Critical Review: Propensity of Premise Plumbing Pipe Materials to Enhance or Diminish Growth of Legionella and Other Opportunistic Pathogens

 , and
, and
Abstract
:1. Introduction
- Growing direct or indirect potable water reuse, which can sometimes alter levels of nutrients and Cu+2 in the source water [67].
- Emphasis on and investment in green building design for water and energy efficiency and associated unintended consequences for in-building hydraulics (e.g., more stagnation, higher surface area to volume ratios of water to plumbing surfaces, required hot water recirculation systems) that alter water chemistry and delivery of nutrients or disinfectants [39,54,71,72].
- Greater use of plastic pipes (e.g., PEX, PVC, polyethylene), which vary in leaching potential by type of plastic and due to the presence of proprietary stabilizers and processes [73].
- Heightened concern about an array of bacterial OPs besides Legionella, including Pseudomonas aeruginosa, Acinetobacter baumannii, and NTM, as well as amoebae (e.g. Acanthamoeba, Vermamoeba), which can themselves be pathogenic or can serve as host organisms for bacterial OP proliferation [8].
2. Direct Effects of Plumbing Material on Pathogen Growth
2.1. Copper Has Both Antimicrobial and Micronutrient Properties
2.2. Copper Pipe as an Antimicrobial Material in Premise Plumbing
2.2.1. Noteworthy Limitations to Copper’s Antimicrobial Efficacy
2.2.2. Water Chemistry Effects on Copper Bioavailability
2.2.3. Copper as a Nutrient in Premise Plumbing
2.3. Direct Release of Organic Carbon by Plastics
2.4. Iron Release from Pipes
2.5. Zinc, Aluminum, Magnesium Plumbing Materials
3. Indirect Effects of Pipe Material on Pathogen Growth
3.1. Pipe Material Effect on Disinfectant Availability
3.2. Effect of Metallic Plumbing Materials on Nutrient Availability via Autotrophic Carbon Fixation
3.2.1. Hydrogen Oxidizing Bacteria
3.2.2. Autotrophic Ammonia and Iron Oxidizing Bacteria
3.2.3. Copper Deposition Corrosion Accelerating H2 Evolution
3.3. Pipe Scaling Effects
4. Influence of Plumbing System Design, Configuration and Operation
4.1. Water Stagnation
4.2. Hot Water Recirculation Lines
4.3. Pipe Aging
4.4. Possible Mixed Material Interactions
5. Mediating Role of Microbiome and other Microbiological Considerations
5.1. The Role of Pipe Material in Shaping the Premise Plumbing Microbiome and Resident Amoeba Host Organisms
5.2. Variation in Copper Tolerance Among Species and Strains
5.3. Confounding Effects of VBNC Bacteria
5.4. Virulence
- Elevated copper levels, selecting for resistance to copper overload within macrophage phagosomes, a component of the innate immune response [309].
- Elevated iron levels, influencing interactions between iron homeostasis and virulence.
- Exposure to lipids, which are generally not well removed by drinking water treatment, priming OPs for lipid-rich environments within hosts. Accumulation of phospholipid fatty acids has been shown to be greater in the biofilms of polyethylene pipes than copper pipes, though these lipids were putatively associated with bacteria [310].
- Low DO levels, selecting for OPs capable of survival in low DO regions of the biofilm in infected host tissue.
- Exposure to eukaryotic predation, selecting for resistance to the host’s immune response (e.g., lung macrophages) or enhanced virulence.
5.5. Antibiotic Resistance and Tolerance
6. Conclusions
- Both the intra-species and inter-species variation of copper resistance amongst OPs, as well as environmental drivers of this variation.
- Effects of copper pipes on OPs in a more holistic sense, with identification of real-world conditions that are drivers for discrepancies in copper’s antimicrobial capacity.
- Copper’s possible micronutrient activity in OPs within premise plumbing contexts, including threshold concentrations required for various physiological functions, as well as physicochemical and ecological factors that influence those thresholds.
- The disease risk that VBNC OPs pose and conditions under which copper and other antimicrobials induce VBNC status in premise plumbing OPs
- The inhibitory action of trace metals on OP growth in premise plumbing, as well as growth requirements for other trace elements exhibited by OPs in premise plumbing.
- Potential mediating effects of the wider microbial community composition resulting from pipe material on OPs.
- Effect of mixed pipe materials on physicochemical parameters of bulk water and OP growth.
- The effects of plumbing materials on OP antibiotic resistance and virulence.
- The impact of stagnation, velocity, sediments, corrosion control, and consumer water use patterns on all of the above.
Author Contributions
Funding
Conflicts of Interest
References
- National Academies of Sciences, Engineering, and Medicine. Management of Legionella in Water Systems; The National Academies Press: Washington, DC, USA, 2019; ISBN 978-0-309-49947-7. [Google Scholar]
- Ashbolt, N.J. Microbial Contamination of Drinking Water and Human Health from Community Water Systems. Curr. Environ. Health Rep. 2015, 2, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United States Occupational Safety & Health Administration Legionnaires’ Disease: Facts and Frequently Asked Questions. Available online: https://www.osha.gov/dts/osta/otm/legionnaires/faq.html (accessed on 1 July 2020).
- United States Occupational Safety & Health Administration Legionnaires’ Disease. Available online: https://www.osha.gov/dts/osta/otm/otm_iii/otm_iii_7.html (accessed on 1 July 2020).
- CDC Legionella (Legionnaires’ Disease and Pontiac Fever) Surveillance and Reporting. Available online: http://www.cdc.gov/legionella/surv-reporting.html (accessed on 1 July 2020).
- Singh, R.; Hamilton, K.A.; Rasheduzzaman, M.; Yang, Z.; Kar, S.; Fasnacht, A.; Masters, S.V.; Gurian, P.L. Managing Water Quality in Premise Plumbing: Subject Matter Experts’ Perspectives and a Systematic Review of Guidance Documents. Water 2020, 12, 347. [Google Scholar] [CrossRef] [Green Version]
- McCoy, W.F. Preventing Legionellosis; IWA Publishing: London, UK, 2005; ISBN 1843390949. [Google Scholar]
- Falkinham, J.; Pruden, A.; Edwards, M. Opportunistic Premise Plumbing Pathogens: Increasingly Important Pathogens in Drinking Water. Pathogens 2015, 4, 373. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control Legionella (Legionnaires’ Disease and Pontiac Fever). Available online: https://www.cdc.gov/legionella/index.html (accessed on 1 July 2020).
- Gomez-Valero, L.; Rusniok, C.; Rolando, M.; Neou, M.; Dervins-Ravault, D.; Demirtas, J.; Rouy, Z.; Moore, R.J.; Chen, H.; Petty, N.K.; et al. Comparative analyses of Legionella species identifies genetic features of strains causing Legionnaires’ disease. Genome Biol. 2014, 15, 505. [Google Scholar] [CrossRef]
- Association of Water Technologies Legionella 2019: A Position Statement and Guidance Document. Available online: https://www.awt.org/pub/035C2942-03BE-3BFF-08C3-4C686FB7395C (accessed on 15 November 2020).
- Allen, M.J.; Edberg, S.C.; Clancy, J.L.; Hrudey, S.E. Drinking water microbial myths. Crit. Rev. Microbiol. 2015, 41, 366–373. [Google Scholar] [CrossRef]
- Barrabeig, I.; Rovira, A.; Garcia, M.; Oliva, J.M.; Vilamala, A.; Ferrer, M.D.; SabriÀ, M.; DomÍNguez, A. Outbreak of Legionnaires’ disease associated with a supermarket mist machine. Epidemiol. Infect. 2010, 138, 1823–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahoney, F.J.; Hoge, C.W.; Farley, T.A.; Barbaree, J.M.; Breiman, R.F.; Benson, R.F.; McFarland, L.M. Communitywide Outbreak of Legionnaires’ Disease Associated with a Grocery Store Mist Machine. J. Infect. Dis. 1992, 165, 736–739. [Google Scholar] [CrossRef] [PubMed]
- Stout, J.E.; Yu, V.L.; Yee, Y.C.; Vaccarello, S.; Diven, W.; Lee, T.C. Legionella pneumophila in residential water supplies: Environmental surveillance with clinical assessment for Legionnaires’ disease. Epidemiol. Infect. 1992, 109, 49–57. [Google Scholar]
- Stout, J.E.; Victor, L.Y.; Muraca, P. Isolation of Legionella pneumophila from the cold water of hospital ice machines: Implications for origin and transmission of the organism. Infect. Control Hosp. Epidemiol. 1985, 6, 141–146. [Google Scholar] [CrossRef]
- Rhoads, W.J.; Bradley, T.N.; Mantha, A.; Buttling, L.; Keane, T.; Pruden, A.; Edwards, M.A. Residential water heater cleaning and occurrence of Legionella in Flint, MI. Water Res. 2020, 115439. [Google Scholar] [CrossRef]
- Gobin, I.; Newton, P.R.; Hartland, E.L.; Newton, H.J. Infections caused by nonpneumophila species of Legionella. Rev. Med. Microbiol. 2009, 20, 1–11. [Google Scholar] [CrossRef]
- Joseph, C.A. Legionnaires’ disease in Europe 2000–2002. Epidemiol. Infect. 2004, 132, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stout, J.E.; Yu, V.L.; Muraca, P.; Joly, J.; Troup, N.; Tompkins, L.S. Potable water as a cause of sporadic cases of community-acquired legionnaires’ disease. N. Engl. J. Med. 1992, 326, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Fliermans, C.B.; Cherry, W.B.; Orrison, L.H.; Smith, S.J.; Tison, D.L.; Pope, D.H. Ecological distribution of Legionella pneumophila. Appl. Environ. Microbiol. 1981, 41, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.F.; Jones-Lee, A. Public Health Significance of Waterborne Pathogens in Domestic Water Supplies and Reclaimed Water. In Report to State of California Environmental Protection Agency Comparative Risk Project; California Environmental Protection Agency: Berkeley, CA, USA, 1993. [Google Scholar]
- Borella, P.; Montagna, M.T.; Romano-Spica, V.; Stampi, S.; Stancanelli, G.; Triassi, M.; Neglia, R.; Marchesi, I.; Fantuzzi, G.; Tatò, D.; et al. Legionella Infection Risk from Domestic Hot Water. Emerg. Infect. Dis. 2004, 10, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Falkinham III, J.O. Common Features of Opportunistic Premise Plumbing Pathogens. Int. J. Environ. Res. Public Health 2015, 12, 4533–4545. [Google Scholar] [CrossRef] [PubMed]
- Parr, A.; Whitney, E.A.; Berkelman, R.L. Legionellosis on the Rise: A Review of Guidelines for Prevention in the United States. J. public Health Manag. Pract. 2015, 21, E17–E26. [Google Scholar] [CrossRef] [Green Version]
- Rhoads, W.J.; Garner, E.; Ji, P.; Zhu, N.; Parks, J.; Schwake, D.O.; Pruden, A.; Edwards, M.A. Distribution System Operational Deficiencies Coincide with Reported Legionnaires’ Disease Clusters in Flint, Michigan. Environ. Sci. Technol. 2017, 51, 11986–11995. [Google Scholar] [CrossRef]
- Mercante, J.W.; Winchell, J.M. Current and Emerging Legionella Diagnostics for Laboratory and Outbreak Investigations. Clin. Microbiol. Rev. 2015, 28, 95–133. [Google Scholar] [CrossRef] [Green Version]
- Garrison, L.E. Vital Signs: Deficiencies in Environmental Control Identified in Outbreaks of Legionnaires’ Disease—North America, 2000–2014. MMWR. Morb. Mortal. Wkly. Rep. 2016, 65, 576–584. [Google Scholar] [CrossRef] [Green Version]
- Pierre, D.; Baron, J.L.; Ma, X.; Sidari, F.P.; Wagener, M.M.; Stout, J.E. Water Quality as a Predictor of Legionella Positivity of Building Water Systems. Pathogens 2019, 8, 295. [Google Scholar] [CrossRef] [Green Version]
- Perrin, Y.; Bouchon, D.; Delafont, V.; Moulin, L.; Héchard, Y. Microbiome of drinking water: A full-scale spatio-temporal study to monitor water quality in the Paris distribution system. Water Res. 2019, 149, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhang, Y.; Knibbe, W.-J.; Feng, C.; Liu, W.; Medema, G.; van der Meer, W. Potential impacts of changing supply-water quality on drinking water distribution: A review. Water Res. 2017, 116, 135–148. [Google Scholar] [CrossRef]
- Pinto, A.J.; Xi, C.; Raskin, L. Bacterial community structure in the drinking water microbiome is governed by filtration processes. Environ. Sci. Technol. 2012, 46, 8851–8859. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Masters, S.; Falkinham, J.O.; Edwards, M.A.; Pruden, A. Distribution System Water Quality Affects Responses of Opportunistic Pathogen Gene Markers in Household Water Heaters. Environ. Sci. Technol. 2015, 49, 8416–8424. [Google Scholar] [CrossRef] [PubMed]
- Baron, J.L.; Vikram, A.; Duda, S.; Stout, J.E.; Bibby, K. Shift in the Microbial Ecology of a Hospital Hot Water System following the Introduction of an On-Site Monochloramine Disinfection System. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwering, M.; Song, J.; Louie, M.; Turner, R.J.; Ceri, H. Multi-species biofilms defined from drinking water microorganisms provide increased protection against chlorine disinfection. Biofouling 2013, 29, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Greub, G.; Raoult, D. Microorganisms Resistant to Free-Living Amoebae. Clin. Microbiol. Rev. 2004, 17, 413. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Edwards, M.A.; Falkinham, J.O.; Pruden, A. Probiotic Approach to Pathogen Control in Premise Plumbing Systems? A Review. Environ. Sci. Technol. 2013, 47, 10117–10128. [Google Scholar] [CrossRef]
- Wargo, M.J. Is the Potable Water System an Advantageous Preinfection Niche for Bacteria Colonizing the Cystic Fibrosis Lung? MBio 2019, 10, e00883-19. [Google Scholar] [CrossRef] [Green Version]
- Rhoads, J.W.; Pearce, A.; Pruden, A.; Edwards, M.A. Anticipating the Effects of Green Buildings on Water Quality and Infrastructure. J. Am. Water Works Assoc. 2015, 107, 50. [Google Scholar] [CrossRef]
- Liu, G.; Tao, Y.; Zhang, Y.; Lut, M.; Knibbe, W.-J.; van der Wielen, P.; Liu, W.; Medema, G.; van der Meer, W. Hotspots for selected metal elements and microbes accumulation and the corresponding water quality deterioration potential in an unchlorinated drinking water distribution system. Water Res. 2017, 124, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Proctor, C.R.; Hammes, F. Drinking water microbiology - from measurement to management. Curr. Opin. Biotechnol. 2015, 33, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Neu, L.; Hammes, F. Feeding the Building Plumbing Microbiome: The Importance of Synthetic Polymeric Materials for Biofilm Formation and Management. Water 2020, 12, 1774. [Google Scholar] [CrossRef]
- Zhang, Y.; Edwards, M. Accelerated chloramine decay and microbial growth by nitrification in premise plumbing. J. Am. Water Works Assoc. 2009, 101, 51–62. [Google Scholar] [CrossRef]
- Al-Jasser, A.O. Chlorine decay in drinking-water transmission and distribution systems: Pipe service age effect. Water Res. 2007, 41, 387–396. [Google Scholar] [CrossRef]
- Durand, M.L. Disinfectants and Plumbing Materials: Effects on the Sensory and Chemical Characteristics of Drinking Water; Virginia Tech: Blacksburg, VA, USA, 2005. [Google Scholar]
- Haas, C.N.; Gupta, M.; Chitluru, R.; Burlingame, G. Chlorine Demand in disinfecting Water Mains. J. Am. Water Works Assoc. 2002, 94, 97–102. [Google Scholar] [CrossRef]
- Clark, R.; Yang, Y.J.; Impellitteri, C.; Haught, R.; Schupp, D.; Panguluri, S.; Krishnan, R. Chlorine Fate and Transport in Distribution Systems: Experimental and modeling studies. J. Am. Water Works Assoc. 2010, 102, 144–156. [Google Scholar] [CrossRef]
- Zhang, C.; Li, C.; Zheng, X.; Zhao, J.; He, G.; Zhang, T. Effect of pipe materials on chlorine decay, trihalomethanes formation, and bacterial communities in pilot-scale water distribution systems. Int. J. Environ. Sci. Technol. 2017, 14, 85–94. [Google Scholar] [CrossRef]
- Lehtola, M.J.; Miettinen, I.T.; Lampola, T.; Hirvonen, A.; Vartiainen, T.; Martikainen, P.J. Pipeline materials modify the effectiveness of disinfectants in drinking water distribution systems. Water Res. 2005, 39, 1962–1971. [Google Scholar] [CrossRef]
- Heim, T.H.; Dietrich, A.M. Sensory aspects and water quality impacts of chlorinated and chloraminated drinking water in contact with HDPE and cPVC pipe. Water Res. 2007, 41, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Wang, Y.; Hammes, F. Short-term organic carbon migration from polymeric materials in contact with chlorinated drinking water. Sci. Total Environ. 2018, 613, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.L.; Dietrich, A.M. Contributions of silane cross-linked PEX pipe to chemical/solvent odours in drinking water. Water Sci. Technol. 2007, 55, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Koo, J.; Kim, S. A general framework of chlorine decay modeling at a pilot-scale water distribution system. J. Water Supply Res. Technol. AQUA 2015, 64, 543–557. [Google Scholar] [CrossRef]
- Nguyen, C.; Elfland, C.; Edwards, M. Impact of advanced water conservation features and new copper pipe on rapid chloramine decay and microbial regrowth. Water Res. 2012, 46, 611–621. [Google Scholar] [CrossRef]
- Westbrook, A.; Digiano, F.A. Rate of chloramine decay at pipe surfaces. J. Am. Water Works Assoc. 2009, 101, 59–70. [Google Scholar] [CrossRef]
- Bucheli-Witschel, M.; Kötzsch, S.; Darr, S.; Widler, R.; Egli, T. A new method to assess the influence of migration from polymeric materials on the biostability of drinking water. Water Res. 2012, 46, 4246–4260. [Google Scholar] [CrossRef]
- Skjevrak, I.; Due, A.; Gjerstad, K.O.; Herikstad, H. Volatile organic components migrating from plastic pipes (HDPE, PEX and PVC) into drinking water. Water Res. 2003, 37, 1912–1920. [Google Scholar] [CrossRef]
- Williams, C.L.; Neu, H.M.; Gilbreath, J.J.; Michel, S.L.J.; Zurawski, D.V.; Merrell, D.S. Copper Resistance of the Emerging Pathogen Acinetobacter baumannii. Appl. Environ. Microbiol. 2016, 82, 6174–6188. [Google Scholar] [CrossRef] [Green Version]
- Proctor, C.R.; Dai, D.; Edwards, M.A.; Pruden, A. Interactive effects of temperature, organic carbon, and pipe material on microbiota composition and Legionella pneumophila in hot water plumbing systems. Microbiome 2017, 5, 130. [Google Scholar] [CrossRef]
- Rushing, J.C.; McNeill, L.S.; Edwards, M. Some effects of aqueous silica on the corrosion of iron. Water Res. 2003, 37, 1080–1090. [Google Scholar] [CrossRef]
- Niu, L.; Cheng, Y.F. Corrosion behavior of X-70 pipe steel in near-neutral pH solution. Appl. Surf. Sci. 2007, 253, 8626–8631. [Google Scholar] [CrossRef]
- Lin, Y.-S.E.; Vidic, R.D.; Stout, J.E.; Yu, V.L. Individual and combined effects of copper and silver ions on inactivation of Legionella pneumophila. Water Res. 1996, 30, 1905–1913. [Google Scholar] [CrossRef]
- Mathys, W.; Stanke, J.; Harmuth, M.; Junge-Mathys, E. Occurrence of Legionella in hot water systems of single-family residences in suburbs of two German cities with special reference to solar and district heating. Int. J. Hyg. Environ. Health 2008, 211, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Buse, H.Y.; Gomez-Alvarez, V.; Struewing, I.; Santo Domingo, J.; Ashbolt, N.J. Impact of drinking water conditions and copper materials on downstream biofilm microbial communities and Legionella pneumophila colonization. J. Appl. Microbiol. 2014, 117, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Merritt, K.; Brown, S.A. Release of hexavalent chromium from corrosion of stainless steel and cobalt—Chromium alloys. J. Biomed. Mater. Res. 1995, 29, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Morton, S.C.; Zhang, Y.; Edwards, M.A. Implications of nutrient release from iron metal for microbial regrowth in water distribution systems. Water Res. 2005, 39, 2883–2892. [Google Scholar] [CrossRef]
- Garner, E.; Zhu, N.; Strom, L.; Edwards, M.; Pruden, A. A human exposome framework for guiding risk management and holistic assessment of recycled water quality. Environ. Sci. Water Res. Technol. 2016, 2, 580–598. [Google Scholar] [CrossRef] [Green Version]
- Anderson, L.E.; Krkošek, W.H.; Stoddart, A.K.; Trueman, B.F.; Gagnon, G.A. Lake Recovery Through Reduced Sulfate Deposition: A New Paradigm for Drinking Water Treatment. Environ. Sci. Technol. 2017, 51, 1414–1422. [Google Scholar] [CrossRef] [Green Version]
- Monteith, D.T.; Stoddard, J.L.; Evans, C.D.; De Wit, H.A.; Forsius, M.; Høgåsen, T.; Wilander, A.; Skjelkvåle, B.L.; Jeffries, D.S.; Vuorenmaa, J.; et al. Dissolved organic carbon trends resulting from changes in atmospheric deposition chemistry. Nature 2007, 450, 537–540. [Google Scholar] [CrossRef]
- European Environment Agency. Air Quality Standards. Available online: https://www.eea.europa.eu/themes/air/air-quality-concentrations/air-quality-standards (accessed on 28 October 2020).
- Rhoads, W.J.; Pruden, A.; Edwards, M.A. Survey of green building water systems reveals elevated water age and water quality concerns. Environ. Sci. Water Res. Technol. 2015, 2, 164–173. [Google Scholar] [CrossRef] [Green Version]
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development. Available online: https://sustainabledevelopment.un.org/post2015/transformingourworld/publication (accessed on 15 November 2020).
- Lee, J.; Kleczyk, E.; Bosch, D.J.; Dietrich, A.M.; Lohani, V.K.; Loganathan, G.V. Homeowners’ decision-making in a premise plumbing failure-prone area. J. Am. Water Works Assoc. 2013, 105, E236–E241. [Google Scholar] [CrossRef]
- Ramamurthy, T.; Ghosh, A.; Pazhani, G.P.; Shinoda, S. Current Perspectives on Viable but Non-Culturable (VBNC) Pathogenic Bacteria. Front. Public Health 2014, 2, 103. [Google Scholar] [CrossRef] [PubMed]
- Alleron, L.; Khemiri, A.; Koubar, M.; Lacombe, C.; Coquet, L.; Cosette, P.; Jouenne, T.; Frere, J. VBNC Legionella pneumophila cells are still able to produce virulence proteins. Water Res. 2013, 47, 6606–6617. [Google Scholar] [CrossRef] [PubMed]
- Dietersdorfer, E.; Kirschner, A.; Schrammel, B.; Ohradanova-Repic, A.; Stockinger, H.; Sommer, R.; Walochnik, J.; Cervero-Aragó, S. Starved viable but non-culturable (VBNC) Legionella strains can infect and replicate in amoebae and human macrophages. Water Res. 2018, 141, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Copper Development Association Inc. Copper Facts. Available online: https://www.copper.org/education/c-facts/facts-print.html (accessed on 18 August 2020).
- Copper Development Association Inc. Antimicrobial Copper Surfaces. Available online: https://www.copper.org/applications/antimicrobial/ (accessed on 18 August 2020).
- European Chemicals Agency. Information on Biocides. Available online: https://www.echa.europa.eu/web/guest/information-on-chemicals/biocidal-active-substances?p_p_id=dissactivesubstances_WAR_dissactivesubstancesportlet&p_p_lifecycle=0&p_p_state=normal&p_p_mode=view&p_p_col_id=column-1&p_p_col_pos=2&p_p_col_count=3&_dissacti (accessed on 10 November 2020).
- Samanovic, M.I.; Ding, C.; Thiele, D.J.; Darwin, K.H. Copper in microbial pathogenesis: Meddling with the metal. Cell Host Microbe 2012, 11, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulay, N.; Edwards, M. Copper in the Urban Water Cycle. Crit. Rev. Environ. Sci. Technol. 2000, 30, 297–326. [Google Scholar] [CrossRef]
- World Health Organization. Copper in Drinking-Water; World Health Organization: Geneva, Switzerland, 2016.
- Lin, Y.E.; Vidic, R.D.; Stout, J.E.; Victor, L.Y. Negative effect of high pH on biocidal efficacy of copper and silver ions in controlling Legionella pneumophila. Appl. Environ. Microbiol. 2002, 68, 2711–2715. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.S.; Stout, J.E.; Yu, V.L.; Vidic, R.D. Disinfection of water distribution systems for Legionella. Semin. Respir. Infect. 1998, 13, 147–159. [Google Scholar]
- Hajialilo, E.; Niyyati, M.; Solaymani, M.; Rezaeian, M. Pathogenic Free-Living Amoebae Isolated From Contact Lenses of Keratitis Patients. Iran. J. Parasitol. 2015, 10, 541–546. [Google Scholar]
- Centers for Disease Control and Prevention, National Center for Emerging and Zoonotic Infectious Diseases (NCEZID), Division of Foodborne, Waterborne, and E.D. (DFWED) Parasites—Naegleria fowleri—Primary Amebic Meningoencephalitis (PAM)—Amebic Encephalitis. Available online: https://www.cdc.gov/parasites/naegleria/general.html (accessed on 21 July 2020).
- Centers for Disease Control and Prevention, National Center for Emerging and Zoonotic Infectious Diseases (NCEZID), Division of Foodborne, Waterborne, and E.D. (DFWED) Parasites—Acanthamoeba—Granulomatous Amebic Encephalitis (GAE). Keratitis. Available online: https://www.cdc.gov/parasites/acanthamoeba/gen_info/acanthamoeba.html (accessed on 10 August 2020).
- Thomas, V.; Bouchez, T.; Nicolas, V.; Robert, S.; Loret, J.F.; Lévi, Y. Amoebae in domestic water systems: Resistance to disinfection treatments and implication in Legionella persistence. J. Appl. Microbiol. 2004, 97, 950–963. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervantes, H.I.; Álvarez, J.A.; Muñoz, J.M.; Arreguín, V.; Mosqueda, J.L.; Macías, A.E. Antimicrobial activity of copper against organisms in aqueous solution: A case for copper-based water pipelines in hospitals? Am. J. Infect. Control 2013, 41, e115–e118. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-I.; Shih, H.-Y.; Lee, C.-M.; Yang, T.C.; Lay, J.-J.; Lin, Y.E. In vitro efficacy of copper and silver ions in eradicating Pseudomonas aeruginosa, Stenotrophomonas maltophilia and Acinetobacter baumannii: Implications for on-site disinfection for hospital infection control. Water Res. 2008, 42, 73–80. [Google Scholar] [CrossRef]
- Shih, H.-Y.Y.; Lin, Y.E. Efficacy of Copper-Silver Ionization in Controlling Biofilm- and Plankton-Associated Waterborne Pathogens. Appl. Environ. Microbiol. 2010, 76, 2032–2035. [Google Scholar] [CrossRef] [Green Version]
- Cateau, E.; Delafont, V.; Hechard, Y.; Rodier, M.H. Free-living amoebae: What part do they play in healthcare-associated infections? J. Hosp. Infect. 2014, 87, 131–140. [Google Scholar] [CrossRef]
- Cateau, E.; Verdon, J.; Fernandez, B.; Hechard, Y.; Rodier, M.-H. Acanthamoeba sp. promotes the survival and growth of Acinetobacter baumanii. FEMS Microbiol. Lett. 2011, 319, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention, National Center for Emerging and Zoonotic Infectious Diseases (NCEZID), D. of H.Q.P. (DHQP) Staphylococcus aureus in Healthcare Settings|HAI|CDC. Available online: https://www.cdc.gov/hai/organisms/staph.html (accessed on 10 August 2020).
- Kozajda, A.; Jeżak, K.; Kapsa, A. Airborne Staphylococcus aureus in different environments-a review. Environ. Sci. Pollut. Res. Int. 2019, 26, 34741–34753. [Google Scholar] [CrossRef] [Green Version]
- Plipat, N.; Spicknall, I.H.; Koopman, J.S.; Eisenberg, J.N. The dynamics of methicillin-resistant Staphylococcus aureus exposure in a hospital model and the potential for environmental intervention. BMC Infect. Dis. 2013, 13, 595. [Google Scholar] [CrossRef] [Green Version]
- Landeen, L.K.; Yahya, M.T.; Kutz, S.M.; Gerba, C.P. Microbiological Evaluation of Copper: Silver Disinfection Units for Use in Swimming Pools. Water Sci. Technol. 1989, 21, 267–270. [Google Scholar] [CrossRef]
- Yahya, M.T.; Landeen, L.K.; Messina, M.C.; Kutz, S.M.; Schulze, R.; Gerba, C.P. Disinfection of bacteria in water systems by using electrolytically generated copper:silver and reduced levels of free chlorine. Can. J. Microbiol. 1990, 36, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Hopkin, M. MRSA “hiding in hospital sinks and vases”. Nature 2006. [Google Scholar] [CrossRef]
- Huws, S.A.; Smith, A.W.; Enright, M.C.; Wood, P.J.; Brown, M.R.W. Amoebae promote persistence of epidemic strains of MRSA. Environ. Microbiol. 2006, 8, 1130–1133. [Google Scholar] [CrossRef] [PubMed]
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denton, M.; Kerr, K.G. Microbiological and clinical aspects of infection associated with Stenotrophomonas maltophilia. Clin. Microbiol. Rev. 1998, 11, 57–80. [Google Scholar] [CrossRef] [Green Version]
- Gomes, I.B.; Simões, L.C.; Simões, M. Influence of surface copper content on Stenotrophomonas maltophilia biofilm control using chlorine and mechanical stress. Biofouling 2020, 36, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.M.; Ashbolt, N.J. Do free-living amoebae in treated drinking water systems present an emerging health risk? Environ. Sci. Technol. 2011, 45, 860–869. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention, National Center for Emerging and Zoonotic Infectious Diseases (NCEZID), D. of H.Q.P. (DHQP) Nontuberculous Mycobacteria (NTM) Infections|HAI|CDC. Available online: https://www.cdc.gov/hai/organisms/nontuberculous-mycobacteria.html (accessed on 10 August 2020).
- Johnson, M.M.; Odell, J.A. Nontuberculous mycobacterial pulmonary infections. J. Thorac. Dis. 2014, 6, 210–220. [Google Scholar] [CrossRef]
- Kusnetsov, J.; Iivanainen, E.; Elomaa, N.; Zacheus, O.; Martikainen, P.J. Copper and silver ions more effective against Legionellae than against mycobacteria in a hospital warm water system. Water Res. 2001, 35, 4217–4225. [Google Scholar] [CrossRef]
- Lin, Y.E.; Vidic, R.D.; Stout, J.E.; McCartney, C.A.; Yu, V.L. Inactivation of Mycobacterium avium by copper and silver ions. Water Res. 1998, 32, 1997–2000. [Google Scholar] [CrossRef]
- Rhoads, W.J.; Pruden, A.; Edwards, M.A. Interactive Effects of Corrosion, Copper, and Chloramines on Legionella and Mycobacteria in Hot Water Plumbing. Environ. Sci. Technol. 2017, 51, 7065–7075. [Google Scholar] [CrossRef] [PubMed]
- Mullis, S.N.; Falkinham, J.O. Adherence and biofilm formation of Mycobacterium avium, Mycobacterium intracellulare and Mycobacterium abscessus to household plumbing materials. J. Appl. Microbiol. 2013, 115, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, J.D.; Falkow, S.; Tompkins, L.S.; Bermudez, L.E. Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infect. Immun. 1997, 65, 3759–3767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igbinosa, I.H.; Igumbor, E.U.; Aghdasi, F.; Tom, M.; Okoh, A.I. Emerging Aeromonas species infections and their significance in public health. ScientificWorldJournal. 2012, 2012, 625023. [Google Scholar] [CrossRef] [Green Version]
- Assant, M.A.; Roy, D.; Montpetit, D. Adhesion of Aeromonas hydrophila to Water Distribution System Pipes after Different Contact Times. J. Food Prot. 1998, 61, 1321–1329. [Google Scholar] [CrossRef]
- Rahman, M.; Abd, H.; Romling, U.; Sandstrom, G.; Möllby, R. Aeromonas-Acanthamoeba interaction and early shift to a viable but nonculturable state of Aeromonas by Acanthamoeba. J. Appl. Microbiol. 2008, 104, 1449–1457. [Google Scholar] [CrossRef]
- Delafont, V.; Perraud, E.; Brunet, K.; Maisonneuve, E.; Kaaki, S.; Rodier, M.H. Vermamoeba vermiformis in hospital network: A benefit for Aeromonas hydrophila. Parasitol. Res. 2019, 118, 3191–3194. [Google Scholar] [CrossRef]
- National Center for Immunization and Respiratory Diseases, D. of B.D. Legionnaires Disease Signs and Symptoms|Legionella|CDC. Available online: https://www.cdc.gov/legionella/about/signs-symptoms.html (accessed on 10 August 2020).
- National Center for Immunization and Respiratory Diseases, D. of B.D. Legionnaires Disease Cause and Spread|Legionella|CDC. Available online: https://www.cdc.gov/legionella/about/causes-transmission.html (accessed on 10 August 2020).
- Stout, J.E.; Lin, Y.-S.E.; Goetz, A.M.; Muder, R.R. Controlling Legionella in hospital water systems: Experience with the superheat-and-flush method and copper-silver ionization. Infect. Control Hosp. Epidemiol. 1998, 19, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Miuetzner, S.; Schwille, R.C.; Farley, A.; Wald, E.R.; Ge, J.H.; States, S.J.; Libert, T.; Wadowsky, R.M. Efficacy of thermal treatment and copper-silver ionization for controlling Legionella pneumophila in high-volume hot water plumbing systems in hospitals. Am. J. Infect. Control 1997, 25, 452–457. [Google Scholar] [CrossRef]
- Gião, M.S.; Wilks, S.A.; Keevil, C.W. Influence of copper surfaces on biofilm formation by Legionella pneumophila in potable water. Biometals 2015, 28, 329. [Google Scholar] [CrossRef] [PubMed]
- Buse, H.Y.; Lu, J.; Struewing, I.T.; Ashbolt, N.J. Preferential colonization and release of Legionella pneumophila from mature drinking water biofilms grown on copper versus unplasticized polyvinylchloride coupons. Int. J. Hyg. Environ. Health 2014, 217, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Rowbotham, T.J. Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae. J. Clin. Pathol. 1980, 33, 1179–1183. [Google Scholar] [CrossRef] [Green Version]
- Grossi, M.; Dey, R.; Ashbolt, N. Searching for Activity Markers that Approximate (VBNC) Legionella pneumophila Infectivity in Amoeba after Ultraviolet (UV) Irradiation. Water 2018, 10, 1219. [Google Scholar] [CrossRef] [Green Version]
- Bodey, G.P.; Bolivar, R.; Fainstein, V.; Jadeja, L. Infections Caused by Pseudomonas aeruginosa. Rev. Infect. Dis. 1983, 5, 279–313. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.J.; Dehnbostel, J.; Blackwell, T.S. Pseudomonas aeruginosa: Host defence in lung diseases. Respirology 2010, 15, 1037–1056. [Google Scholar] [CrossRef] [PubMed]
- Bédard, E.; Charron, D.; Lalancette, C.; Déziel, E.; Prévost, M. Recovery of Pseudomonas aeruginosa culturability following copper- and chlorine-induced stress. FEMS Microbiol. Lett. 2014, 356, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Moritz, M.M.; Flemming, H.C.; Wingender, J. Integration of Pseudomonas aeruginosa and Legionella pneumophila in drinking water biofilms grown on domestic plumbing materials. Int. J. Hyg. Environ. Health 2010, 213, 190–197. [Google Scholar] [CrossRef]
- Jeanvoine, A.; Meunier, A.; Puja, H.; Bertrand, X.; Valot, B.; Hocquet, D. Contamination of a hospital plumbing system by persister cells of a copper-tolerant high-risk clone of Pseudomonas aeruginosa. Water Res. 2019, 157, 579–586. [Google Scholar] [CrossRef]
- Petignat, C.; Francioli, P.; Nahimana, I.; Wenger, A.; Bille, J.; Schaller, M.-D.; Revelly, J.-P.; Zanetti, G.; Blanc, D.S. Exogenous Sources of Pseudomonas aeruginosa in Intensive Care Unit Patients Implementation of Infection Control Measures and Follow-Up With Molecular Typing. Infect. Control Hosp. Epidemiol. 2006, 27, 953–957. [Google Scholar] [CrossRef]
- Teitzel, G.M.; Parsek, M.R. Heavy Metal Resistance of Biofilm and Planktonic Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2003, 69, 2313. [Google Scholar] [CrossRef] [Green Version]
- Dwidjosiswojo, Z.; Richard, J.; Moritz, M.M.; Dopp, E.; Flemming, H.-C.; Wingender, J. Influence of copper ions on the viability and cytotoxicity of Pseudomonas aeruginosa under conditions relevant to drinking water environments. Int. J. Hyg. Environ. Health 2011, 214, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Dopp, E.; Richard, J.; Dwidjosiswojo, Z.; Simon, A.; Wingender, J. Influence of the copper-induced viable but non-culturable state on the toxicity of Pseudomonas aeruginosa towards human bronchial epithelial cells in vitro. Int. J. Hyg. Environ. Health 2017, 220, 1363–1369. [Google Scholar] [CrossRef] [PubMed]
- Pukatzki, S.; Kessin, R.H.; Mekalanos, J.J. The human pathogen Pseudomonas aeruginosa utilizes conserved virulence pathways to infect the social amoeba Dictyostelium discoideum. Proc. Natl. Acad. Sci. USA 2002, 99, 3159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- US EPA Office of Water. Technologies for Legionella Control in Premise Plumbing Systems; US Environmental Protection Agency: Washington, DC, USA, 2016.
- Hans, M.; Erbe, A.; Mathews, S.; Chen, Y.; Solioz, M.; Mücklich, F. Role of Copper Oxides in Contact Killing of Bacteria. Langmuir 2013, 29, 16160–16166. [Google Scholar] [CrossRef] [PubMed]
- Cachafeiro, S.P.; Naveira, I.M.; García, I.G. Is copper-silver ionisation safe and effective in controlling legionella? J. Hosp. Infect. 2007, 67, 209–216. [Google Scholar] [CrossRef]
- Kim, B.R.; Anderson, J.E.; Mueller, S.A.; Gaines, W.A.; Kendall, A.M. Literature review—Efficacy of various disinfectants against Legionella in water systems. Water Res. 2002, 36, 4433–4444. [Google Scholar] [CrossRef]
- Rakshit, A.; Khatua, K.; Shanbhag, V.; Comba, P.; Datta, A. Cu2+ selective chelators relieve copper-induced oxidative stress: In vivo. Chem. Sci. 2018, 9, 7916–7930. [Google Scholar] [CrossRef] [Green Version]
- Van der Kooij, D.; Veenendaal, H.R.; Scheffer, W.J.H. Biofilm formation and multiplication of Legionella in a model warm water system with pipes of copper, stainless steel and cross-linked polyethylene. Water Res. 2005, 39, 2789–2798. [Google Scholar] [CrossRef]
- Dodrill, D.M.; Edwards, M. Corrosion control on the basis of utility experience. J. Am. Water Works Assoc. 1995, 87, 74–85. [Google Scholar] [CrossRef]
- Kimbrough, D.E. Brass corrosion as a source of lead and copper in traditional and all-plastic distribution systems. J. Am. Water Works Assoc. 2007, 99, 70–76. [Google Scholar] [CrossRef]
- June, S.G.; Dziewulski, D.M. Copper and Silver Biocidal Mechanisms, Resistance Strategies, and Efficacy for Legionella Control. Journal-American Water Work. Assoc. 2018, 110, E13–E35. [Google Scholar] [CrossRef]
- Assaidi, A.; Ellouali, M.; Latrache, H.; Mabrouki, M.; Hamadi, F.; Timinouni, M.; Zahir, H.; El Mdaghri, N.; Barguigua, A.; Mliji, E.M. Effect of temperature and plumbing materials on biofilm formation by Legionella pneumophila serogroup 1 and 2-15. J. Adhes. Sci. Technol. 2018, 32, 1471–1484. [Google Scholar] [CrossRef]
- Ashbolt, N.J. Environmental (Saprozoic) Pathogens of Engineered Water Systems: Understanding Their Ecology for Risk Assessment and Management. Pathog. 2015, 4, 390–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morvay, A.A.; Decun, M.; Scurtu, M.; Sala, C.; Morar, A.; Sarandan, M. Biofilm formation on materials commonly used in household drinking water systems. Water Sci. Technol. Water Supply 2011, 11, 252–257. [Google Scholar] [CrossRef]
- Dai, D.; Proctor, C.R.; Williams, K.; Edwards, M.A.; Pruden, A. Mediation of effects of biofiltration on bacterial regrowth, Legionella pneumophila, and the microbial community structure under hot water plumbing conditions. Environ. Sci. Water Res. Technol. 2018, 4, 183–194. [Google Scholar] [CrossRef]
- Soothill, J.S. Carbapenemase-bearing Klebsiella spp. in sink drains: Investigation into the potential advantage of copper pipes. J. Hosp. Infect. 2016, 93, 152–154. [Google Scholar] [CrossRef]
- Norton, C.D.; LeChevallier, M.W.; Falkinham, J.O. Survival of Mycobacterium avium in a model distribution system. Water Res. 2004, 38, 1457–1466. [Google Scholar] [CrossRef]
- Leoni, E.; De Luca, G.; Legnani, P.P.; Sacchetti, R.; Stampi, S.; Zanetti, F. Legionella waterline colonization: Detection of Legionella species in domestic, hotel and hospital hot water systems. J. Appl. Microbiol. 2005, 98, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Marrie, T.; Green, P.; Burbridge, S.; Bezanson, G.; Neale, S.; Hoffman, P.S.; Haldane, D. Legionellaceae in the potable water of Nova Scotia hospitals and Halifax residences. Epidemiol. Infect. 1994, 112, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Borella, P.; Montagna, M.T.; Stampi, S.; Stancanelli, G.; Romano-Spica, V.; Triassi, M.; Marchesi, I.; Bargellini, A.; Tatò, D.; Napoli, C. Legionella contamination in hot water of Italian hotels. Appl. Environ. Microbiol. 2005, 71, 5805–5813. [Google Scholar] [CrossRef] [Green Version]
- Mathys, W.; Hohmann, C.P.; Junge-Mathys, E. Efficacy of Copper-Silver Ionization in Controlling Legionella in a Hospital Hot Water Distribution System: A German Experience. In Legionella; Marre, R., Kwaik, Y.A., Bartlett, C., Cianciotto, N.P., Fields, B.S., Frosch, M., Hacker, J., Luck, P.C., Eds.; ASM Press: Washington, DC, USA, 2002; pp. 419–424. ISBN 1-55581-230-9. [Google Scholar]
- Bédard, E.; Paranjape, K.; Lalancette, C.; Villion, M.; Quach, C.; Laferrière, C.; Faucher, S.P.; Prévost, M. Legionella pneumophila levels and sequence-type distribution in hospital hot water samples from faucets to connecting pipes. Water Res. 2019, 156, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Bédard, E.; Trigui, H.; Liang, J.; Doberva, M.; Paranjape, K.; Lalancette, C.; Faucher, S.P.; Prévost, M. Local adaptation of Legionella pneumophila within a hospital hot water system increases tolerance to copper. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Lytle, D.A.; Liggett, J. Impact of water quality on chlorine demand of corroding copper. Water Res. 2016, 92, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.; Powers, K.; Hidmi, L.; Schock, M.R. The role of Pipe Ageing in Copper Corrosion by-Product Release; IWA Publishing: London, UK, 2001; Volume 1, pp. 25–32. [Google Scholar]
- Zevenhuizen, L.P.T.M.; Dolfing, J.; Eshuis, E.J.; Scholten-Koerselman, I.J. Inhibitory effects of copper on bacteria related to the free ion concentration. Microb. Ecol. 1979, 5, 139–146. [Google Scholar] [CrossRef]
- Garvey, J.E.; Owen, H.A.; Winner, R.W. Toxicity of copper to the green alga, Chlamydomonas reinhardtii (Chlorophyceae), as affected by humic substances of terrestrial and freshwater origin. Aquat. Toxicol. 1991, 19, 89–96. [Google Scholar] [CrossRef]
- Meador, J.P. The interaction of pH, dissolved organic carbon, and total copper in the determination of ionic copper and toxicity. Aquat. Toxicol. 1991, 19, 13–31. [Google Scholar] [CrossRef]
- Ebrahimpour, M.; Alipour, H.; Rakhshah, S. Influence of water hardness on acute toxicity of copper and zinc on fish. Toxicol. Ind. Health 2010, 26, 361–365. [Google Scholar] [CrossRef]
- Pourkhabbaz, A.; Kasmani, M.E.; Kiyani, V.; Hosynzadeh, M.H. Effects of water hardness and Cu and Zn on Lc50 in Gambusia holbrooki. Chem. Speciat. Bioavailab. 2011, 23, 224–228. [Google Scholar] [CrossRef]
- Riethmuller, N.; Markich, S.; Parry, D.; van Dam, R. The Effect of True Water Hardness and Alkalinity on the Toxicity of Cu and U to Two Tropical Australian Freshwater Organisms; Supervising Scientist, Environment Australia: Darwin NT, Australia, 2000.
- Yang, S.; Amy, P.; Edwards, M.; Rhoads, W. Natural Organic Matter, Orthophosphate, pH, and Growth Phase Can Limit Copper Antimicrobial Efficacy for Legionella in Drinking Water. Environ. Sci. Technol. 2020. Submitted. [Google Scholar]
- Stauber, J.L.; Florence, T.M. Mechanism of toxicity of ionic copper and copper complexes to algae. Mar. Biol. 1987, 94, 511–519. [Google Scholar] [CrossRef]
- Ives, D.J.G.; Rawson, A.E. Copper Corrosion. J. Electrochem. Soc. 1962, 109, 447. [Google Scholar] [CrossRef]
- Galai, S.; Touhami, Y.; Marzouki, M.N. Response surface methodology applied to laccases activities exhibited by Stenotrophomonas maltophilia AAP56 in different growth conditions. BioResources 2012, 7, 706–726. [Google Scholar]
- Wingender, J.; Flemming, H.C. Biofilms in drinking water and their role as reservoir for pathogens. Int. J. Hyg. Environ. Health 2011, 214, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Van der Kooij, D.; van der Wielen, P.W.J.J.; Rosso, D.; Shaw, A.; Borchardt, D.; Ibisch, R.; Apgar, D.; Witherspoon, J.; Di Toro, D.M.; Paquin, P.R. Microbial Growth in Drinking Water Supplies; IWA Publishing: London, UK, 2013; ISBN 1780400403. [Google Scholar]
- Connell, M.; Stenson, A.; Weinrich, L.; LeChevallier, M.; Boyd, S.L.; Ghosal, R.R.; Dey, R.; Whelton, A.J. PEX and PP Water Pipes: Assimilable Carbon, Chemicals, and Odors. J. Am. Water Work Assoc. 2016, 108, E192–E204. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, D.; Kajihara, Y.; Shimidzu, N.; Hamamura, K.; Nagase, M. Hydrogen sulfide production by sulfate-reducing bacteria utilizing additives eluted from plastic resins. Waste Manag. Res. 2011, 29, 594–601. [Google Scholar] [CrossRef]
- Kelley, K.M. The Impact of Cross-linked Polyethylene (PEX) Pipe on Drinking Water Chemical and Odor Quality; University of South Alabama: Mobile, AL, USA, 2014. [Google Scholar]
- Corfitzen, C.B. Afgivelse af Organisk Stof Fra Polymere Materialer—Mikrobiel Vækst [Release of Organic Matter from Polymeric Materials—Microbial Growth]; The Danish Environmental Protection Agency: Odense, Denmark, 2002. [Google Scholar]
- Kowalska, B.; Kowalski, D.; Rozej, A. Organic compounds migrating from plastic pipes into water. J. Water Supply Res. Technol. AQUA 2011, 60, 137–146. [Google Scholar] [CrossRef]
- Pedersen, K. Biofilm development on stainless steel and pvc surfaces in drinking water. Water Res. 1990, 24, 239–243. [Google Scholar] [CrossRef]
- Hallam, N.B.; West, J.R.; Forster, C.F.; Simms, J. The potential for biofilm growth in water distribution systems. Water Res. 2001, 35, 4063–4071. [Google Scholar] [CrossRef]
- Thomson, R.M.; Carter, R.; Tolson, C.; Coulter, C.; Huygens, F.; Hargreaves, M. Factors associated with the isolation of Nontuberculous mycobacteria (NTM) from a large municipal water system in Brisbane, Australia. BMC Microbiol. 2013, 13, 89. [Google Scholar] [CrossRef] [Green Version]
- Learbuch, K.L.G.; Lut, M.C.; Liu, G.; Smidt, H.; van der Wielen, P.W.J.J. Legionella growth potential of drinking water produced by a reverse osmosis pilot plant. Water Res. 2019, 157, 55–63. [Google Scholar] [CrossRef]
- Buse, H.Y.; Morris, B.; Struewing, I.T.; Szabo, J.G. Chlorine and monochloramine disinfection of Legionella pneumophila colonizing copper and PVC drinking water biofilms. Appl. Environ. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Griffin, A.; Edwards, M. Effect of nitrification on corrosion of galvanized iron, copper, and concrete. J. Am. Water Works Assoc. 2010, 102, 83–93. [Google Scholar] [CrossRef]
- Sarin, P.; Snoeyink, V.L.; Bebee, J.; Jim, K.K.; Beckett, M.A.; Kriven, W.M.; Clement, J.A. Iron release from corroded iron pipes in drinking water distribution systems: Effect of dissolved oxygen. Water Res. 2004, 38, 1259–1269. [Google Scholar] [CrossRef]
- WHO. Legionella and the Prevention of Legionellosis; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Faraldo-Gómez, J.D.; Sansom, M.S.P. Acquisition of siderophores in gram-negative bacteria. Nat. Rev. Mol. Cell Biol. 2003, 4, 105–116. [Google Scholar] [CrossRef]
- States, S.J.; Conley, L.F.; Ceraso, M.; Stephenson, T.E.; Wolford, R.S.; Wadowsky, R.M.; McNamara, A.M.; Yee, R.B. Effects of metals on Legionella pneumophila growth in drinking water plumbing systems. Appl. Environ. Microbiol. 1985, 50, 1149–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakić, A.; Perić, J.; Foglar, L. Influence of temperature, chlorine residual and heavy metals on the presence of Legionella pneumophila in hot water distribution systems. Ann. Agric. Environ. Med. 2012, 19, 431–436. [Google Scholar] [PubMed]
- van der Lugt, W.; Euser, S.M.; Bruin, J.P.; Den Boer, J.W.; Walker, J.T.; Crespi, S. Growth of Legionella anisa in a model drinking water system to evaluate different shower outlets and the impact of cast iron rust. Int. J. Hyg. Environ. Health 2017, 220, 1295–1308. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Strom, O.; Pruden, A.; Edwards, M. Copper Pipe, Stagnant Conditions, Corrosion Control, and Trace Disinfectant Residual Enhanced Reduction of Legionella pneumophila during Simulations of the Flint Water Crisis. Pathogens 2020, in press. [Google Scholar] [CrossRef]
- Wang, H.; Masters, S.; Hong, Y.; Stallings, J.; Falkinham, J.O.; Edwards, M.A.; Pruden, A. Effect of Disinfectant, Water Age, and Pipe Material on Occurrence and Persistence of Legionella, mycobacteria, Pseudomonas aeruginosa, and Two Amoebas. Environ. Sci. Technol. 2012, 46, 11566–11574. [Google Scholar] [CrossRef]
- Oyem, H.H.; Oyem, I.M.; Usese, A.I. Iron, manganese, cadmium, chromium, zinc and arsenic groundwater contents of Agbor and Owa communities of Nigeria. Springerplus 2015, 4, 104. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Zinc in Drinking Water: Background Document for Development of Guidelines for Drinking-Water Quality. Health Criteria and Other Supporting Information; World Health Organization: Geneva, Switzerland, 2003; Volume 1. [Google Scholar]
- World Health Organization Water Sanitation. Guidelines for Drinking-Water Quality: Incorporating First Addendum. Vol. 1, Recommendations; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- Salehi, M.; Odimayomi, T.; Ra, K.; Ley, C.; Julien, R.; Nejadhashemi, A.P.; Hernandez-Suarez, J.S.; Mitchell, J.; Shah, A.D.; Whelton, A. An investigation of spatial and temporal drinking water quality variation in green residential plumbing. Build. Environ. 2020, 169, 106566. [Google Scholar] [CrossRef]
- Sharrett, A.R.; Carter, A.P.; Orheimt, R.M.; Feinleib, M. Daily intake of lead, cadmium, copper, and zinc from drinking water: The seattle study of trace metal exposure. Environ. Res. 1982, 28, 456–475. [Google Scholar] [CrossRef]
- Howard, C.D. Zinc Contamination in Drinking Water. J. Am. Water Works Assoc. 1923, 10, 411–414. [Google Scholar] [CrossRef]
- Silver, S.; Lusk, J.E. Bacterial Magnesium, Manganese, and Zinc Transport. In Ion Transport in Prokaryotes; Rosen, B.P., Silver, S., Eds.; Academic Press: Cambridge, MA, USA, 1987; pp. 165–180. ISBN 978-0-12-596935-2. [Google Scholar]
- Choudhury, R.; Srivastava, S. Zinc resistance mechanisms in bacteria. Curr. Sci. 2001, 81, 768–775. [Google Scholar]
- Ma, L.; Terwilliger, A.; Maresso, A.W. Iron and zinc exploitation during bacterial pathogenesis. Metallomics 2015, 7, 1541–1554. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sharma, M.R.; Koripella, R.K.; Yang, Y.; Kaushal, P.S.; Lin, Q.; Wade, J.T.; Gray, T.A.; Derbyshire, K.M.; Agrawal, R.K.; et al. Zinc depletion induces ribosome hibernation in mycobacteria. Proc. Natl. Acad. Sci. USA 2018, 115, 8191. [Google Scholar] [CrossRef] [Green Version]
- Suryawati, B. Zinc homeostasis mechanism and its role in bacterial virulence capacity. AIP Conf. Proc. 2018, 2021, 070021. [Google Scholar] [CrossRef]
- Nies, D.H. Resistance to cadmium, cobalt, zinc, and nickel in microbes. Plasmid 1992, 27, 17–28. [Google Scholar] [CrossRef]
- Reeves, M.W.; Pine, L.; Hutner, S.H.; George, J.R.; Harrell, W.K. Metal requirements of Legionella pneumophila. J. Clin. Microbiol. 1981, 13, 688–695. [Google Scholar] [CrossRef] [Green Version]
- Kirschner, R.A.; Parker, B.C.; Falkinham, J.O. Epidemiology of Infection by Nontuberculous Mycobacteria: Mycobacterium avium, Mycobacterium intracellulare, and Mycobacterium scrofulaceum in Acid, Brown-Water Swamps of the Southeastern United States and Their Association with Environmental Variables. Am. Rev. Respir. Dis. 1992, 145, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Norberg, A.B.; Molin, N. Toxicity of cadmium, cobalt, uranium and zinc to Zoogloea ramigera. Water Res. 1983, 17, 1333–1336. [Google Scholar] [CrossRef]
- Nweke, C.; Chibuogwu, O.; Alisi, C. Response of planktonic bacteria of New Calabar River to zinc stress. Afr. J. Biotechnol. 2006, 5, 653–658. [Google Scholar]
- Nweke, C.; Alisi, C.; Chibuogwu, O.; Nwanyanwu, C. Toxicity of Zinc to heterotrophic bacteria from a tropical river sediment. Appl. Ecol. Environ. Res. 2007, 5, 123–132. [Google Scholar] [CrossRef]
- Babich, H.; Stotzky, G. Toxicity of zinc to fungi, bacteria, and coliphages: Influence of chloride ions. Appl. Environ. Microbiol. 1978, 36, 906. [Google Scholar] [CrossRef] [Green Version]
- United States Environmental Protection Agency. Secondary Drinking Water Standards: Guidance for Nuisance Chemicals; United States Environmental Protection Agency: Washington, DC, USA, 2015.
- Standardization Administration of China Guobiao 5749-2006. Available online: https://www.chinesestandard.net/PDF.aspx/GB5749-2006 (accessed on 10 November 2020).
- Hwang, M.G.; Katayama, H.; Ohgaki, S. Effect of Intracellular Resuscitation of Legionella pneumophila in Acanthamoeba polyphage Cells on the Antimicrobial Properties of Silver and Copper. Environ. Sci. Technol. 2006, 40, 7434–7439. [Google Scholar] [CrossRef]
- Barker, J.; Brown, M.R.; Collier, P.J.; Farrell, I.; Gilbert, P. Relationship between Legionella pneumophila and Acanthamoeba polyphaga: Physiological status and susceptibility to chemical inactivation. Appl. Environ. Microbiol. 1992, 58, 2420–2425. [Google Scholar] [CrossRef] [Green Version]
- García, M.T.; Jones, S.; Pelaz, C.; Millar, R.D.; Abu Kwaik, Y. Acanthamoeba polyphaga resuscitates viable non-culturable Legionella pneumophila after disinfection. Environ. Microbiol. 2007, 9, 1267–1277. [Google Scholar] [CrossRef]
- Adékambi, T.; Ben Salah, S.; Khlif, M.; Raoult, D.; Drancourt, M. Survival of Environmental Mycobacteria in Acanthamoeba polyphaga. Appl. Environ. Microbiol. 2006, 72, 5974. [Google Scholar] [CrossRef] [Green Version]
- Cervero-Aragó, S.; Rodríguez-Martínez, S.; Puertas-Bennasar, A.; Araujo, R.M. Effect of Common Drinking Water Disinfectants, Chlorine and Heat, on Free Legionella and Amoebae-Associated Legionella. PLoS ONE 2015, 10, e0134726. [Google Scholar] [CrossRef] [Green Version]
- Marciano-Cabral, F.; Jamerson, M.; Kaneshiro, E.S. Free-living amoebae, Legionella and Mycobacterium in tap water supplied by a municipal drinking water utility in the USA. J. Water Health 2009, 8, 71–82. [Google Scholar] [CrossRef]
- Inkinen, J.; Jayaprakash, B.; Ahonen, M.; Pitkänen, T.; Mäkinen, R.; Pursiainen, A.; Santo Domingo, J.W.; Salonen, H.; Elk, M.; Keinänen-Toivola, M.M. Bacterial community changes in copper and PEX drinking water pipeline biofilms under extra disinfection and magnetic water treatment. J. Appl. Microbiol. 2018, 124, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Vikesland, P.J.; Valentine, R.L. Reaction Pathways Involved in the Reduction of Monochloramine by Ferrous Iron. Environ. Sci. Technol. 2000, 34, 83–90. [Google Scholar] [CrossRef]
- Zhang, Z.; Stout, J.E.; Yu, V.L.; Vidic, R. Effect of pipe corrosion scales on chlorine dioxide consumption in drinking water distribution systems. Water Res. 2008, 42, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.K.; Powers, K.A.; Raetz, M.A.; Parks, J.L.; Edwards, M.A. Rapid free chlorine decay in the presence of Cu(OH)2: Chemistry and practical implications. Water Res. 2011, 45, 5302–5312. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Qu, J.; Liu, R.; Zhao, X.; Qiang, Z. The influence of Cu(II) on the decay of monochloramine. Chemosphere 2009, 74, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.A.; Parks, J.; Griffin, A.; Raetz, M.A.; Martin, A.K.; Scardina, P.; Elfland, C. Lead and Copper Corrosion Control in New Construction; The Water Research Foundation: Denver, CO, USA, 2011; ISBN 978-1-60573-137-7. [Google Scholar]
- Brazeau, R.H.; Edwards, M.A. Role of Hot Water System Design on Factors Influential to Pathogen Regrowth: Temperature, Chlorine Residual, Hydrogen Evolution, and Sediment. Environ. Eng. Sci. 2013, 30, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Ishizaki, A.; Tanaka, K.; Taga, N. Microbial production of poly-D-3-hydroxybutyrate from CO2x. Appl. Microbiol. Biotechnol. 2001, 57, 6–12. [Google Scholar] [CrossRef]
- Edwards, M.; Pruden, A.; Falkinham III, J.O.; Brazeau, R.; Williams, K.; Wang, H.; Martin, A.; Rhoads, W. Relationship between Biodegradable Organic Matter and Pathogen Concentrations in Premise Plumbing. Available online: https://www.researchgate.net/publication/282650492_Relationship_Between_Biodegradable_Organic_Matter_and_Pathogen_Concentrations_in_Premise_Plumbing (accessed on 7 October 2020).
- Dai, D.; Rhoads, W.J.; Edwards, M.A.; Pruden, A. Shotgun Metagenomics Reveals Taxonomic and Functional Shifts in Hot Water Microbiome Due to Temperature Setting and Stagnation. Front. Microbiol. 2018, 9, 2695. [Google Scholar] [CrossRef]
- González-Cabaleiro, R.; Curtis, T.P.; Ofiţeru, I.D. Bioenergetics analysis of ammonia-oxidizing bacteria and the estimation of their maximum growth yield. Water Res. 2019, 154, 238–245. [Google Scholar] [CrossRef]
- Neubauer, S.C.; Emerson, D.; Megonigal, J.P. Life at the energetic edge: Kinetics of circumneutral iron oxidation by lithotrophic iron-oxidizing bacteria isolated from the wetland-plant rhizosphere. Appl. Environ. Microbiol. 2002, 68, 3988–3995. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.K.; Edwards, M.A.; Pruden, A.J.; Falkinham, J.O. Organic Carbon Generation Mechanisms in Main and Premise Distribution Systems; Virginia Tech: Blacksburg, VA, USA, 2012. [Google Scholar]
- Clark, B.; Clair, J.S.; Edwards, M. Copper Deposition Corrosion Elevates Lead Release to Potable Water. J. Am. Water Works Assoc. 2015, 107, E627–E637. [Google Scholar] [CrossRef]
- Brazeau, R.H.; Edwards, M.A.; Falkinham, J.O.; Pearce, A.R.; Pruden, A.J. Sustainability of Residential Hot Water Infrastructure: Public Health, Environmental Impacts, and Consumer Drivers; Virginia Tech: Blacksburg, VA, USA, 2012. [Google Scholar]
- Fox, P.; Abbaszadegan, M. Impact of Scale Formation on Biofilm Growth in Premise Plumbing. Available online: https://www.semanticscholar.org/paper/Impact-of-Scale-Formation-on-Biofilm-Growth-in-Fox-Abbaszadegan/b615ff0df2516a1bac2ac11854019ab0554e6fc2 (accessed on 31 August 2020).
- Sarin, P.; Snoeyink, V.L.; Lytle, D.A.; Kriven, W.M. Iron Corrosion Scales: Model for Scale Growth, Iron Release, and Colored Water Formation. J. Environ. Eng. 2004, 130, 364–373. [Google Scholar] [CrossRef]
- Yang, F.; Shi, B.; Gu, J.; Wang, D.; Yang, M. Morphological and physicochemical characteristics of iron corrosion scales formed under different water source histories in a drinking water distribution system. Water Res. 2012, 46, 5423–5433. [Google Scholar] [CrossRef] [PubMed]
- Tuovinen, O.H.; Button, K.S.; Vuorinen, A.; Carlson, L.; Mair, D.M.; Yut, L.A. Bacterial, chemical, and mineralogical characteristics of tubercles in distribution pipelines. J. Am. Water Works Assoc. 1980, 72, 626–635. [Google Scholar] [CrossRef]
- Lee, W.; Lewandowski, Z.; Nielsen, P.H.; Allan Hamilton, W. Role of sulfate-reducing bacteria in corrosion of mild steel: A review. Biofouling 1995, 8, 165–194. [Google Scholar] [CrossRef]
- Nguyen, C.K.; Stone, K.R.; Dudi, A.; Edwards, M.A. Corrosive Microenvironments at Lead Solder Surfaces Arising from Galvanic Corrosion with Copper Pipe. Environ. Sci. Technol. 2010, 44, 7076–7081. [Google Scholar] [CrossRef]
- Hasit, Y.J.; Anderson, J.L.; Anthony, J.; Parolari, T.; Rockaway, T.D.; Frenc, M.L. Distribution Water Quality Issues Related to New Development or Low Usage; Awwa Research Foudation: Denver, CO, USA, 2006. [Google Scholar]
- McNeill, L.S.; Edwards, M. Iron pipe corrosion in distribution systems. J. Am. Water Works Assoc. 2001, 93, 88–100. [Google Scholar] [CrossRef]
- Nawrocki, J.; Raczyk-Stanisławiak, U.; Świetlik, J.; Olejnik, A.; Sroka, M.J. Corrosion in a distribution system: Steady water and its composition. Water Res. 2010, 44, 1863–1872. [Google Scholar] [CrossRef]
- Merkel, T.H.; Groß, H.J.; Werner, W.; Dahlke, T.; Reicherter, S.; Beuchle, G.; Eberle, S.H. Copper corrosion by-product release in long-term stagnation experiments. Water Res. 2002, 36, 1547–1555. [Google Scholar] [CrossRef]
- Lytle, D.A.; Schock, M.R. Impact of stagnation time on metal dissolution from plumbing materials in drinking water. J. Water Supply Res. Technol. AQUA 2000, 49, 243–257. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, S. Investigation of organic compounds migration from polymeric pipes into drinking water under long retention times. Procedia Eng. 2014, 70, 1753–1761. [Google Scholar] [CrossRef] [Green Version]
- Richard, R.; Hamilton, K.A.; Westerhoff, P.; Boyer, T.H. Tracking copper, chlorine, and occupancy in a new, multi-story, institutional green building. Environ. Sci. Water Res. Technol. 2020, 6, 1672–1680. [Google Scholar] [CrossRef]
- Haig, S.J.; Kotlarz, N.; Lipuma, J.J.; Raskin, L. A high-throughput approach for identification of nontuberculous mycobacteria in drinking water reveals relationship between water age and Mycobacterium avium. MBio 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, C.; Proctor, C.; Singh, G.; Ra, K.; Noh, Y.; Odimayomi, T.; Salehi, M.; Julien, R.; Mitchell, J.; Nejadhashemi, A.P.; et al. Drinking water microbiology in a water-efficient building: Stagnation, seasonality, and physiochemical effects on opportunistic pathogen and total bacteria proliferation. Environ. Sci. Water Res. Technol. 2020. [Google Scholar] [CrossRef]
- Hozalski, R.M.; Lapara, T.M.; Zhao, X.; Kim, T.; Waak, M.B.; Burch, T.; Mccarty, M. Flushing of stagnant premise water systems after the COVID-19 shutdown can reduce infection risk by Legionella and Mycobacterium spp. medRxiv 2020. [Google Scholar] [CrossRef]
- Sánchez-Busó, L.; Olmos, M.P.; Camaró, M.L.; Adrián, F.; Calafat, J.M.; González-Candelas, F. Phylogenetic analysis of environmental Legionella pneumophila isolates from an endemic area (Alcoy, Spain). Infect. Genet. Evol. 2015, 30, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Pryor, M.; Springthorpe, S.; Riffard, S.; Brooks, T.; Huo, Y.; Davis, G.; Sattar, S.A. Investigation of opportunistic pathogens in municipal drinking water under different supply and treatment regimes. Water Sci. Technol. 2004, 50, 83–90. [Google Scholar] [CrossRef]
- Guidance for Reopening Buildings After Prolonged Shutdown or Reduced Operation|CDC. Available online: https://www.cdc.gov/coronavirus/2019-ncov/php/building-water-system.html (accessed on 26 October 2020).
- Proctor, C.; Rhoads, W.; Keane, T.; Salehi, M.; Hamilton, K.; Pieper, K.J.; Cwiertny, D.M.; Prévost, M.; Whelton, A. Considerations for Large Building Water Quality after Extended Stagnation. AWWA Water Sci. 2020, 2, e1186. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. Information on Maintaining or Restoring Water Quality in Buildings with Low or No Use. Available online: https://www.epa.gov/coronavirus/information-maintaining-or-restoring-water-quality-buildings-low-or-no-use (accessed on 26 October 2020).
- Rhoads, W.J.; Prévost, M.; Pieper, K.J.; Keane, T.; Whelton, A.J.; Rölli, F.; Proctor, C.R.; Grimard-Conea, M. Responding to Water Stagnation in Buildings with Reduced or No Water Use; American Water Works Association: Denver, CO, USA, 2020. [Google Scholar]
- 2021 Uniform Plumbing Code; Twenty-Nin.; International Association of Plumbing and Mechanical Officials: Ontario, CA, USA, 2020.
- 2018 International Plumbing Code; International Code Council, Inc.: Country Club Hills, IL, USA, 2017; ISBN 978-1-60983-745-7.
- 2015 International Energy Conservation Code; International Code Council, Inc: Country Club Hills, IL, USA, 2014; ISBN 978-1-60983-486-9.
- Ally, M.R. Water and Energy Savings using Demand Hot Water Recirculating Systems in Residential Homes: A Case Study of Five Homes in Palo Alto, California. Available online: http://www.osti.gov/servlets/purl/885864-jNK2Il/ (accessed on 26 August 2020).
- Rhoads, W.J.; Ji, P.; Pruden, A.; Edwards, M.A. Water heater temperature set point and water use patterns influence Legionella pneumophila and associated microorganisms at the tap. Microbiome 2015, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Flannery, B.; Gelling, L.B.; Vugia, D.J.; Weintraub, J.M.; Salerno, J.J.; Conroy, M.J.; Stevens, V.A.; Rose, C.E.; Moore, M.R.; Fields, B.S.; et al. Reducing Legionella colonization of water systems with monochloramine. Emerg. Infect. Dis. 2006, 12, 588. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.J.; Cook, D.C.; Townsend, H.E. Characterization of Iron Oxides Commonly Formed as Corrosion Products on Steel. Hyperfine Interact. 1998, 112, 59–66. [Google Scholar] [CrossRef]
- Stout, J.E.; Yu, V.L.; Best, M.G. Ecology of Legionella pneumophila within water distribution systems. Appl. Environ. Microbiol. 1985, 49, 221–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Wu, C.; Zhong, D.; Yuan, Y.; Shan, L.; Zhang, J. Effects of pipe materials on chlorine-resistant biofilm formation under long-term high chlorine level. Appl. Biochem. Biotechnol. 2014, 173, 1564–1578. [Google Scholar] [CrossRef] [PubMed]
- Sancy, M.; Gourbeyre, Y.; Sutter, E.M.M.; Tribollet, B. Mechanism of corrosion of cast iron covered by aged corrosion products: Application of electrochemical impedance spectrometry. Corros. Sci. 2010, 52, 1222–1227. [Google Scholar] [CrossRef]
- Salehi, M.; Abouali, M.; Wang, M.; Zhou, Z.; Nejadhashemi, A.P.; Mitchell, J.; Caskey, S.; Whelton, A.J. Case study: Fixture water use and drinking water quality in a new residential green building. Chemosphere 2018. [Google Scholar] [CrossRef]
- Lund, V.; Anderson-Glenna, M.; Skjevrak, I.; Steffensen, I.-L. Long-term study of migration of volatile organic compounds from cross-linked polyethylene (PEX) pipes and effects on drinking water quality. J. Water Health 2011, 9, 483–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanemura, D.; Yamada, K.; Nishimura, H.; Igawa, K.; Higuchi, Y. Investigation of degradation mechanism by copper catalytic activity and mechanical property of polyethylene pipes for hot water supply. Annu. Tech. Conf. ANTEC Conf. Proc. 2014, 3, 2022–2026. [Google Scholar]
- Wright, D. Failure of Plastics and Rubber Products: Causes, Effects and Case Studies Involving Degradation; Rapra Technology Limited: Shawbury, UK, 2001; ISBN 185957517X. [Google Scholar]
- Plastic Pipe Institute. Proper Integration of Copper Tubing and Components with PP-R Piping Materials for Plumbing Applications; The Plastic Pipe Institute: Irving, TX, USA, 2018. [Google Scholar]
- Bulletin, A.T. Intermixing Copper Tube and Aquatherm; Aquatherm GmbH: Attendom, Germany, 2012. [Google Scholar]
- Huang, X.; Pieper, K.J.; Cooper, H.K.; Diaz-Amaya, S.; Zemlyanov, D.Y.; Whelton, A.J. Corrosion of upstream metal plumbing components impact downstream PEX pipe surface deposits and degradation. Chemosphere 2019, 236, 124329. [Google Scholar] [CrossRef]
- Lewis, J.R. The Catalytic Decomposition of Sodium Hypochlorite Solutions.II. Iron Oxide as Promoter in the Copper Oxide Catalysis of Sodium Hypochlorite. J. Phys. Chem. 1928, 32, 1808–1819. [Google Scholar] [CrossRef]
- Cruse, H. Dissolved-Copper Effect on Iron Pipe. J. Am. Water Works Assoc. 1971, 63, 79–81. [Google Scholar] [CrossRef]
- Cartier, C.; Nour, S.; Richer, B.; Deshommes, E.; Prévost, M. Impact of water treatment on the contribution of faucets to dissolved and particulate lead release at the tap. Water Res. 2012, 46, 5205–5216. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, M.K.; Triantafyllidou, S.; Schock, M.R.; Lytle, D.A. Mineralogical Evidence of Galvanic Corrosion in Drinking Water Lead Pipe Joints. Environ. Sci. Technol. 2018, 52, 3365–3374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.G. Chapter 10: Galvanic Corrosion. In Uhlig’s Corrosion Handbook: Third Edition; Revie, R.W., Ed.; The Electrochemical Society: Pennington, NJ, USA, 2011; pp. 123–143. ISBN 978-0-8031-0981-0. [Google Scholar]
- Hack, H.P. Galvanic Corrosion—Google Books; American Society for Testing and Materials: Philadelphia, PA, USA, 1988. [Google Scholar]
- Souto, R.M.; González-García, Y.; Bastos, A.C.; Simões, A.M. Investigating corrosion processes in the micrometric range: A SVET study of the galvanic corrosion of zinc coupled with iron. Corros. Sci. 2007, 49, 4568–4580. [Google Scholar] [CrossRef]
- Marques, A.G.; Taryba, M.G.; Panão, A.S.; Lamaka, S.V.; Simões, A.M. Application of scanning electrode techniques for the evaluation of iron-zinc corrosion in nearly neutral chloride solutions. Corros. Sci. 2016, 104, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Kuntyi, O.I.; Zozulya, H.I.; Dobrovets’ka, O.Y.; Kornii, S.A.; Reshetnyak, O.V. Deposition of Copper, Silver, and Nickel on Aluminum by Galvanic Replacement. Mater. Sci. 2018, 53, 488–494. [Google Scholar] [CrossRef]
- Jorcin, J.-B.; Blanc, C.; Pébère, N.; Tribollet, B.; Vivier, V. Galvanic Coupling Between Pure Copper and Pure Aluminum. J. Electrochem. Soc. 2008, 155, C46. [Google Scholar] [CrossRef]
- Clark, B.N.; Masters, S.V.; Edwards, M.A. Lead Release to Drinking Water from Galvanized Steel Pipe Coatings. Environ. Eng. Sci. 2015, 32, 713–721. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, C.; Xu, S.; Gao, Y.; Jiang, S. Accelerated degradation rate of AZ31 magnesium alloy by copper additions. Mater. Corros. 2018, 69, 760–769. [Google Scholar] [CrossRef]
- Tada, E.; Sugawara, K.; Kaneko, H. Distribution of pH during galvanic corrosion of a Zn/steel couple. Electrochim. Acta 2004, 49, 1019–1026. [Google Scholar] [CrossRef]
- United States Bureau of the Census. American Housing Survey, 2017 National Data. Available online: https://www.census.gov/programs-surveys/ahs/data/2017/ahs-2017-public-use-file--puf-/ahs-2017-national-public-use-file--puf-.html (accessed on 10 October 2020).
- Ewann, F.; Hoffman, P.S. Cysteine Metabolism in Legionella pneumophila: Characterization of an L-Cystine-Utilizing Mutant. Appl. Environ. Microbiol. 2006, 72, 3993–4000. [Google Scholar] [CrossRef] [Green Version]
- Shaheen, M.; Scott, C.; Ashbolt, N.J. Long-term persistence of infectious Legionella with free-living amoebae in drinking water biofilms. Int. J. Hyg. Environ. Health 2019, 222, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Richards, A.M.; Von Dwingelo, J.E.; Price, C.T.; Abu Kwaik, Y. Cellular microbiology and molecular ecology of Legionella–amoeba interaction. Virulence 2013, 4, 307–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, R.; Burghardt, H.; Bergmann, H. Acanthamoeba, naturally intracellularly infected with Pseudomonas aeruginosa, after their isolation from a microbiologically contaminated drinking water system in a hospital. Zent. Hyg. Umweltmed. 1995, 196, 532–544. [Google Scholar]
- Ji, P.; Rhoads, W.J.; Edwards, M.A.; Pruden, A. Impact of water heater temperature setting and water use frequency on the building plumbing microbiome. Isme J. 2017, 11, 1318–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buse, H.Y.; Lu, J.; Lu, X.; Mou, X.; Ashbolt, N.J. Microbial diversities (16S and 18S rRNA gene pyrosequencing) and environmental pathogens within drinking water biofilms grown on the common premise plumbing materials unplasticized polyvinylchloride and copper. FEMS Microbiol. Ecol. 2014, 88, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Falkinham, J.O.; Hilborn, E.D.; Arduino, M.J.; Pruden, A.; Edwards, M.A. Epidemiology and Ecology of Opportunistic Premise Plumbing Pathogens: Legionella pneumophila, Mycobacterium avium, and Pseudomonas aeruginosa. Environ. Health Perspect. 2015, 123, 749–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Pierre, D.; Bibby, K.; Stout, J.E. Bacterial community structure correlates with Legionella pneumophila colonization of New York City high rise building premises plumbing systems. Environ. Sci. Water Res. Technol. 2020, 6, 1324–1335. [Google Scholar] [CrossRef]
- Paranjape, K.; Bédard, É.; Whyte, L.G.; Ronholm, J.; Prévost, M.; Faucher, S.P. Presence of Legionella spp. in cooling towers: The role of microbial diversity, Pseudomonas, and continuous chlorine application. Water Res. 2020, 169, 115252. [Google Scholar] [CrossRef]
- Garner, E.; McLain, J.; Bowers, J.; Engelthaler, D.M.; Edwards, M.A.; Pruden, A. Microbial Ecology and Water Chemistry Impact Regrowth of Opportunistic Pathogens in Full-Scale Reclaimed Water Distribution Systems. Environ. Sci. Technol. 2018, 52, 9056–9068. [Google Scholar] [CrossRef]
- Ji, P.; Parks, J.; Edwards, M.A.; Pruden, A. Impact of Water Chemistry, Pipe Material and Stagnation on the Building Plumbing Microbiome. PLoS ONE 2015, 10, e0141087. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Yu, Z.; Chen, X.; Liu, R.; Zhang, H. Molecular characterization of natural biofilms from household taps with different materials: PVC, stainless steel, and cast iron in drinking water distribution system. Appl. Microbiol. Biotechnol. 2012, 97. [Google Scholar] [CrossRef] [PubMed]
- Proctor, C.R.; Gächter, M.; Kötzsch, S.; Rölli, F.; Sigrist, R.; Walser, J.-C.; Hammes, F. Biofilms in shower hoses–choice of pipe material influences bacterial growth and communities. Environ. Sci. Water Res. Technol. 2016, 2, 670–682. [Google Scholar] [CrossRef]
- Corre, M.-H.; Delafont, V.; Legrand, A.; Berjeaud, J.-M.; Verdon, J. Exploiting the Richness of Environmental Waterborne Bacterial Species to Find Natural Legionella pneumophila Competitors. Front. Microbiol. 2019, 9, 3360. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Tateda, K.; Ishii, Y.; Horikawa, M.; Miyairi, S.; Gotoh, N.; Ishiguro, M.; Yamaguchi, K. Pseudomonas aeruginosa Las quorum sensing autoinducer suppresses growth and biofilm production in Legionella species. Microbiology 2009, 155, 1934–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Declerck, P.; Behets, J.; Delaedt, Y.; Margineanu, A.; Lammertyn, E.; Ollevier, F. Impact of Non-Legionella Bacteria on the Uptake and Intracellular Replication of Legionella pneumophila in Acanthamoeba castellanii and Naegleria lovaniensis. Microb. Ecol. 2005, 50, 536–549. [Google Scholar] [CrossRef]
- Berry, D.; Horn, M.; Xi, C.; Raskin, L. Mycobacterium avium Infections of Acanthamoeba Strains: Host Strain Variability, Grazing-Acquired Infections, and Altered Dynamics of Inactivation with Monochloramine. Appl. Environ. Microbiol. 2010, 76, 6685–6688. [Google Scholar] [CrossRef] [Green Version]
- Weitere, M.; Bergfeld, T.; Rice, S.A.; Matz, C.; Kjelleberg, S. Grazing resistance of Pseudomonas aeruginosa biofilms depends on type of protective mechanism, developmental stage and protozoan feeding mode. Environ. Microbiol. 2005, 7, 1593–1601. [Google Scholar] [CrossRef]
- Matz, C.; Moreno, A.M.; Alhede, M.; Manefield, M.; Hauser, A.R.; Givskov, M.; Kjelleberg, S. Pseudomonas aeruginosa uses type III secretion system to kill biofilm-associated amoebae. ISME J. 2008, 2, 843–852. [Google Scholar] [CrossRef]
- Steinberg, C.; Laurent, J.; Edel-Hermann, V.; Barbezant, M.; Sixt, N.; Dalle, F.; Aho, S.; Bonnin, A.; Hartemann, P.; Sautour, M. Adaptation of Fusarium oxysporum and Fusarium dimerum to the specific aquatic environment provided by the water systems of hospitals. Water Res. 2015, 76, 53–65. [Google Scholar] [CrossRef]
- Petitjean, M.; Martak, D.; Silvant, A.; Bertrand, X.; Valot, B.; Hocquet, D. Genomic characterization of a local epidemic Pseudomonas aeruginosa reveals specific features of the widespread clone ST395. Microb. Genom. 2017, 3, e000129. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.D. Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 2010, 34, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, K.; Pruden, A.; Falkinham, J.O.; Edwards, M. Relationship between Organic Carbon and Opportunistic Pathogens in Simulated Glass Water Heaters. Pathogens 2015, 4, 355–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusserre, E.; Ginevra, C.; Hallier-Soulier, S.; Vandenesch, F.; Festoc, G.; Etienne, J.; Jarraud, S.; Molmeret, M. A PCR-based method for monitoring Legionella pneumophila in water samples detects viable but noncultivable legionellae that can recover their cultivability. Appl. Environ. Microbiol. 2008, 74, 4817–4824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Bédard, E.; Prévost, M.; Camper, A.K.; Hill, V.R.; Pruden, A. Methodological approaches for monitoring opportunistic pathogens in premise plumbing: A review. Water Res. 2017, 117, 68–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, J.L.; Niederweis, M. Resistance mechanisms of Mycobacterium tuberculosis against phagosomal copper overload. Tuberculosis 2012, 92, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Lehtola, M.J.; Miettinen, I.T.; Keinänen, M.M.; Kekki, T.K.; Laine, O.; Hirvonen, A.; Vartiainen, T.; Martikainen, P.J. Microbiology, chemistry and biofilm development in a pilot drinking water distribution system with copper and plastic pipes. Water Res. 2004, 38, 3769–3779. [Google Scholar] [CrossRef]
- Schwan, W.R.; Warrener, P.; Keunz, E.; Kendall Stover, C.; Folger, K.R. Mutations in the cueA gene encoding a copper homeostasis P-type ATPase reduce the pathogenicity of Pseudomonas aeruginosa in mice. Int. J. Med. Microbiol. 2005, 295, 237–242. [Google Scholar] [CrossRef]
- Abdollahi, S.; Rasooli, I.; Mousavi Gargari, S.L. The role of TonB-dependent copper receptor in virulence of Acinetobacter baumannii. Infect. Genet. Evol. 2018, 60, 181–190. [Google Scholar] [CrossRef]
- Alquethamy, S.; Khorvash, M.; Pederick, V.; Whittall, J.; Paton, J.; Paulsen, I.; Hassan, K.; McDevitt, C.; Eijkelkamp, B. The Role of the CopA Copper Efflux System in Acinetobacter baumannii Virulence. Int. J. Mol. Sci. 2019, 20, 575. [Google Scholar] [CrossRef] [Green Version]
- Huston, W.M.; Jennings, M.P.; McEwan, A.G. The multicopper oxidase of Pseudomonas aeruginosa is a ferroxidase with a central role in iron acquisition. Mol. Microbiol. 2002, 45, 1741–1750. [Google Scholar] [CrossRef] [Green Version]
- James, B.W.; Mauchline, W.S.; Fitzgeorge, R.B.; Dennis, P.J.; Keevil, C.W. Influence of iron-limited continuous culture on physiology and virulence of Legionella pneumophila. Infect. Immun. 1995, 63, 4224–4230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, P.N.; Koch, G.; Papaioannou, E.; Wahjudi, M.; Krzeslak, J.; Coenye, T.; Cool, R.H.; Quax, W.J. Role of PvdQ in Pseudomonas aeruginosa virulence under iron-limiting conditions. Microbiology 2010, 156, 49–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasil, M.L.; Ochsner, U.A. The response of Pseudomonas aeruginosa to iron: Genetics, biochemistry and virulence. Mol. Microbiol. 1999, 34, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Sokol, P.A.; Woods, D.E. Relationship of iron and extracellular virulence factors to Pseudomonas aeruginosa lung infections. J. Med. Microbiol. 1984, 18, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjorn, M.J.; Iglewski, B.H.; Ives, S.K.; Sadoff, J.C.; Vasil, M.L. Effect of iron on yields of exotoxin A in cultures of Pseudomonas aeruginosa PA-103. Infect. Immun. 1978, 19, 785–791. [Google Scholar] [CrossRef] [Green Version]
- Penwell, W.F.; Arivett, B.A.; Actis, L.A. The Acinetobacter baumannii entA Gene Located Outside the Acinetobactin Cluster Is Critical for Siderophore Production, Iron Acquisition and Virulence. PLoS ONE 2012, 7, e36493. [Google Scholar] [CrossRef] [Green Version]
- Fiester, S.E.; Arivett, B.A.; Schmidt, R.E.; Beckett, A.C.; Ticak, T.; Carrier, M.V.; Ghosh, R.; Ohneck, E.J.; Metz, M.L.; Sellin Jeffries, M.K.; et al. Iron-Regulated Phospholipase C Activity Contributes to the Cytolytic Activity and Virulence of Acinetobacter baumannii. PLoS ONE 2016, 11, e0167068. [Google Scholar] [CrossRef] [Green Version]
- Pruden, A. Balancing Water Sustainability and Public Health Goals in the Face of Growing Concerns about Antibiotic Resistance. Environ. Sci. Technol. 2014, 48, 5–14. [Google Scholar] [CrossRef]
- Berg, J.; Tom-Petersen, A.; Nybroe, O. Copper amendment of agricultural soil selects for bacterial antibiotic resistance in the field. Lett. Appl. Microbiol. 2005, 40, 146–151. [Google Scholar] [CrossRef]
- Hu, H.-W.; Wang, J.-T.; Li, J.; Li, J.-J.; Ma, Y.-B.; Chen, D.; He, J.-Z. Field-based evidence for copper contamination induced changes of antibiotic resistance in agricultural soils. Environ. Microbiol. 2016, 18, 3896–3909. [Google Scholar] [CrossRef]
- Knapp, C.W.; McCluskey, S.M.; Singh, B.K.; Campbell, C.D.; Hudson, G.; Graham, D.W. Antibiotic Resistance Gene Abundances Correlate with Metal and Geochemical Conditions in Archived Scottish Soils. PLoS ONE 2011, 6, e27300. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.; Thorsen, M.K.; Holm, P.E.; Jensen, J.; Nybroe, O.; Brandt, K.K. Cu Exposure under Field Conditions Coselects for Antibiotic Resistance as Determined by a Novel Cultivation-Independent Bacterial Community Tolerance Assay. Environ. Sci. Technol. 2010, 44, 8724–8728. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.W.; Olivares-Rieumont, S.; Knapp, C.W.; Lima, L.; Werner, D.; Bowen, E. Antibiotic Resistance Gene Abundances Associated with Waste Discharges to the Almendares River near Havana, Cuba. Environ. Sci. Technol. 2011, 45, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Chen, L.; Ye, C.; Yu, X. Co-selection of antibiotic resistance via copper shock loading on bacteria from a drinking water bio-filter. Environ. Pollut. 2018, 233, 132–141. [Google Scholar] [CrossRef]
- Becerra-Castro, C.; Machado, R.; Vaz-Moreira, I.; Manaia, C. Assessment of copper and zinc salts as selectors of antibiotic resistance in Gram-negative bacteria. Sci. Total Environ. 2015, 530. [Google Scholar] [CrossRef]
- Barker, J.; Scaife, H.; Brown, M.R. Intraphagocytic growth induces an antibiotic-resistant phenotype of Legionella pneumophila. Antimicrob. Agents Chemother. 1995, 39, 2684. [Google Scholar] [CrossRef] [Green Version]
- Oglesby-Sherrouse, A.G.; Djapgne, L.; Nguyen, A.T.; Vasil, A.I.; Vasil, M.L. The complex interplay of iron, biofilm formation, and mucoidy affecting antimicrobial resistance of Pseudomonas aeruginosa. Pathog. Dis. 2014, 70, 307–320. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Liu, S.; Wang, Z.; Zhang, X.; Li, M.; Wu, B. Metagenomic profiles and antibiotic resistance genes in gut microbiota of mice exposed to arsenic and iron. Chemosphere 2014, 112, 1–8. [Google Scholar] [CrossRef]
- European Commisssion. Council Directive 98/83/EC of 3 November 1998 on the quality of water intended for human consumption. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A31998L0083 (accessed on 10 November 2020).





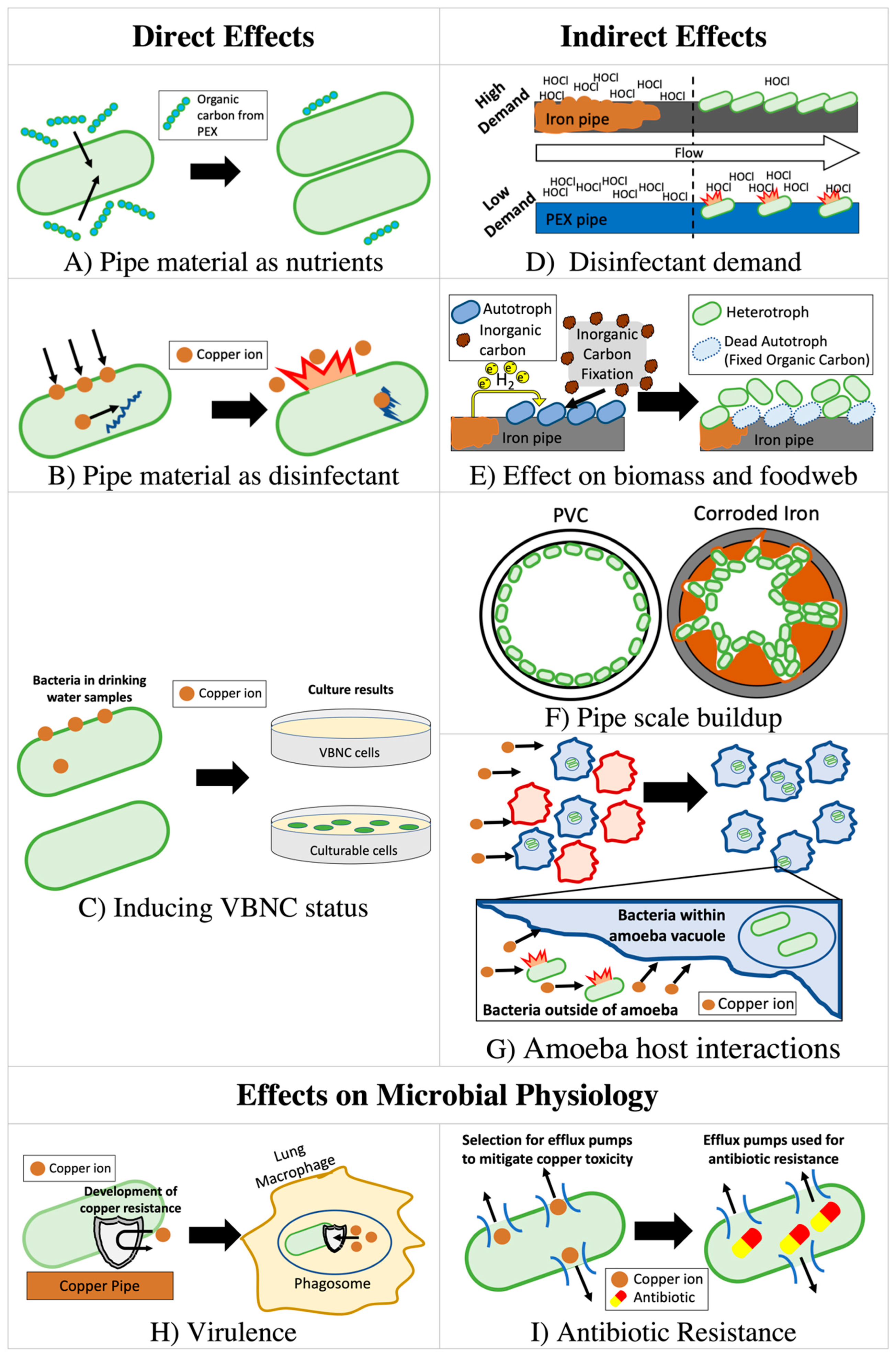{kind=link}
{kind=link}
{kind=link}
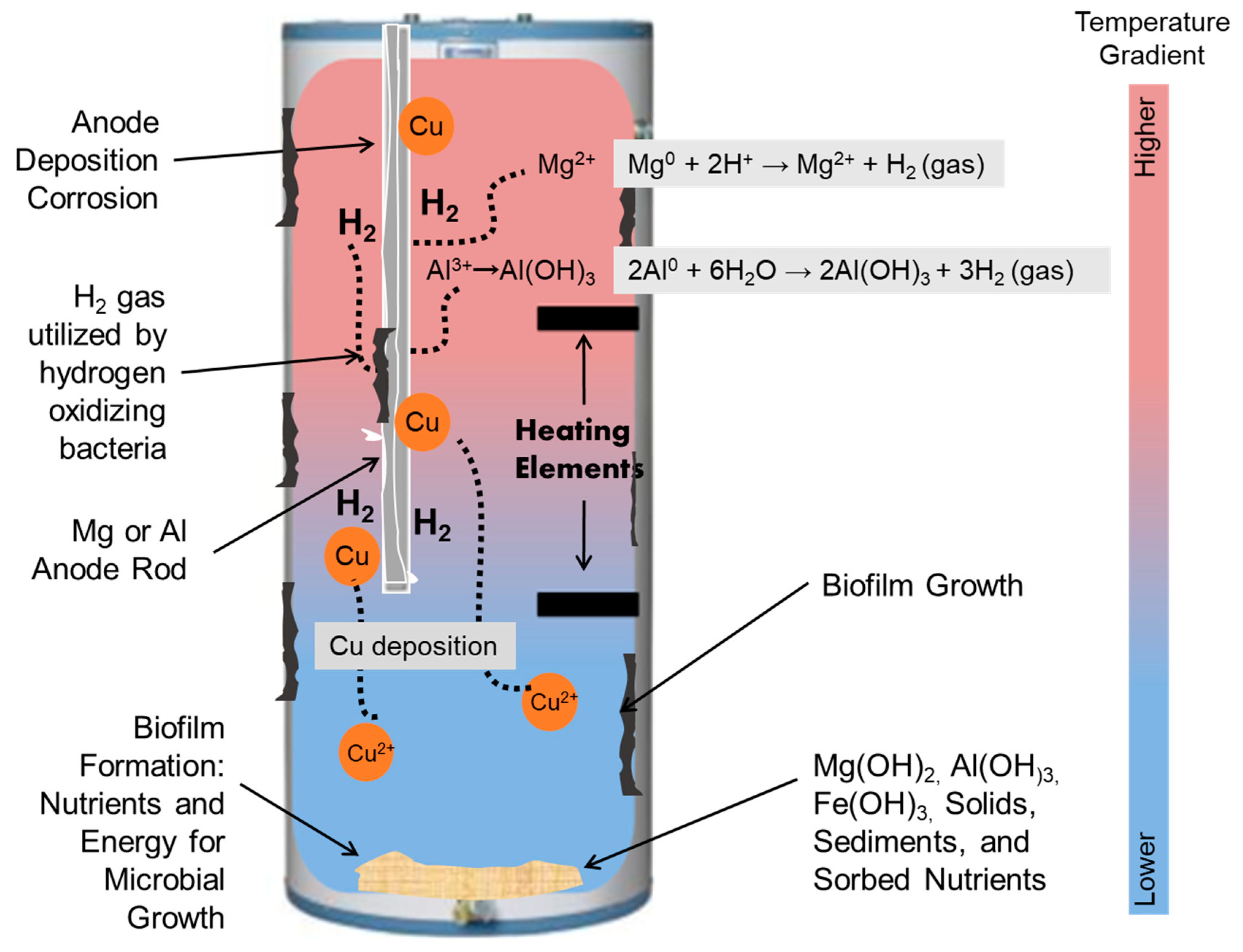{kind=link}
| Water Chemistry Attribute Influenced by Pipe Materials | Relevance to OPs | Effect of Pipe Materials on OPs Control as Mediated by the Indicated Water Chemistry Attribute | ||||
|---|---|---|---|---|---|---|
| Copper | PVC | PEX | SS | Iron 1 | ||
| Chlorine | Disinfectant | - [43] | 0 [43,44,45,46,47,48,49,50] | - [43,51,52] | 0/- [43,44,45,48,53] | -- [43,44,45,46,47,48] |
| Chloramine | Disinfectant | - [43,54] | 0 [43,50] | 0 [43,52] | 0 [43] | -- [43,55,56] |
| Assimilable Organic Carbon | Carbon source | 0 | - [42,56,57] | -- [42,56,58,59] | 0 | 0 |
| Hydrogen Gas (aq) | Food web | 0 | 0 | 0 | 0 | - [60,61] |
| Release of Metals | Release of metals | +/- [59,62,63,64] | 0 | 0 | 0 [65] | -- [66] |
| Opportunistic Pathogen | Associated Diseases | Exposure Route(s) | Inactivation via Copper | Growth via Copper | ||
|---|---|---|---|---|---|---|
| Antimicrobial Efficacy * | Evidence for Cu-Induced VBNC | Micronutrient Activity | Amoeba-Mediated Growth | |||
| Amoebae | Encephalitis, Eye infections, Primary amebic meningoencephalitis [85,86,87] | Dermal, Inhalation, [85,86,87] | Moderate to Somewhat inhibited [59,88] | Unknown and unlikely | Possible that organisms are copper deficient and additional copper could increase growth [59,80] | NA |
| Acinetobacter baumannii | Bacteremia, Meningitis, Pneumonia, Urinary tract infections [89] | Dermal, Inhalation [89] | Moderate to Somewhat inhibited [58,90,91,92] | Unknown | Yes [93,94] | |
| Staphylococcus aureus | Bacteremia, Endocarditis, Osteomyelitis, Pneumonia, Sepsis, Skin infections [95] | Dermal, Inhalation [96,97] | Moderate [90,98,99] | Unknown | Yes [100,101] | |
| Stenotrophomonas maltophilia | Bacteremia, Endocarditis, Eye infections, Meningitis, Pneumonia, Sepsis, Skin infections, Urinary tract infections [102,103] | Dermal, Inhalation [102,103] | Moderate [91,92] | Limited [104] | Yes [105] | |
| Nontuberculous Mycobacteria (NTM): Mycobacterium avium complex; Mycobacterium abscessus complex; Mycobacterium kansasii and other species | Bacteremia, Pneumonia, Skin infections [106] | Dermal, Ingestion, Inhalation [107] | Moderate [108,109,110,111] | Limited [109] | Yes [36,112] | |
| Aeromonas hydrophila | Gastroenteritis, Meningitis, Peritonitis, Pneumonia, Skin infections [113] | Ingestion, Inhalation [113] | Unknown [114] | Unknown | Yes [115,116] | |
| Legionella pneumophila | Legionnaires’ disease, Pontiac fever [117] | Inhalation [118] | Somewhat inhibited to High [62,83,119,120] | Moderate [121,122] | Yes [123,124] | |
| Pseudomonas aeruginosa | Bacteremia, Endocarditis, Eye infections, Gastroenteritis, Osteomyelitis, Pneumonia, Sepsis, Skin infections, Urinary tract infections [125] | Dermal, Ingestion, Inhalation [125,126] | Somewhat inhibited to High [90,91,92,98,99,127,128,129,130,131,132] | Strong [127,132,133] | Yes [36,134] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cullom, A.C.; Martin, R.L.; Song, Y.; Williams, K.; Williams, A.; Pruden, A.; Edwards, M.A. Critical Review: Propensity of Premise Plumbing Pipe Materials to Enhance or Diminish Growth of Legionella and Other Opportunistic Pathogens. Pathogens 2020, 9, 957. https://doi.org/10.3390/pathogens9110957
Cullom AC, Martin RL, Song Y, Williams K, Williams A, Pruden A, Edwards MA. Critical Review: Propensity of Premise Plumbing Pipe Materials to Enhance or Diminish Growth of Legionella and Other Opportunistic Pathogens. Pathogens. 2020; 9(11):957. https://doi.org/10.3390/pathogens9110957
Chicago/Turabian StyleCullom, Abraham C., Rebekah L. Martin, Yang Song, Krista Williams, Amanda Williams, Amy Pruden, and Marc A. Edwards. 2020. "Critical Review: Propensity of Premise Plumbing Pipe Materials to Enhance or Diminish Growth of Legionella and Other Opportunistic Pathogens" Pathogens 9, no. 11: 957. https://doi.org/10.3390/pathogens9110957