401, 120–124. [Google Scholar] [CrossRef]
- Żaczek, M.; Weber-Dabrowska˛, B.; Górski, A. Phages as a cohesive prophylactic and therapeutic approach in aquaculture systems. Antibiotics 2020, 9, 564. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.J.; Costa, L.; Pereira, C.; Mateus, C.; Cunha, Â.; Calado, R.; Gomes, N.C.M.; Pardo, M.A.; Hernandez, I.; Almeida, A. Phage Therapy as an Approach to Prevent Vibrio anguillarum Infections in Fish Larvae Production. PLoS ONE 2014, 9, e114197. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic Susceptibility Testing by a Standardized Single Disk Method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. M45: Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, 3rd ed.; CLSI: Wayne, PA, USA, 2016. [Google Scholar]
- Clinical and Laboratory Standards Institute. M100-S25; Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement; CLSI: Wayne, PA, USA, 2015. [Google Scholar]
- Martha Clokie, R.J.; Kropinski, A.M. (Eds.) Bacteriophages: Methods and Protocols Volume 1: Isolation, Characterization, and Interactions; Humana Press: New York, NY, USA, 2009; Volume 501. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Kutter, E. Phage host range and efficiency of plating. Methods Mol. Biol. 2009, 501, 141–149. [Google Scholar] [CrossRef]
- Misol, G.N.; Kokkari, C.; Katharios, P. Complete Genome Sequence of a Jumbo Bacteriophage, vB_pir03, against Vibrio harveyi. Microbiol. Resour. Announc. 2020, 9, 1–2. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Bushnell, B. BBMap: A fast, accurate, splice-aware aligner. In Proceedings of the Conference: 9th Annual Genomics of Energy & Environment Meeting, Walnut Creek, CA, USA, 17–20 March 2014. [Google Scholar]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Ech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- Altschup, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Gene Ontology Consortium. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2019, 47, D351–D360. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33. [Google Scholar] [CrossRef] [Green Version]
- Moura, A.; Soares, M.; Pereira, C.; Leitão, N.; Henriques, I.; Correia, A. INTEGRALL: A database and search engine for integrons, integrases and gene cassettes. Bioinformatics 2009, 25, 1096–1098. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Kleinheinz, K.A.; Joensen, K.G.; Larsen, M.V. Applying the ResFinder and VirulenceFinder web-services for easy identification of acquired antibiotic resistance and E. coli virulence genes in bacteriophage and prophage nucleotide sequences. Bacteriophage 2014, 4, e27943. [Google Scholar] [CrossRef] [Green Version]
- Lima-Mendez, G.; van Helden, J.; Toussaint, A.; Leplae, R. Prophinder: A computational tool for prophage prediction in prokaryotic genomes. Bioinformatics 2008, 24, 863–865. [Google Scholar] [CrossRef] [Green Version]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darling, A.E.; Mau, B.; Perna, N.T. Progressivemauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The viral proteomic tree server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. BMC Bioinform. 2004, 5. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. CABIOS 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, 242–245. [Google Scholar] [CrossRef]
- Gomez Gil-RS, B.; Abreu-Grobois, F.A.; Romero-Jarero, J.; de los Herrera-Vega, M. Chemical Disinfection of Artemia Nauplii. J. World Aquac. Soc. 1994, 25, 579–583. [Google Scholar] [CrossRef]
- Crab, R.; Lambert, A.; Defoirdt, T.; Bossier, P.; Verstraete, W. The application of bioflocs technology to protect brine shrimp (Artemia franciscana) from pathogenic Vibrio harveyi. J. Appl. Microbiol. 2010, 109, 1643–1649. [Google Scholar] [CrossRef]
- Haynes, W. Tukey’s Test. In Encyclopedia of Systems Biology; Werner, D., Wolkenhauer, O., Yokota, H., Cho, K.H., Eds.; Springer: New York, NY, USA, 2013; pp. 2303–2304. ISBN 978-1-4419-9863-7. [Google Scholar]
- Hammer, Ø. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Efficiency of Plating of vB_VhaM_pir03 | Antibiotic Susceptibility Testing | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species/Strain | Host Range | EOP | Zone of Inhibition Diameter (mm) | |||||||||||
| 100 | 10−1 | 10−2 | 10−3 | 10−4 | 10−5 | (%) | AMP | TE | OT | OA | FFC | SXT | UB | |
| Vibrio harveyi | ||||||||||||||
| *DSM19623 | +++ | +++ | +++ | +++ | ++ | + | High | R | S | I | R | S | S | S |
| SNGR | +++ | +++ | +++ | +++ | ++ | ++ | High | R | S | R | I | S | R | S |
| KS6 | ++ | ++ | ++ | + | + | - | Low | R | S | R | R | R | R | S |
| Vh2 | +++ | +++ | ++ | ++ | + | - | Medium | R | S | S | R | S | I | S |
| Vh5 | +++ | +++ | +++ | +++ | ++ | + | High | R | S | I | S | S | S | S |
| VhSernFr | +++ | +++ | +++ | +++ | ++ | + | High | R | S | S | R | I | I | S |
| VhP1Liv | +++ | +++ | +++ | +++ | ++ | + | High | R | S | I | S | S | S | S |
| Vhp1Spl | ++ | + | + | - | - | - | Low | |||||||
| VhKarx | ++ | - | - | - | - | - | NF | R | S | I | R | S | S | S |
| RG1 | +++ | ++ | ++ | + | - | - | Medium | R | S | R | S | S | S | S |
| Barb A4/1.1 | - | - | - | - | - | - | NF | R | S | I | I | S | S | S |
| SerKid | +++ | +++ | +++ | +++ | ++ | + | Medium | R | S | I | I | S | S | S |
| SerKid2 | +++ | +++ | +++ | +++ | ++ | + | High | |||||||
| SerSd | +++ | ++ | ++ | ++ | + | + | High | |||||||
| SA 5.1 | +++ | +++ | ++ | ++ | ++ | + | Medium | R | S | I | I | S | R | S |
| SA 6.1 | ++ | ++ | + | + | - | - | Low | R | S | I | I | R | R | R |
| SA 9.2 | ++ | ++ | ++ | + | + | + | Medium | R | S | R | I | S | R | I |
| SA 1.2 | ++ | ++ | + | + | - | - | Low | R | S | I | R | R | R | R |
| SA 7.1 | +++ | +++ | ++ | + | - | - | Low | R | S | S | I | I | R | S |
| SA 3.1 | +++ | +++ | ++ | ++ | ++ | + | Medium | R | S | I | R | S | R | S |
| SA 4.1 | +++ | + | + | + | - | - | Low | R | S | I | I | R | R | R |
| SA 2.1 | ++ | + | + | + | - | - | Low | R | S | R | R | S | S | S |
| Vh No. 22 | ++ | ++ | ++ | + | - | - | Low | R | S | I | I | I | S | I |
| Vh6 | - | - | - | - | - | - | NF | R | S | I | I | S | I | S |
| Vibrio alginolyticus | ||||||||||||||
| V1 | +++ | +++ | +++ | +++ | ++ | + | High | R | S | I | I | S | S | S |
| V2 | +++ | +++ | ++ | ++ | + | - | Low | R | S | I | I | S | S | S |
| HCMR 1 Art. 3 | ++ | ++ | ++ | ++ | + | - | High | R | S | S | I | S | R | S |
| Vibrio campbellii | ||||||||||||||
| VIB391 | +++ | +++ | +++ | ++ | - | - | Medium | R | S | I | I | S | S | S |
| Vibrio owensii | ||||||||||||||
| SA 1.1 | ++ | ++ | ++ | ++ | + | + | Medium | R | S | I | R | R | R | R |
| SA 9.1 | ++ | + | + | + | - | - | Low | R | S | R | I | R | R | S |
| Other Vibrio spp. | ||||||||||||||
| Art. 2 | +++ | +++ | ++ | ++ | ++ | + | High | R | S | R | I | R | R | R |
| Rot. 2 | +++ | ++ | - | - | - | - | Low | R | S | R | S | R | R | I |
| Barb A4/1.2 | +++ | +++ | ++ | ++ | + | - | High | R | S | I | I | S | S | S |
| Rot. Vib. 5 | +++ | +++ | +++ | ++ | - | - | Medium | R | S | S | I | R | R | S |
| Predicted Functions | Start | End | Length | Direction | NCBI BLASTP Best Hit | E-Value | Similarity Score (%) | Gene Onthology | InterPro | NCBI CDD Best Hit | E-Value | HHPRED Best Hit | E-Value | Probability (%) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ORF 2 | Baseplate structural protein | 3242 | 6268 | 3027 | Reverse | Vibrio phage BONAISHI AXH71039.1 | 1.05 × 10−15 | 42.48 | cl33689|long tail fiber, proximal subunit | 5.51 × 10−3 | 1S2E_B|Bacteriophage T4 | 1.20 × 10−5 | 98.20 | ||
| ORF 3 | Hypothetical protein | 6278 | 7180 | 903 | Reverse | Vibrio phage BONAISHI AXH71040.1 | 2.0 × 10−40 | 57.70 | |||||||
| ORF 4 | Putative tail protein | 7192 | 8181 | 990 | Reverse | Vibrio phage BONAISHI AXH71041.1 | 5.39 × 10−72 | 56.00 | |||||||
| ORF 6 | Hypothetical protein | 8315 | 9526 | 1212 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96157.1 | 3.0 × 10−30 | 47.20 | |||||||
| ORF 7 | ABC-type ATPase | 9568 | 12,276 | 2709 | Reverse | Vibrio phage vB_VmeM-Yong XC31 QAX96150.1 | 1.76 × 10−109 | 50.20 | cd00267|ATP-binding cassette transporter nucleotide-binding domain | 1.30 × 10−7 | 6S6V_D|Escherichia coli | 4.40 × 10−23 | 99.96 | ||
| ORF 8 | Membrane-puncturing device | 12,113 | 12,937 | 825 | Forward | Vibrio phage BONAISHI AXH70744.1 | 2.51 × 10−66 | 61.50 | 6ORJ_A|Pseudomonas virus phiKZ | 4.60 × 10−89 | 100.00 | ||||
| ORF 9 | Hypothetical protein | 12,947 | 13,567 | 621 | Forward | Vibrio phage BONAISHI AXH70745.1 | 1.43 × 10−10 | 56.31 | |||||||
| ORF 10 | PAAR-repeat containing protein | 13,560 | 13,859 | 300 | Forward | Vibrio phage BONAISHI AXH70746.1 | 1.30 × 10−21 | 69.40 | IPR008727 | cd14737|proline-alanine-alanine-arginine (PAAR) domain | 1.46 × 10−29 | 4KU0_D|Bacteriophage T4 | 2.70 × 10−11 | 99.40 | |
| ORF 11 | PAAR-motif protein | 13,718 | 13,891 | 174 | Forward | Vibrio phage 03O._10N.2646.F8] AUR83144.1 | 9.00 × 10−3 | 6.64 | |||||||
| ORF 12 | Hypothetical protein | 13,901 | 14,512 | 612 | Reverse | Vibrio phage vB_VmeM-Yong XC31 QAX96156.1 | 4.76 × 10−31 | 54.80 | |||||||
| ORF 13 | Hypothetical protein | 14,523 | 158,181 | 1296 | Reverse | Vibrio phage vB_VmeM-Yong XC31 QAX96155.1 | 8.43 × 10−68 | 53.60 | |||||||
| ORF 16 | Ribonuclease HI (EC 3.1.26.4) CDS | 16,667 | 18,028 | 1362 | Forward | Vibrio phage BONAISHI AXH70751.1 | 1.35 × 10−59 | 48.00 | F: GO:0016787 | IPR036397 | cd09278|RNase HI family found mainly in prokaryotes | 1.40 × 10−36 | 4MH8_A|Moloney murine leukemia virus | 2.90 × 10−9 | 99.07 |
| ORF 17 | Hypothetical protein | 18,081 | 18,920 | 840 | Reverse | Vibrio phage BONAISHI AXH70752.1 | 8.97 × 10−21 | 50.70 | |||||||
| ORF 18 | UvsX protein | 18,757 | 20,484 | 1728 | Forward | Vibrio phage BONAISHI AXH70753.1 | 3.12 × 10−171 | 67.50 | 3IO5_B|Bacteriophage T4. | 4.60 × 10−26 | 99.95 | ||||
| ORF 20 | DNA-directed RNA polymerase | 20,726 | 21,496 | 771 | Forward | Vibrio phage BONAISHI AXH70756.1 | 2.85 × 10−61 | 66.70 | 2A6H_M|Thermus thermophilus | 3.80 × 10−32 | 99.98 | ||||
| ORF 21 | Hypothetical protein | 21,489 | 21,896 | 408 | Forward | Vibrio phage BONAISHI AXH70757.1 | 7.61 × 10−7 | 51.18 | |||||||
| ORF 22 | Hypothetical protein | 21,889 | 22,572 | 684 | Forward | Vibrio phage BONAISHI AXH70758.1 | 3.23 × 10−23 | 49.10 | |||||||
| ORF 25 | Hypothetical protein | 23,638 | 24,297 | 660 | Forward | Vibrio phage BONAISHI AXH70761.1 | 2.49 × 10−42 | 58.30 | |||||||
| ORF 26 | Hypothetical protein | 24,395 | 25,489 | 1095 | Forward | Vibrio phage BONAISHI AXH70762.1 | 1.87 × 10−9 | 40.40 | |||||||
| ORF 27 | Hypothetical protein | 25,529 | 27,553 | 2025 | Reverse | Vibrio phage BONAISHI AXH70763.1 | 4.37 × 10−162 | 59.50 | |||||||
| ORF 28 | Glycoside hydrolase | 27,596 | 33,355 | 5760 | Reverse | Vibrio phage BONAISHI AXH70764.1 | 6.29 × 10−81 | 52.30 | pfam01551|Peptidase family M23 | 1.40 × 10−35 | 4RNZ_A|Helicobacter pylori | 4.70 × 10−15 | 99.57 | ||
| ORF 29 | DNA-directed RNA polymerase subunit alpha | 33,458 | 34,960 | 1503 | Forward | Vibrio phage BONAISHI AXH70765.1 | 1.65 × 10−173 | 74.10 | 5ZX3_D|Mycobacterium tuberculosis | 2.00 × 10−49 | 100.00 | ||||
| ORF 30 | HNH endonuclease | 35,019 | 35,972 | 954 | Forward | Chryseobacterium gleum] WP_002984461.1 | 5.39 × 10−13 | 57.30 | F: GO:0004519 | IPR010896 | pfam13392|HNH endonuclease | 1.45 × 10−10 | 1U3E_M|Bacillus phage SPO1 | 4.90 × 10−29 | 99.97 |
| ORF 33 | Hypothetical protein | 36,322 | 36,495 | 174 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96130.1 | 6.44 × 10−10 | 70.90 | |||||||
| ORF 34 | RNA-polymerase beta subunit | 36,504 | 37,889 | 1386 | Forward | Vibrio phage BONAISHI AXH70766.1 | 7.28 × 10−71 | 55.40 | |||||||
| ORF 35 | Homing endonuclease | 37,939 | 39,027 | 1089 | Forward | Vibrio phage vB_VmeM-Yong XC32 QAX96446.1 | 2.5 × 10−101 | 61.50 | 3R3P_A|Bacillus phage 0305phi8-36 | 1.70 × 10−3 | 97.46 | ||||
| ORF 38 | DNA-directed RNA polymerase subunit alpha | 39,437 | 42,001 | 2565 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96126.1 | 0.0 × 100 | 64.00 | F: GO:0003899 | SSF64484 | 6J9E_C|Xanthomonas oryzae (strain PXO99A) | 1.50 × 10−77 | 100.00 | ||
| ORF 40 | Hypothetical protein | 42,278 | 43,774 | 1497 | Forward | Vibrio phage BONAISHI AXH70769.1 | 4.42 × 10−38 | 46.31 | |||||||
| ORF 41 | Hypothetical protein | 43,830 | 44,558 | 729 | Forward | Vibrio phage BONAISHI AXH70770.1 | 1.11 × 10−65 | 71.50 | |||||||
| ORF 42 | Prohead core protein protease | 44,567 | 45,388 | 822 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96122.1 | 1.91 × 10−37 | 50.40 | 5JBL_E|Bacteriophage T4 | 2.20 × 10−25 | 99.93 | ||||
| ORF 51 | Tail-tube protein | 48,267 | 49,133 | 867 | Reverse | Vibrio phage vB_VmeM-Yong XC31 QAX96106.1 | 4.32 × 10−61 | 59.70 | 5IV5_DE|Bacteriophage T4 | 5.80 × 10−2 | 96.67 | ||||
| ORF 52 | Tail sheath protein | 49,188 | 51,218 | 2031 | Reverse | Vibrio phage BONAISHI AXH70778.1 | 6.24 × 10−162 | 59.70 | 3SPE_A|Pseudomonas phage phiKZ | 2.40 × 10−69 | 100.00 | ||||
| ORF 54 | Hypothetical protein | 51,437 | 52,378 | 942 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96104.1 | 8.14 × 10−19 | 48.10 | |||||||
| ORF 55 | Virion structural protein | 52,341 | 54,992 | 2652 | Forward | Vibrio phage BONAISHI AXH70780.1 | 1.42 × 10−101 | 48.90 | |||||||
| ORF 56 | Hypothetical protein | 55,004 | 56,665 | 1662 | Forward | Vibrio phage BONAISHI AXH70781.1 | 1.49 × 10−120 | 58.20 | |||||||
| ORF 57 | Phage DNA helicase or terminase, large subunit | 56,711 | 58,900 | 2190 | Forward | Vibrio phage BONAISHI AXH70782.1 | 0.0 × 100 | 69.50 | 3CPE_A|Bacteriophage T4 | 2.60 × 10−34 | 100.00 | ||||
| ORF 58 | Hypothetical protein | 58,943 | 60,159 | 1217 | Reverse | Vibrio phage BONAISHI AXH70784.1 | 3.34 × 10−110 | 65.90 | |||||||
| ORF 60 | Hypothetical protein | 60,314 | 61,015 | 702 | Forward | Vibrio phage BONAISHI AXH70786.1 | 1.02 × 10−8 | 48.60 | |||||||
| ORF 62 | Hypothetical protein | 61,415 | 62,446 | 1032 | Forward | Vibrio phage BONAISHI AXH70788.1 | 2.60 × 10−46 | 60.50 | |||||||
| ORF 64 | RNA-binding protein | 62,625 | 64,175 | 1551 | Reverse | Vibrio phage BONAISHI AXH70790.1 | 6.79 × 10−5 | 44.30 | F: GO:0003723 | IPR037214 | 2NVO_A|Deinococcus radiodurans | 1.60 × 10−62 | 100.00 | ||
| ORF 90 | Hypothetical protein | 69,563 | 70,327 | 765 | Reverse | Vibrio phage BONAISHI AXH70799.1 | 5.98 × 10−37 | 58.00 | |||||||
| ORF 99 | Hypothetical protein | 74,169 | 74,585 | 417 | Forward | Vibrio phage BONAISHI AXH70809.1 | 2.55 × 10−10 | 51.20 | |||||||
| ORF 101 | DNA polymerase | 74,767 | 76,938 | 2172 | Forward | Vibrio phage BONAISHI AXH70810.1 | 0.0 × 100 | 64.80 | F: GO:0000166 | IPR006134 | cl33389|DNA polymerase type-B family | 4.62 × 10−3 | 3QEX_A|Bacteriophage RB69 | 2.60 × 10−28 | 99.97 |
| ORF 102 | Hypothetical protein | 76,983 | 78,047 | 1065 | Reverse | Vibrio phage BONAISHI AXH70811.1 | 1.42 × 10−36 | 52.10 | |||||||
| ORF 105 | Hypothetical protein | 78,386 | 80,323 | 1938 | Forward | Vibrio phage BONAISHI AXH70812.1 | 7.30 × 10−68 | 47.20 | |||||||
| ORF 106 | DNA-directed RNA polymerase | 80,416 | 81,771 | 1356 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96059.1 | 8.5 × 10−130 | 67.60 | F: GO:0003677 | SSF64484 | cl37096|DNA-directed RNA polymerase, beta subunit | 1.24 × 10−3 | 6RFL_A|Vaccinia virus GLV-1h68 | 1.30 × 10−36 | 100.00 |
| ORF 107 | HNH endonuclease | 81,598 | 82,680 | 1083 | Forward | Pseudomonas sp. JY-Q WP_064614171.1 | 1.0 × 10−9 | 45.21 | |||||||
| ORF 109 | RNA polymerase beta prime subunit | 82,888 | 83,265 | 378 | Forward | Vibrio phage BONAISHI AXH70813.1 | 6.86 × 10−8 | 53.10 | |||||||
| ORF 112 | Hypothetical protein | 84,239 | 84,934 | 606 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96047.1 | 2.32 × 10−17 | 49.00 | |||||||
| ORF 113 | Hypothetical protein | 84,968 | 85,369 | 402 | Forward | Vibrio phage BONAISHI AXH70815.1 | 5.83 × 10−20 | 54.50 | |||||||
| ORF 114 | Hypothetical protein | 85,371 | 85,847 | 477 | Forward | Vibrio phage BONAISHI AXH70816.1 | 3.08 × 10−18 | 59.20 | |||||||
| ORF 115 | Putative nuclease SbcD subunit D | 85,759 | 86,961 | 1203 | Forward | Vibrio phage BONAISHI AXH70817.1 | 1.88 × 10−74 | 56.10 | F: GO:0016787 | IPR029052 | cl33866|DNA repair exonuclease SbcCD nuclease subunit | 1.92 × 10−9 | 6S6V_B|Escherichia coli | 4.30 × 10−30 | 100.00 |
| ORF 116 | Hypothetical protein | 86,958 | 87,755 | 798 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96043.1 | 6.97 × 10−44 | 55.00 | |||||||
| ORF 117 | Hypothetical protein | 87,818 | 88,516 | 699 | Forward | Vibrio phage BONAISHI AXH70819.1 | 7.16 × 10−35 | 55.40 | |||||||
| ORF 118 | Hypothetical protein | 88,692 | 90,155 | 1464 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96041.1 | 2.29 × 10−68 | 52.10 | |||||||
| ORF 119 | Hypothetical protein | 90,170 | 91,627 | 1458 | Forward | Vibrio phage BONAISHI AXH70821.1 | 1.49 × 10−67 | 50.00 | |||||||
| ORF 120 | Hypothetical protein | 91,671 | 93,587 | 1917 | Forward | Vibrio phage BONAISHI AXH70822.1 | 7.01 × 10−27 | 49.00 | |||||||
| ORF 122 | RNA polymerase beta subunit | 93,898 | 96,105 | 2208 | Forward | Vibrio phage BONAISHI AXH70824.1 | 1.04 × 10−152 | 55.90 | F: GO:0003677 | IPR007120 | cl37028|DNA-directed RNA polymerase, beta subunit. | 2.01 × 10−8 | 6PST_I|Escherichia coli | 1.00 × 1079 | 100.00 |
| ORF 123 | RNA polymerase beta subunit | 96,116 | 98,080 | 1965 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96036.1 | 0.00 × 100 | 60.40 | F: GO:0003899 | 6PST_J|Escherichia coli | 1.30 × 10−35 | 100.00 | |||
| ORF 125 | ATP-dependent DNA helicase uvsW | 98,305 | 99,693 | 1389 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96032.1 | 1.91 × 10−153 | 68.70 | cl34083|Superfamily II DNA or RNA helicase | 1.64 × 10−12 | 2OCA_A| Bacteriophage T4 | 9.20 × 10−32 | 100.00 | ||
| ORF 127 | ATP-dependent Clp protease proteolytic subunit | 100,236 | 100,730 | 495 | Forward | Vibrio phage BONAISHI AXH70828.1 | 9.08 × 10−26 | 54.00 | F: GO:0004252 | IPR001907 | cl23717|Crotonase/Enoyl-Coenzyme A (CoA) hydratase superfamily | 5.81 × 10−25 | 2FZS_H|Escherichia coli | 1.20 × 10−22 | 99.93 |
| ORF 129 | Hypothetical protein | 101,203 | 102,672 | 1470 | Reverse | Vibrio phage BONAISHI AXH70830.1 | 1.17 × 10−14 | 44.60 | |||||||
| ORF 132 | Hypothetical protein | 103,126 | 103,947 | 822 | Forward | Vibrio phage BONAISHI AXH70832.1 | 7.46 × 10−41 | 55.60 | |||||||
| ORF 133 | RNA polymerase beta prime subunit | 103,982 | 105,211 | 1230 | Forward | Vibrio phage BONAISHI AXH70833.1 | 2.45 × 10−123 | 66.00 | F: GO:0003899 | cl32391|DNA-directed RNA polymerase subunit beta | 1.46 × 10−8 | 6PST_J|Escherichia coli | 7.20 × 10−46 | 100.00 | |
| ORF 134 | Putative replication protein A family | 105,237 | 105,911 | 675 | cl09930|Replication protein A, class 2b aminoacyl-tRNA synthetases | ||||||||||
| ORF 137 | Hypothetical protein | 106,600 | 107,946 | 1347 | Forward | Vibrio phage BONAISHI AXH70836.1 | 5.35 × 10−5 | 45.90 | |||||||
| ORF 138 | DNA polymerase | 107,997 | 109,724 | 1728 | Reverse | Vibrio phage BONAISHI AXH70837.1 | 0.00 × 100 | 74.50 | F: GO:0003676 | IPR036397 | smart00486|DNA polymerase type-B family | 5.325 × 10−8 | 3QEX_A|Bacteriophage RB69 | 6.40 × 10−44 | 100.00 |
| ORF 139 | Virion structural protein | 109,804 | 111,066 | 1263 | Forward | Vibrio phage BONAISHI AXH70838.1 | 1.04 × 10−36 | 48.90 | F: GO:0004222 | 6AIT_C|Escherichia coli (strain K12) | 3.30 × 10−6 | 99.05 | |||
| ORF 140 | Hypothetical protein | 111,070 | 113,046 | 1977 | Forward | Vibrio phage BONAISHI AXH70839.1 | 4.21 × 10−9 | 42.80 | |||||||
| ORF 141 | Virion structural protein | 113,085 | 115,931 | 2847 | Reverse | Vibrio phage BONAISHI AXH70840.1 | 1.42 × 10−178 | 56.70 | |||||||
| ORF 142 | Virion structural protein | 115,924 | 116,970 | 1047 | Reverse | Vibrio phage BONAISHI AXH70841.1 | 1.32 × 10−95 | 60.60 | |||||||
| ORF 143 | Capsid protein | 117,030 | 118,136 | 1107 | Forward | Vibrio phage BONAISHI AXH70842.1 | 1.63 × 10−68 | 58.60 | |||||||
| ORF 144 | Virion structural protein | 118,147 | 119,028 | 882 | Forward | Vibrio phage BONAISHI AXH70843.1 | 7.55 × 10−24 | 50.00 | |||||||
| ORF 146 | Hypothetical protein | 119,612 | 120,913 | 1302 | Forward | Vibrio phage BONAISHI AXH70845.1 | 2.53 × 10−6 | 43.20 | |||||||
| ORF 148 | Putative internal head protein | 122,313 | 123,377 | 1245 | Forward | cl20461|phiKZ-like phage internal head proteins | 1.32 × 10−3 | . | |||||||
| ORF 149 | Hypothetical protein | 123,449 | 125,053 | 1605 | Forward | Vibrio phage BONAISHI AXH70848.1 | 2.17 × 10−108 | 56.10 | |||||||
| ORF 150 | Virion structural protein | 125,053 | 126,270 | 1218 | Forward | Vibrio phage BONAISHI AXH70849.1 | 1.52 × 10−65 | 55.20 | |||||||
| ORF 152 | Virion structural protein | 126,970 | 128,334 | 1365 | Forward | Vibrio phage BONAISHI AXH70851.1 | 1.52 × 10−46 | 50.80 | |||||||
| ORF 153 | DNA helicase | 128,374 | 129,921 | 1548 | Reverse | Vibrio phage BONAISHI AXH70852.1 | 2.72 × 10−177 | 70.10 | F: GO:0003678 | 6BBM_A|Escherichia coli O111 | 1.50 × 10−36 | 100.00 | |||
| ORF 156 | Major capsid protein | 130,604 | 132,769 | 2166 | Forward | Vibrio phage BONAISHI AXH70854.1 | 3.91 × 10−147 | 63.10 | |||||||
| ORF 159 | Hypothetical protein | 134,018 | 135,589 | 1572 | Forward | Vibrio phage BONAISHI AXH70856.1 | 9.35 × 10−141 | 61.50 | |||||||
| ORF 162 | DUF723 domain-containing protein | 138,774 | 139,013 | 240 | Forward | Vibrio mediterranei WP_096444327.1 | 5.15 × 10−12 | 61.90 | F: GO:0004519 | ||||||
| ORF 163 | Endonuclease | 139,080 | 139,541 | 462 | Forward | Vibrio phage 1.225.O._10N.261.48.B7 AUR96455.1 | 2.54 × 10−7 | 46.00 | 6SEI_A|Thielavia terrestris | 1.80 × 10−4 | 97.77 | ||||
| ORF 164 | Holliday junction resolvase | 139,578 | 140,171 | 594 | Reverse | Vibrio phage BONAISHI AXH70859.1 | 8.65 × 10−41 | 62.90 | P: GO:0009987 | IPR036397 | cl21482|Crossover junction endodeoxyribonuclease RuvC and similar proteins | 1.05 × 10−3 | 6LW3_B|Escherichia coli | 4.70 × 10−22 | 99.91 |
| ORF 165 | Virion structural protein | 140,171 | 141,001 | 831 | Reverse | Vibrio phage BONAISHI AXH70860.1 | 2.26 × 10−69 | 60.40 | |||||||
| ORF 166 | Virion structural protein | 141,013 | 143,070 | 2058 | Reverse | Vibrio phage BONAISHI AXH70861.1 | 1.93 × 10−125 | 56.50 | |||||||
| ORF 167 | Putative portal protein | 143,175 | 145,940 | 2766 | Forward | Vibrio phage BONAISHI AXH70862.1 | 3.34 × 10−132 | 55.60 | cl27451|Hypothetical protein | 3.55 × 10−6 | 3JA7_I|Bacteriophage T4 | 2.60 × 10−5 | 98.45 | ||
| ORF 168 | Putative hydrolase | 145,952 | 146,905 | 954 | Forward | Vibrio phage BONAISHI AXH70863.1 | 9.39 × 10−31 | 50.20 | C: GO:0016020 | 4F55_A|Bacillus cereus | 1.10 × 10−24 | 99.92 | |||
| ORF 177 | Hypothetical protein | 150,443 | 151,204 | 762 | Forward | Vibrio phage vB_VhaS-a ANO57550.1 | 1.74 × 10−30 | 57.50 | |||||||
| ORF 178 | Hypothetical protein | 151,281 | 151,952 | 672 | Forward | Vibrio phage vB_VhaS-a ANO57549.1 | 9.91 × 10−40 | 61.50 | |||||||
| ORF 182 | Glycohydrolase | 154,582 | 155,496 | 915 | Forward | Vibrio phage BONAISHI AXH70879.1 | 2.61 × 10−62 | 54.81 | F: GO:0016787 | IPR002477 | |||||
| ORF 183 | Transcription factor: type II DNA-Binding | 155,609 | 156,363 | 855 | 1WTU_A|Bacillus phage SPO1 | 6.60 × 10−3 | 97.04 | ||||||||
| ORF 184 | Transcription factor: type II DNA-Binding | 156,379 | 156,868 | 510 | 1WTU_A|Bacillus phage SPO1 | 6.60 × 10−3 | 97.04 | ||||||||
| ORF 185 | Hypothetical protein | 156,896 | 157,618 | 723 | Forward | Vibrio phage BONAISHI AXH70881.1 | 6.55 × 10−12 | 47.83 | |||||||
| ORF 193 | Putative DNA repair exonuclease | 161,416 | 162,201 | 786 | Forward | Vibrio phage pVa-21 AQT28114.1 | 1.92 × 10−20 | 53.60 | F: GO:0004527 | IPR036412 | 5UJ0_A|Bacteriophage T4 | 7.70 × 10−11 | 99.26 | ||
| ORF 194 | Nucleotide binding protein | 162,149 | 162,760 | 612 | Forward | Vibrio phage BONAISHI AXH70900.1 | 9.48 × 10−16 | 54.40 | cl17018|Fanconi anemia ID complex proteins FANCI and FANCD2 | 7.57 × 10−3 | 3GH1_B|Vibrio cholerae O1 biovar El Tor str. N16961 | 5.20 × 10−7 | 98.66 | ||
| ORF 196 | Hypothetical protein | 163,750 | 164,421 | 672 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96244.1 | 2.14 × 10−100 | 81.60 | |||||||
| ORF 197 | Hypothetical protein | 164,476 | 164,808 | 333 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96240.1| | 1.00 × 10−3 | 38.89 | |||||||
| ORF 198 | DEAD-like helicase | 164,871 | 166,841 | 1971 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96243.1 | 1.70 × 10−166 | 60.90 | P:GO:0000733 | IPR014001 | cd18793|C-terminal helicase domain of the SNF family helicases. | 1.80 × 10−16 | 2OCA_A|Bacteriophage T4 | 1.10 × 10−29 | 99.98 |
| ORF 199 | EAR-like protein | 166,948 | 168,591 | 1647 | IPR009039 | ||||||||||
| ORF 200 | Putative Palindromic Amphipathic Repeat Coding Elements (PARCEL) | 168,504 | 187,867 | 19,254 | IPR011889 | pfam03382|Mycoplasma protein of unknown function, DUF285 | 1.29 × 10−44 | ||||||||
| ORF 202 | DNA-packaging protein: hydrolase | 188,001 | 188,966 | 966 | Forward | Pseudomonas virus phiKZ NP_803591.1 | 5.69 × 10−20 | 53.10 | C: GO:0016020 | 2O0J_A|Bacteriophage T4 | 1.80 × 10−24 | 99.92 | |||
| ORF 207 | UV-endonuclease | 191,809 | 192,780 | 972 | Forward | Vibrio phage 1.084.O._10N.261.49.F5 AUR86431.1 | 6.23 × 10−67 | 55.90 | F: GO:0004519 | IPR004601 | cl23721|AP endonuclease family 2 | 2.84 × 10−48 | 3TC3_B|Sulfolobus acidocaldarius | 1.20 × 10−26 | 99.96 |
| ORF 209 | Dihydrofolate reductase (EC 1.5.1.3) CDS | 193,421 | 194,152 | 732 | Forward | Meiothermus ruber HFG20084.1 | 2.1 × 10−22 | 59.30 | P: GO:0008152 | IPR001796 | cd00209|Dihydrofolate reductase (DHFR) | 9.07 × 10−39 | 1JUV_A|Bacteriophage T4 | 1.50 × 10−20 | 99.87 |
| ORF 223 | Putative nucleotidyl transferase | 202,749 | 203,570 | 822 | Forward | Yersinia phage phiR1-37 YP_004934311.1 | 3.51 × 10−13 | 51.80 | F: GO:0016740 | IPR043519 | cl35051|elongation factor Tu | 3.27 × 10−4 | 2FCL_A|Thermotoga maritima | 6.70 × 10−12 | 99.44 |
| ORF 225 | Putative N-acetyltransferase | 204,006 | 204,428 | 423 | IPR016181 | 5Z6N_A| Escherichia coli (strain K12) | |||||||||
| ORF 229 | Putative HTH-type transcription I regulator MqsA | 205,857 | 506,309 | 453 | 3GA8_A|Escherichia coli K-12 | ||||||||||
| ORF 230 | Hypothetical protein VPIG_00040 | 206,325 | 206,852 | 528 | Forward | Vibrio phage PWH3a-P1 YP_007675900.1 | 5.86 × 10−9 | 56.40 | |||||||
| ORF 231 | Hypothetical protein SAMN05421742_1266 | 206,849 | 207,121 | 453 | Forward | Roseospirillum parvum SDH93001.1| | 1.94 × 10−5 | 54.70 | |||||||
| ORF 243 | Putative glycosylhydrolase | 214,545 | 215,822 | 1278 | IPR013320 | ||||||||||
| ORF 245 | Hypothetical protein BCS93_11070 | 217,217 | 218,539 | 1323 | Reverse | Vibrio breoganii PMP10208.1 | 8.63 × 10−13 | 59.50 | 3ZYP_A|Hypocrea jecorina | 6.00 × 10−3 | 97.33 | ||||
| ORF 247 | Hypothetical protein | 218,552 | 219,949 | 1398 | Reverse | Vibrio breoganii WP_133150968.1 | 1.32 × 10−10 | 42.50 | 3ZYP_A|Hypocrea jecorina | 8.80 × 10−4 | 97.72 | ||||
| ORF 248 | Hypothetical protein NVP1169O_83 | 221,333 | 222,682 | 1350 | Reverse | Vibrio phage 1.169.O._10N.261.52.B1 AUR92111.1 | 5.68 × 10−10 | 40.40 | |||||||
| ORF 249 | Hypothetical protein BDU10_8600 | 222,682 | 223,677 | 996 | Reverse | Burkholderia sp. CF145 OYD65949.1 | 9.15 × 10−15 | 53.10 | |||||||
| ORF 250 | Polynucleotide kinase | 223,801 | 224,589 | 789 | Forward | Aeromonas virus Aeh1 NP_943967.1 | 1.30 × 10−11 | 37.40 | cl40282|HAD domain in Swiss Army Knife RNA repair proteins. | 1.12 × 10−8 | 5UJ0_A|Bacteriophage T4 | 1.80 × 10−4 | 98.02 | ||
| ORF 256 | SH3 protein | 228,286 | 228,981 | 695 | cl17036|Src Homology 3 domain superfamily | 6.19 × 10−3 | |||||||||
| ORF 262 | Phosphagen kinase | 232,254 | 232,973 | 720 | Forward | cl02823|Phosphagen (guanidino) kinases | |||||||||
| ORF 263 | DNA polymerase accessory protein 44: AAA+, ATP hydrolase | 233,047 | 234,390 | 1344 | Forward | Salmonella enterica EAZ2022740.1 | 2.56 × 10−30 | 50.70 | F: GO:0000166 | IPR003593 | 3U61_D|Bacteriophage T4} | 9.00 × 10−12 | 99.45 | ||
| ORF 267 | Ribonuclease E/G | 236,230 | 236,958 | 729 | cl29166|Ribonuclease E/G family | 1.32 × 10−3 | |||||||||
| ORF 272 | DNA polymerase II large subunit | 239,038 | 239,373 | 336 | cl36419|DNA-directed DNA polymerase II large subunit | 3.90 × 10−3 | |||||||||
| ORF 275 | NAD-dependent DNA ligase LigA | 240,880 | 242,817 | 1938 | Forward | Salinivibrio sp. ES.052 WP_074213176.1 | 2.87 × 10−131 | 57.20 | P: GO:0006259 | IPR001357 | cl35633|NAD-dependent DNA ligase LigA; Validated | 0.00 × 100 | 5TT5_A|Escherichia coli K12 | 1.50 × 10−121 | 100.00 |
| ORF 278 | GTP cyclohydrolase II | 244,297 | 244,800 | 504 | Forward | Vibrio phage PWH3a-P1 YP_007676007.1 | 2.19 × 10−30 | 61.60 | |||||||
| ORF 281 | Thymiylate kinase | 245,594 | 246,316 | 723 | Forward | Firmicutes bacterium CAG:582 CDB28696.1 | 3.93 × 10−34 | 54.80 | P: GO:0006796. | IPR039430 | cl17190|Nucleoside/nucleotide kinase (NK). | 1.10 × 10−26 | 3LV8_A|Vibrio cholerae O1 biovar El Tor | 2.50 × 10−22 | 99.92 |
| ORF 283 | Ribonucleotide reductase of class Ia (aerobic), alpha subunit | 246,875 | 249,163 | 2289 | Forward | Rodentibacter pneumotropicus WP_077664105.1 | 0.00 × 100 | 71.30 | F: GO:0000166 | IPR005144 | cl32350|ribonucleoside-diphosphate reductase subunit alpha. | 0.00 × 100 | 2XAP_A|Escherichia coli | 5.10 × 10−23 | 100.00 |
| ORF 286 | Ribonucleotide reductase of class Ia (aerobic), beta subunit | 250,213 | 251,331 | 1119 | Forward | Sulfurivirga caldicuralii WP_074201546.1 | 8.00 × 10−139 | 69.40 | F: GO:0004748 | cl00264|Ferritin-like superfamily of diiron-containing four-helix-bundle proteins | 0.00 × 100 | 1MXR_B|Escherichia coli | 1.90 × 10−56 | 100.00 | |
| ORF 287 | BspA family leucine-rich repeat surface protein | 251,408 | 255,091 | 3684 | Forward | Helicobacter bizzozeronii WP_158656920.1 | 1.25 × 10−36 | 46.00 | C: GO:0016020 | IPR005046 | cl37689|Mycoplasma protein of unknown function, DUF285 | 2.51 × 10−27 | |||
| ORF 299 | PIN terminus | 261,891 | 262,838 | 978 | IPR002716 | cl28905|PIN (PilT N terminus) domain: Superfamily | 2.33 × 10−6 | 2HWY_A|Homo sapiens | 4.10 × 10−3 | 96.35 | |||||
| ORF 308 | Hypothetical protein | 266,214 | 266,954 | 741 | Forward | Cellulomonas aerilata WP_146903668.1 | 2.66 × 10−4 | 41.20 | 6HIY_DS|Trypanosoma brucei brucei | 5.50 × 10−11 | 99.13 | ||||
| ORF 310 | Glutaredoxin | 267,350 | 268,000 | 651 | cl35908|glutaredoxin 2 | 2.14 × 10−3 | |||||||||
| ORF 316 | Asp/Glu/Hydantoin racemase | 270,693 | 271,877 | 1185 | cl00518|Asp/Glu/Hydantoin racemase | 5.48 × 10−3 | |||||||||
| ORF 319 | RNA-binding protein | 274,073 | 274,750 | 678 | Forward | Vibrio phage BONAISHI AXH70995.1 | 4.95 × 10−20 | 51.30 | F: GO:0016787 | IPR036397 | cl10012|DnaQ-like (or DEDD) 3′-5′ exonuclease domain superfamily | 1.01 × 10−13 | 6N6A_A|Vibrio cholerae | 1.60 × 10−15 | 99.70 |
| ORF 323 | Thymidylate synthase (EC 2.1.1.45) | 276,131 | 277,057 | 927 | Forward | Vibrio phage 2.275.O._10N.286.54.E11 AUS02985.1 | 3.26 × 10−76 | 62.60 | P: GO:0008152 | IPR023451 | cl19097|Thymidylate synthase and pyrimidine hydroxy methylase. | 1TIS_A|Bacteriophage T4 | |||
| ORF 326 | Nucleoside Triphosphate Pyrophosphohydrolase | 277,926 | 278,654 | 729 | Forward | Vibrio phage vB_VmeM-Yong XC31 QAX96185.1 | 4.77 × 10−10 | 58.40 | cl16941|Nucleoside Triphosphate Pyrophosphohydrolase (EC 3.6.8) MazG-like domain superfamily | 4.07 × 10−3 | 2YF4_B|Deinococcus radiodurans | 8.80 × 10−20 | 99.83 | ||
| ORF 329 | Hypothetical protein | 279,832 | 280,650 | 819 | Forward | IPR006530 | |||||||||
| ORF 330 | Hypothetical protein | 280,703 | 281,116 | 414 | Reverse | Vibrio phage BONAISHI AXH71034.1 | 1.91 × 10−13 | 54.40 | |||||||
| ORF 331 | Hypothetical protein yiiX | 281,272 | 281,703 | 432 | Reverse | Vibrio phage vB_VmeM-Yong XC31 QAX96165.1 | 2.49 × 10−27 | 55.60 | IPR038765 | cl21534|NlpC/P60 family. | 4.98 × 10−3 | 2IF6_A|Escherichia coli | 4.10 × 10−21 | 99.87 | |
| ORF 332 | Hypothetical protein | 281,713 | 282,711 | 999 | Reverse | Vibrio phage BONAISHI AXH71036.1 | 1.57 × 10−41 | 47.40 |
| Bacterial Strains | Method of Identification | Location |
|---|---|---|
| Vibrio harveyi | ||
| DSM19623 | ENA Accession No: BAOD01000001 | USA |
| SNGR | BIOLOG GENiii | Greece |
| KS6 | BIOLOG GENiii | Greece |
| Vh2 | BIOLOG GENiii, toxR (+) | Greece |
| Vh5 | BIOLOG GENiii, toxR (+) | Greece |
| VhSernFr | BIOLOG GENiii, toxR (+) | Greece |
| VhP1 Liv | toxR (+) | Greece |
| VhP1 Spl | toxR (+) | Greece |
| VhKarx | BIOLOG GENiii, toxR (+) | Greece |
| RG1 | toxR (+) | Greece |
| Barb A4/1.1 | BIOLOG GENiii, toxR (+) | Greece |
| SerKid | BIOLOG GENiii | Greece |
| SerKid2 | BIOLOG GENiii | Greece |
| SerSd | BIOLOG GENiii | Greece |
| SA 5.1 | 16S rRNA | KSA |
| SA 6.1 | 16S rRNA | KSA |
| SA 9.2 | 16S rRNA | KSA |
| SA 1.2 | 16S rRNA | KSA |
| SA 7.1 | 16S rRNA | KSA |
| SA 3.1 | 16S rRNA | KSA |
| SA 4.1 | 16S rRNA | KSA |
| SA 2.1 | 16S rRNA | KSA |
| Epith. D | BIOLOG GENiii | Greece |
| Vh No. 22 | BIOLOG GENiii | Greece |
| Vh6 | BIOLOG GENiii | Greece |
| Vibrio alginolyticus | ||
| V1 | BIOLOG GENiii | Greece |
| V2 | BIOLOG GENiii | Greece |
| HCMR 1 Art. 3 | Clinical strain | Greece |
| DSM 2171 | ENA Accession No.: AB372523 | Japan |
| Valg HCMR | BIOLOG GENiii | Greece |
| Skironis-2 | BIOLOG GENiii | Greece |
| NS A6 | BIOLOG GENiii | Greece |
| Rot. Vib. 5 | BIOLOG GENiii | Greece |
| Vibrio anguillarum | ||
| 90-11-286 | Clinical strain | Denmark |
| VIB 44 | Clinical strain | Italy |
| VIB 64 | Clinical strain | Spain |
| VIB 243 | Clinical strain | USA |
| Vibrio campbellii | ||
| VIB391 | NCBI RefSeq No: GCF_002078065.1 | Thailand |
| Vibrio owensii | ||
| SA 1.1 | 16S rRNA | KSA |
| SA 9.1 | 16S rRNA | KSA |
| Vibrio parahaemolyticus | ||
| VPINH | BIOLOG GENiii | Greece |
| Vibrio splendidus | ||
| Barb A4/2 | BIOLOG GENiii | Greece |
| VaAn | Clinical strain | Greece |
| Other Vibrio spp. | ||
| Art. 1 | TCBS | Greece |
| Art. 2 | TCBS | Greece |
| Rot. 2 | toxR (+) | Greece |
| Barb A4/1.2 | TCBS | Greece |
| Rot. Vib. 1 | TCBS | Greece |
| Rot. Vib. 2 | TCBS | Greece |
| Rot. Vib. 3 | TCBS | Greece |
| Rot. Vib. 4 | TCBS | Greece |
| Rot. Vib. 6 | TCBS | Greece |
| Antimicrobial Agent | Disk Diffusion (μg) | Zone Diameter (mm) Interpretive Criteria | ||
|---|---|---|---|---|
| S | I | R | ||
| Ampicillin | 10 | ≥17 | 14–16 | ≤13 |
| Tetracycline | 30 | ≥15 | 12–14 | ≤11 |
| Sulphamethoxazole/trimethoprim | 25 | ≥16 | 11–15 | ≤10 |
| Oxytetracycline a | 30 | ≥27 | 17–26 | ≤16 |
| Florfenicol b | 30 | ≥18 | 13–17 | ≤12 |
| Oxalinic acid c | 2 | ≥19 | 14–18 | ≤13 |
| Flumequine c | 30 | ≥19 | 14–18 | ≤13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Misol, G.N., Jr.; Kokkari, C.; Katharios, P. Biological and Genomic Characterization of a Novel Jumbo Bacteriophage, vB_VhaM_pir03 with Broad Host Lytic Activity against Vibrio harveyi. Pathogens 2020, 9, 1051. https://doi.org/10.3390/pathogens9121051
Misol GN Jr., Kokkari C, Katharios P. Biological and Genomic Characterization of a Novel Jumbo Bacteriophage, vB_VhaM_pir03 with Broad Host Lytic Activity against Vibrio harveyi. Pathogens. 2020; 9(12):1051. https://doi.org/10.3390/pathogens9121051
Chicago/Turabian StyleMisol, Gerald N., Jr., Constantina Kokkari, and Pantelis Katharios. 2020. "Biological and Genomic Characterization of a Novel Jumbo Bacteriophage, vB_VhaM_pir03 with Broad Host Lytic Activity against Vibrio harveyi" Pathogens 9, no. 12: 1051. https://doi.org/10.3390/pathogens9121051