Anaplasma phagocytophilum and Babesia Species of Sympatric Roe Deer (Capreolus capreolus), Fallow Deer (Dama dama), Sika Deer (Cervus nippon) and Red Deer (Cervus elaphus) in Germany


Abstract
:1. Introduction
2. Results
2.1. Anaplasma phagocytophilum
2.2. Babesia Species
- 18 sequences of B. capreoli from roe deer, with 99%–100% similarity to other GenBank® entries for B. capreoli from roe deer from Germany (e.g., accession number KU351826; [8].
- Three sequences of Babesia sp. EU1 from roe deer, with 100% identity to GenBank® entries for Babesia sp. EU1 from I. ricinus from Spain (KM289158), Ixodes persulcatus from Mongolia (LC005775) and reindeer (Rangifer tarandus) from Germany (KM657247).
- Eight sequences of B. odocoilei-like from three sika deer and five red deer (four wild deer, one farmed deer) showing 99–100% similarity to sequences found in fallow deer in Germany (KU351828) and in red deer in Austria (JN543180).
- Eight sequences from two sika deer and six red deer (two wild deer, four farmed deer) with 100% identity to B. divergens from I. ricinus in Poland (KU748896) and red deer in Austria (KX018019). In addition, sequences obtained from one fallow deer and one sika deer were closely related to both B. capreoli and B. divergens but did not allow for discrimination of the two species, and the sequence from a roe deer was short and could only be identified to Babesia genus level.
2.3. Anaplasma phagocytophilum/Babesia Species Co-Infections
3. Discussion
3.1. Anaplasma phagocytophilum
3.2. Anaplasma phagocytophilum Variants
3.3. Babesia Species
3.4. Co-Infections
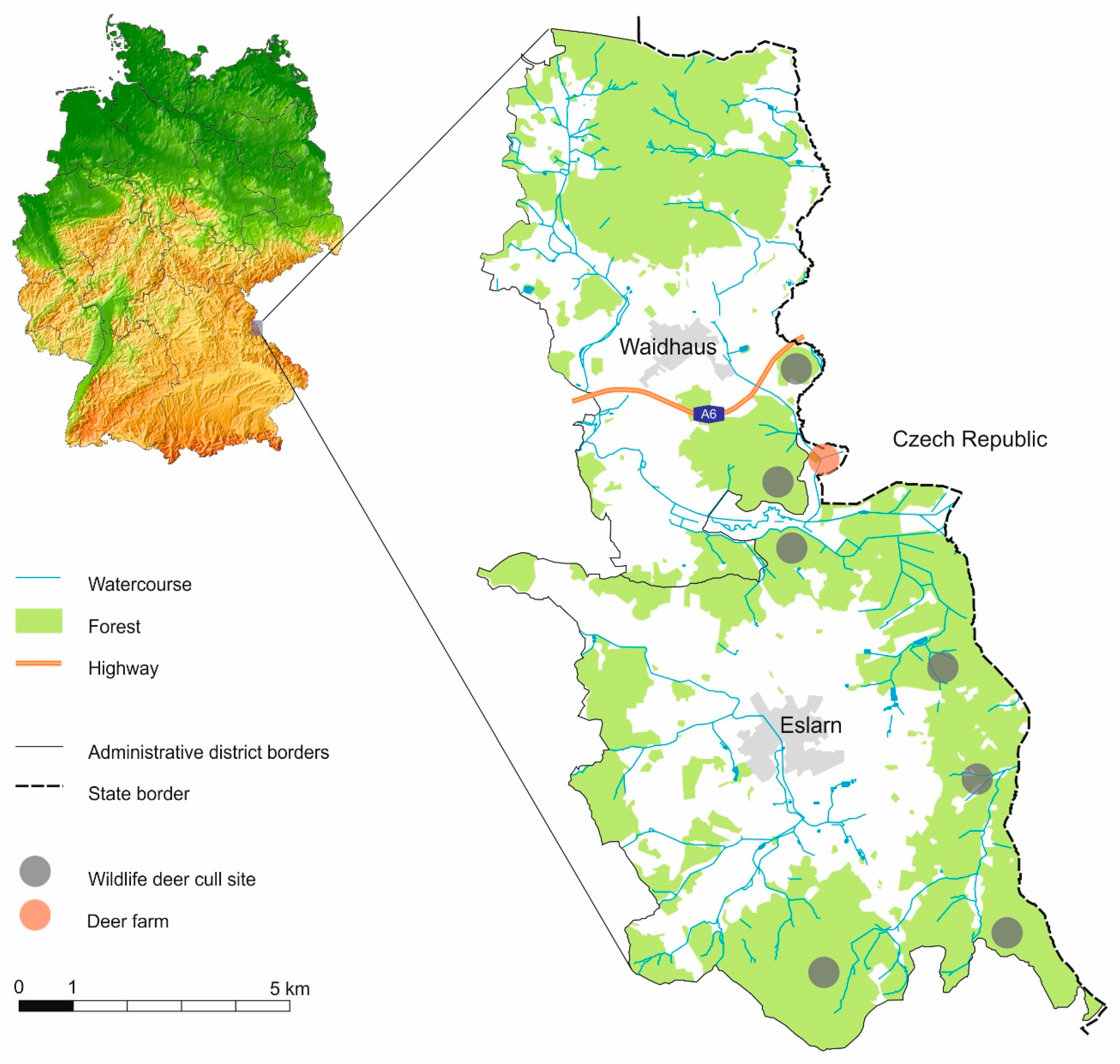
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalská, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef] [PubMed]
- Stuen, S.; Granquist, E.G.; Silaghi, C. Anaplasma phagocytophilum—A widespread multi-host pathogen with highly adaptive strategies. Front. Cell Infect. Microbiol. 2013, 3, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrandt, A.; Gray, J.S.; Hunfeld, K.P. Human babesiosis in Europe: What clinicians need to know. Infection 2013, 41, 1057–1072. [Google Scholar] [CrossRef] [PubMed]
- Häselbarth, K.; Tenter, A.M.; Brade, V.; Krieger, G.; Hunfeld, K.P. First case of human babesiosis in Germany—Clinical presentation and molecular characterisation of the pathogen. Int. J. Med. Microbiol. 2007, 297, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, A.; Hunfeld, K.P.; Baier, M.; Krumbholz, A.; Sachse, S.; Lorenzen, T.; Kiehntopf, M.; Fricke, H.J.; Straube, E. First confirmed autochthonous case of human Babesia microti infection in Europe. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Overzier, E.; Pfister, K.; Herb, I.; Mahling, M.; Bock, G., Jr.; Silaghi, C. Detection of tick-borne pathogens in roe deer (Capreolus capreolus), in questing ticks (Ixodes ricinus), and in ticks infesting roe deer in southern Germany. Ticks Tick Borne Dis. 2013, 4, 320–328. [Google Scholar] [CrossRef]
- Kauffmann, M.; Rehbein, S.; Hamel, D.; Lutz, W.; Heddergott, M.; Pfister, K.; Silaghi, C. Anaplasma phagocytophilum and Babesia spp. in roe deer (Capreolus capreolus), fallow deer (Dama dama) and mouflon (Ovis musimon) in Germany. Mol. Cell Probes 2017, 31, 46–54. [Google Scholar] [CrossRef]
- Enigk, K.; Friedhoff, K. Babesia capreoli n. sp. beim Reh (Capreolus capreolus). Zschr. Tropenmed. Parasitol. 1962, 13, 8–20. (In German) [Google Scholar]
- Duh, D.; Petrovec, M.; Bidovec, A.; Avsic-Zupanc, T. Cervids as babesiae hosts, Slovenia. Emerg. Infect. Dis. 2005, 11, 1121–1123. [Google Scholar] [CrossRef]
- Tampieri, M.P.; Galuppi, R.; Bonoli, C.; Cancrini, G.; Moretti, A.; Pietrobelli, M. Wild ungulates as Babesia hosts in northern and central Italy. Vector Borne Zoonotic Dis. 2008, 8, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Silaghi, C.; Hamel, D.; Pfister, K.; Rehbein, S. Babesia species and co-infection with Anaplasma phagocytophilum in free-ranging ungulates from Tyrol, Austria. Wien. Tierärztl Mschr. 2011, 98, 268–274. [Google Scholar]
- Zintl, A.; Finnerty, E.J.; Murphy, T.M.; de Waal, T.; Gray, J.S. Babesias of red deer (Cervus elaphus) in Ireland. Vet. Res. 2011, 42, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jouglin, M.; Fernández-de-Mera, I.G.; de la Cotte, N.; Ruiz-Fons, F.; Gortázar, C.; Moreau, E.; Bastian, S.; de la Fuente, J.; Malandrin, L. Isolation and characterization of Babesia pecorum sp. nov. from farmed red deer (Cervus elaphus). Vet. Res. 2014, 45, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, A.O.; Mathis, A.; Ryser-Degiorgis, M.P. Babesia spp. in European wild ruminant species: Parasite diversity and risk factors for infection. Vet. Res. 2014, 45, 65. [Google Scholar] [CrossRef] [Green Version]
- Zanet, S.; Trisciuoglio, A.; Bottero, E.; de Mera, I.G.; Gortazar, C.; Carpignano, M.G.; Ferroglio, E. Piroplasmosis in wildlife: Babesia and Theileria affecting free-ranging ungulates and carnivores in the Italian Alps. Parasites Vectors 2014, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Pūraitė, I.; Rosef, O.; Radzijevskaja, J.; Lipatova, I.; Paulauskas, A. The first detection of species of Babesia Starcovici, 1893 in moose, Alces alces (Linnaeus), in Norway. Fol. Parasitol. 2016, 63, 9. [Google Scholar]
- Hoby, S.; Robert, N.; Mathis, A.; Schmid, N.; Meli, M.L.; Hofmann-Lehmann, R.; Lutz, H.; Deplazes, P.; Ryser-Degiorgis, M.-P. Babesiosis in free-ranging chamois (Rupicapra r. rupicapra) from Switzerland. Vet. Parasitol. 2007, 148, 341–345. [Google Scholar] [CrossRef]
- Malandrin, L.; Jouglin, M.; Sun, Y.; Brisseau, N.; Chauvin, A. Redescription of Babesia capreoli (Enigk and Friedhoff, 1962) from roe deer (Capreolus capreolus): Isolation, cultivation, host specificity, molecular characterisation and differentiation from Babesia divergens. Int. J. Parasitol. 2010, 40, 277–284. [Google Scholar] [CrossRef]
- Stuen, S.; Olsson Engvall, E.; van de Pol, I.; Schouls, L.M. Granulocytic ehrlichiosis in a roe deer calf in Norway. J. Wildl. Dis. 2001, 37, 614–616. [Google Scholar] [CrossRef] [Green Version]
- Eick, E. Das Sikawild. Eine Monographie; Cw Nordwest Media Verl.: Grevesmühlen, Germany, 2018. [Google Scholar]
- Gangl, C. Hübsche kleine Japaner im Kommen. Jagd. Bayern. 2013, 11, 20–21. (In German) [Google Scholar]
- Helm, G. Königsklasse Sikajagd. Jagd. Bayern 2013, 11, 22–23. (In German) [Google Scholar]
- Weigl, H. Kleiner Hirsch auf Abwegen I. Jagd. Bayern 2015, 9, 28–30. (In German) [Google Scholar]
- Weigl, H. Kleiner Hirsch auf Abwegen II. Jagd. Bayern 2015, 10, 38–40. (In German) [Google Scholar]
- Dvořák, J.; Palyzová, L. Analysis of the development and spatial distribution of sika deer (Cervus nippon) populations on the territory of the Czech Republic. Acta Univ. Agric. Silvic. Mendel. Brun. 2016, 64, 1507–1515. [Google Scholar] [CrossRef]
- Silaghi, C.; Hamel, D.; Thiel, C.; Pfister, K.; Passos, L.M.; Rehbein, S. Genetic variants of Anaplasma phagocytophilum in wild caprine and cervid ungulates from the Alps in Tyrol, Austria. Vector Borne Zoonotic Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Scharf, W.; Schauer, S.; Freyburger, F.; Petrovec, M.; Schaarschmidt-Kiener, D.; Liebisch, G.; Runge, M.; Ganter, M.; Kehl, A.; Dumler, J.S.; et al. Distinct host species correlate with Anaplasma phagocytophilum ankA gene clusters. J. Clin. Microbiol. 2011, 49, 790–796. [Google Scholar] [CrossRef] [Green Version]
- Hornok, S.; Sugár, L.; Fernández de Mera, I.G.; de la Fuente, J.; Horváth, G.; Kovács, T.; Micsutka, A.; Gönczi, E.; Flaisz, B.; Takács, N.; et al. Tick- and fly-borne bacteria in ungulates: The prevalence of Anaplasma phagocytophilum, haemoplasmas and rickettsiae in water buffalo and deer species in Central Europe, Hungary. BMC Vet. Res. 2018, 14, 98. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.T.; Shaw, S.E.; Morgan, E.R. Anaplasma phagocytophilum infection in a multi-species deer community in the New Forest, England. Eur. J. Wildl Res. 2009, 55, 439–442. [Google Scholar] [CrossRef]
- Ebani, V.V.; Cerri, D.; Fratini, F.; Ampola, M.; Andreani, E. Anaplasma phagocytophilum infection in a fallow deer (Dama dama) population in a preserve of central Italy. New Microbiol. 2007, 30, 161–165. [Google Scholar]
- Rehbein, S.; Visser, M.; Jekel, I.; Silaghi, C. Endoparasites of the fallow deer (Dama dama) of the Antheringer Au in Salzburg, Austria. Wien. Klin. Wochenschr. 2014, 126, S37–S41. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wuritu; Yoshikawa, Y.; Gaowa; Kawamori, F.; Ikegaya, A.; Ohtake, M.; Ohashi, M.; Shimada, M.; Takada, A.; et al. A molecular and serological survey of Rickettsiales bacteria in wild sika deer (Cervus nippon nippon) in Shizuoka Prefecture, Japan: High prevalence of Anaplasma species. Jpn. J. Infect. Dis. 2015, 68, 434–437. [Google Scholar] [CrossRef] [PubMed]
- Ybanez, A.; Matsumoto, K.; Kishimoto, T.; Inokuma, H. Molecular analyses of a potentially novel Anaplasma species closely related to Anaplasma phagocytophilum detected in sika deer (Cervus nippon yesoensis) in Japan. Vet. Microbiol. 2012, 157, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, M.A.M.; Lee, K.; Taylor, K.; Nakao, R.; Sashika, M.; Shimozuru, M.; Tsubota, T. Molecular characterizaton and specific detection of Anaplasma species (AP-sd) in sika deer and its first detection in wild brown bears and rodents in Hokkaiko, Japan. Infect. Genet. Evol 2015, 36, 268–274. [Google Scholar] [CrossRef]
- Hapunik, J.; Víchová, B.; Karbowiak, G.; Wita, I.; Bogdaszewski, M.; Pet’ko, B.; Häselbarth, K. Wild and farm breeding cervids infections with Anaplasma phagocytophilum. Ann. Agric. Environ. Med. 2011, 18, 73–77. [Google Scholar]
- Yang, J.; Liu, Z.; Niu, Q.; Luo, J.; Wang, X.; Yin, H. Molecular detection of Anaplasma phagocytophilum in wild cervids and hares in China. J. Wildl. Dis. 2017, 53, 420–423. [Google Scholar] [CrossRef]
- Silaghi, C.; Kohn, B.; Chirek, A.; Thiel, C.; Nolte, I.; Liebisch, G.; Pfister, K. Relationship of molecular and clinical findings on Anaplasma phagocytophilum involved in natural infections of dogs. J. Clin. Microbiol. 2011, 49, 4413–4414. [Google Scholar] [CrossRef] [Green Version]
- Silaghi, C.; Liebisch, G.; Pfister, K. Genetic variants of Anaplasma phagocytophilum from 14 equine granulocytic anaplasmosis cases. Parasites Vectors 2011, 16, 161. [Google Scholar] [CrossRef] [Green Version]
- Nieder, M.; Silaghi, C.; Hamel, D.; Pfister, K.; Schmäschke, R.; Pfeffer, M. Tick-borne fever caused by Anaplasma phagocytophilum in Germany: First laboratory confirmed case in a dairy cattle herd. Tierärztl. Prax. Ausg. G 2012, 40, 101–106. [Google Scholar] [CrossRef]
- Granquist, E.G.; Bårdsen, K.; Bergström, K.; Stuen, S. Variant-and individual dependent nature of persistent Anaplasma phagocytophilum infection. Acta Vet. Scand. 2010, 52, 25. [Google Scholar] [CrossRef] [Green Version]
- Huhn, C.; Winter, C.; Wolfsperger, T.; Wüppenhorst, N.; Strašek Smrdel, K.; Skuballa, J.; Pfäffle, M.; Petney, T.; Silaghi, C.; Dyachenko, V.; et al. Analysis of the population structure of Anaplasma phagocytophilum using multilocus sequence typing. PLoS ONE 2014, 9, e93725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; van Leeuwen, A.D.; Hengeveld, P.; Heylen, D.; Heyman, P.; van Maanen, C.; Butler, C.M.; Földvári, G.; et al. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasites Vectors 2014, 7, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aardeema, M.L.; von Loewenich, F.D. Varying influences of selection and demography in host-adapted populations of the tick-transmitted bacterium, Anaplasma phagocytophilum. BMC Evol. Biol. 2015, 15, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, S.; Jouglin, M.; L’Hostis, M.; Chauvin, A. Babesia sp. EU1 from roe deer and transmission within Ixodes ricinus. Emerg. Infect. Dis. 2007, 8, 1208–1210. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, E.; Moustafa, M.A.M.; Lee, K.; Mohamed, W.M.A.; Nakao, R.; Shimozuru, M.; Sashika, M.; Younis, E.E.A.; El-Khodery, S.A.; Tsuboa, T. Molecular identification and characterization of piroplasm species in Hokkaido sika deer (Cervus nippon yesoensis), Japan. Ticks Tick BBMorne. Dis. 2017, 8, 802–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Yang, J.; Guan, G.; Liu, A.; Wang, B.; Luo, J.; Yin, H. Molecular detection and identification of piroplasms in sika deer (Cervus nippon) from Jilin province, China. Parasites Vectors 2016, 9, 156. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Khan, M.K.; Zhang, W.-J.; Zhang, Q.-L.; Zhou, Y.-Q.; Hu, M.; Zhao, J. Detection and identification of Theileria infection in sika deer (Cervus nippon) in China. J. Parasitol. 2012, 98, 598–603. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Z.; Zhijie, L.; Liu, J.; Yang, J.; Li, Q.; Li, Y.; Cen, S.; Guan, G.; Ren, Q.; et al. Molecular identification of Theileria parasites of northwestern Chinese Cervidae. Parasites Vectors 2014, 7, 225. [Google Scholar] [CrossRef] [Green Version]
- Zamoto-Niikura, A.; Tsuji, M.; Imaoka, K.; Kimura, M.; Moirkawa, S.; Holman, P.; Hirata, H.; Ishihara, C. Sika deer carrying Babesia parasites closely related B. divergens, Japan. Emerg. Inf. Dis. 2014, 20, 1398–1400. [Google Scholar] [CrossRef]
- Gray, J.S.; Murphy, T.M.; Waldrup, K.A.; Wagner, G.G.; Blewett, D.A.; Harrington, R. Comparative studies of Babesia spp. from white-tailed and sika deer. J. Wildl. Dis. 1991, 27, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Plötz, C.; Rehbein, S.; Bamler, H.; Reindl, H.; Pfister, K.; Scheuerle, M.C. Fascioloides magna epizootiology in a deer farm in Germany. Berl. Münch. Tierärztl. Wochenschr. 2015, 128, 177–182. [Google Scholar] [PubMed]
- Rehbein, S.; Visser, M.; Plötz, C.; Schwarz, L.; Lindner, T.; Bamler, H.; Pfister, K. Endoparasites of red deer (Cervus elaphus) from a deer farm endemic with fascioloidosis in Germany. Berl. Münch. Tierärztl. Wochenschr. 2019, 132, 464–471. [Google Scholar]
- Courtney, J.; Kostelnik, L.M.; Zeidner, N.S.; Massung, R.F. Multiplex real-time PCR for detection of Anaplasma phagocytophilum and Borrelia burgdorferi. J. Clin. Microbiol. 2004, 42, 3164–3168. [Google Scholar] [CrossRef] [Green Version]
- Massung, R.F.; Slater, K.; Owens, J.H.; Nicholson, W.L.; Mather, T.N.; Solberg, V.B.; Olson, J.G. Nested PCR assay for detection of granulocytic ehrlichiae. J. Clin. Microbiol. 1998, 36, 1090–1095. [Google Scholar] [CrossRef] [Green Version]
- Casati, S.; Sager, H.; Gern, L.; Piffaretti, J.C. Presence of potentially pathogenic Babesia sp. for human in Ixodes ricinus in Switzerland. Ann. Agric. Environ. Med. 2006, 13, 65–70. [Google Scholar] [PubMed]


{kind=link}
| Variant | Number of Variants | GenBank® Accession Numbers | ||||
|---|---|---|---|---|---|---|
| Total | Roe Deer | Fallow Deer | Sika Deer | Red Deer | ||
| B | 5 | 0 | 0 | 3 | 2 | KU705180, KU705181, KU705133, KU705136 |
| I | 7 | 6 | 0 | 1 | 0 | KU705140, KU705141, KU705184, KU705152, KU705154, KU705158, KU705160 |
| J | 1 | 0 | 0 | 1 | 0 | KU705187 |
| S | 9 | 0 | 0 | 4 | 5 | KU705178, KU705182, KU705124, KU705128, KU705186, KU705189, KU705131, KU705134, KU705138 |
| V | 2 | 1 | 1 | 0 | 0 | KU705198, KU705153 |
| W | 13 | 2 | 1 | 4 | 6 | KU705176, KU705177, KU705179, KU705125, KU705127, KU705129, KU705145, KU705197, KU705185, KU705132, KU705135, KU705137, KU705156 |
| X | 8 | 6 | 1 | 1 | 0 | KU705142, KU705146, KU705188, KU705149, KU705150, KU705155, KU705159 |
| Y | 5 | 5 | 0 | 0 | 0 | KU705143, KU705144, KU705148, KU705151, KU705157 |
| 16S-24 | 1 | 1 | 0 | 0 | 0 | KU705147 |
| 16S-25 | 1 | 0 | 0 | 0 | 1 | KU705130 |
| 16S-26 | 1 | 0 | 0 | 0 | 1 | KU705139 |
| 16S-30 | 1 | 0 | 0 | 0 | 1 | KU705126 |
| Total | 54 | 21 | 3 | 14 | 16 | --- |
| Cervid Species and Origin | Number of Deer Sampled, Total (Male, Female) | Age Group, Total (Male, Female) | ||
|---|---|---|---|---|
| <1 Year | 1–2 Years | >2 Years | ||
| Roe deer, wild | 30 (9, 21) | 2 (1, 1) | 22 (6, 16) | 6 (2, 4) |
| Fallow deer, wild | 7 (7, 0) | 0 | 3 (3, 0) | 4 (4, 0) |
| Sika deer, wild | 22 (17, 5) | 0 | 13 (11, 2) | 9 (6, 3) |
| Red deer, wild | 15 (10, 5) | 6 (5, 1) | 2 (1, 1) | 7 (4, 3) |
| Combined wild deer | 74 (43, 31) | 8 (6, 2) | 40 (21, 19) | 26 (16, 10) |
| Red deer, farmed | 27 (17, 10) | 25 (17, 8) | 0 | 2 (0, 2) |
| Total deer | 101 (60, 41) | 33 (23, 10) | 40 (21, 19) | 28 (16, 12) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silaghi, C.; Fröhlich, J.; Reindl, H.; Hamel, D.; Rehbein, S. Anaplasma phagocytophilum and Babesia Species of Sympatric Roe Deer (Capreolus capreolus), Fallow Deer (Dama dama), Sika Deer (Cervus nippon) and Red Deer (Cervus elaphus) in Germany. Pathogens 2020, 9, 968. https://doi.org/10.3390/pathogens9110968
Silaghi C, Fröhlich J, Reindl H, Hamel D, Rehbein S. Anaplasma phagocytophilum and Babesia Species of Sympatric Roe Deer (Capreolus capreolus), Fallow Deer (Dama dama), Sika Deer (Cervus nippon) and Red Deer (Cervus elaphus) in Germany. Pathogens. 2020; 9(11):968. https://doi.org/10.3390/pathogens9110968
Chicago/Turabian StyleSilaghi, Cornelia, Julia Fröhlich, Hubert Reindl, Dietmar Hamel, and Steffen Rehbein. 2020. "Anaplasma phagocytophilum and Babesia Species of Sympatric Roe Deer (Capreolus capreolus), Fallow Deer (Dama dama), Sika Deer (Cervus nippon) and Red Deer (Cervus elaphus) in Germany" Pathogens 9, no. 11: 968. https://doi.org/10.3390/pathogens9110968