Ursolic Acid Potentializes Conventional Therapy in Experimental Leishmaniasis

 ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Chemical Analysis of Ursolic Acid
2.2. Evaluation of the Therapeutic Potential of Drug Association in Experimental Visceral Leishmaniasis
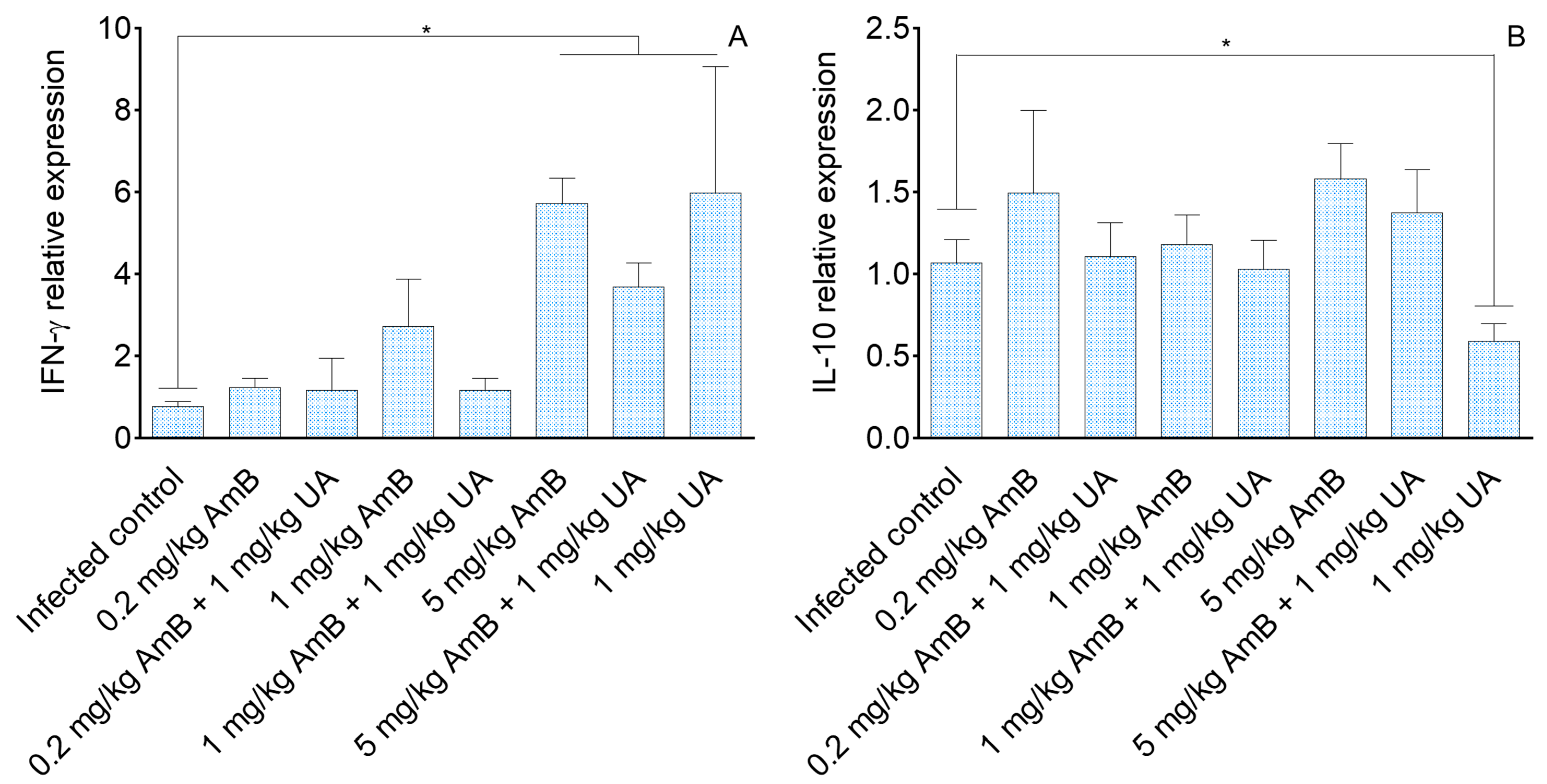
2.3. Analysis of Cellular Immune Response in Visceral Leishmaniasis
2.4. Evaluation of the Therapeutic Potential of Drug Association in Experimental Cutaneous Leishmaniasis
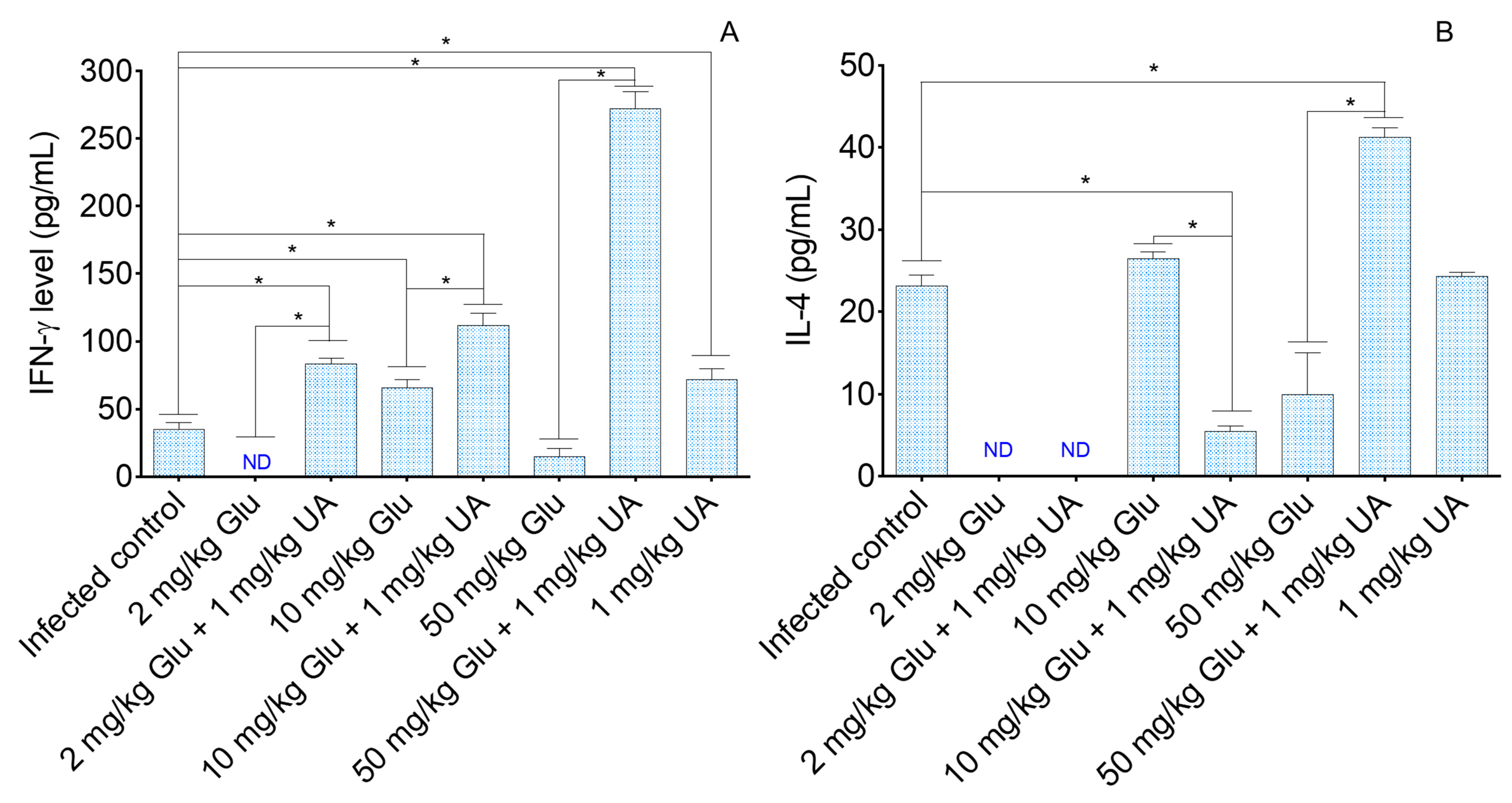
2.5. Analysis of Cellular Immune Response in Experimental Cutaneous Leishmaniasis
3. Discussion
4. Materials and Methods
4.1. Drugs
4.2. Chemical Analysis of Ursolic Acid
4.3. Experimental Animals and Ethical Considerations
4.4. Parasites
4.5. Evaluation of the Therapeutic Potential of Drug Associations in Visceral and Cutaneous Leishmaniasis
4.6. Determination of Parasite Load
4.7. Analysis of Cellular Immune Response in VL and CL
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO Leishmaniasis. Available online: https://www.who.int/leishmaniasis/en/ (accessed on 3 July 2019).
- Stockdale, L.; Newton, R. A review of preventative methods against human leishmaniasis infection. PLoS Negl. Trop. Dis. 2013, 7, e2278. [Google Scholar] [CrossRef] [PubMed]
- PAHO/OMS. Leishmanioses: Informe Epidemiológico nas Américas: Washington: Organização Pan-Americana da Saúde. Available online: www.paho.org/leishmaniasis (accessed on 10 December 2019).
- Torres-Guerrero, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J.; Arenas, R. Leishmaniasis: A review. F1000Research 2017, 6, 750. [Google Scholar] [CrossRef] [PubMed]
- Soyer, T.G.; Mendonça, D.V.C.; Tavares, G.S.V.; Lage, D.P.; Dias, D.S.; Ribeiro, P.A.F.; Perin, L.; Ludolf, F.; Coelho, V.T.S.; Ferreira, A.C.G.; et al. Evaluation of the in vitro and in vivo antileishmanial activity of a chloroquinolin derivative against Leishmania species capable of causing tegumentary and visceral leishmaniasis. Exp. Parasitol. 2019, 199, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Sundar, S.; Chakravarty, J.; Meena, L.P. Leishmaniasis: Treatment, drug resistance and emerging therapies. Expert Opin. Orphan Drugs 2019, 7, 1–10. [Google Scholar] [CrossRef]
- Dorlo, T.P.C.; Balasegaram, M.; Beijnen, J.H.; de Vries, P.J. Miltefosine: A review of its pharmacology and therapeutic efficacy in the treatment of leishmaniasis. J. Antimicrob. Chemother. 2012, 67, 2576–2597. [Google Scholar] [CrossRef]
- Chakravarty, J.; Sundar, S. Current and emerging medications for the treatment of leishmaniasis. Expert Opin. Pharmacother. 2019, 20, 1251–1265. [Google Scholar] [CrossRef]
- Uliana, S.R.B.; Trinconi, C.T.; Coelho, A.C. Chemotherapy of leishmaniasis: Present challenges. Parasitology 2018, 145, 464–480. [Google Scholar] [CrossRef] [PubMed]
- Van Griensven, J.; Balasegaram, M.; Meheus, F.; Alvar, J.; Lynen, L.; Boelaert, M. Combination therapy for visceral leishmaniasis. Lancet Infect. Dis. 2010, 10, 184–194. [Google Scholar] [CrossRef]
- Pastor, J.; García, M.; Steinbauer, S.; Setzer, W.N.; Scull, R.; Gille, L.; Monzote, L. Combinations of ascaridole, carvacrol, and caryophyllene oxide against Leishmania. Acta Trop. 2015, 145, 31–38. [Google Scholar] [CrossRef]
- Kremsner, P.G.; Krishna, S. Antimalarial combinations. Lancet 2004, 364, 285–294. [Google Scholar] [CrossRef]
- Rodrigues-Junior, V.S.; Villela, A.D.; Abbadi, B.L.; Sperotto, N.D.M.; Pissinate, K.; Picada, J.N.; Bondan da Silva, J.; Bizarro, C.V.; Machado, P.; Basso, L.A. Nonclinical evaluation of IQG-607, an anti-tuberculosis candidate with potential use in combination drug therapy. Regul. Toxicol. Pharmacol. 2020, 111, 104553. [Google Scholar] [CrossRef]
- Ghorbani, M.; Farhoudi, R. Leishmaniasis in humans: Drug or vaccine therapy? Drug Des. Devel. Ther. 2017, 12, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Sundar, S.; Sinha, P.K.; Rai, M.; Verma, D.K.; Nawin, K.; Alam, S.; Chakravarty, J.; Vaillant, M.; Verma, N.; Pandey, K.; et al. Comparison of short-course multidrug treatment with standard therapy for visceral leishmaniasis in India: An open-label, non-inferiority, randomised controlled trial. Lancet 2011, 377, 477–486. [Google Scholar] [CrossRef]
- Sundar, S.; Rai, M.; Chakravarty, J.; Agarwal, D.; Agrawal, N.; Vaillant, M.; Olliaro, P.; Murray, H.W. New Treatment Approach in Indian Visceral Leishmaniasis: Single-Dose Liposomal Amphotericin B Followed by Short-Course Oral Miltefosine. Clin. Infect. Dis. 2008, 47, 1000–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezaei Riabi, T.; Sharifi, I.; Miramin Mohammadi, A.; Khamesipour, A.; Hakimi Parizi, M. Evaluation of a Possible Synergistic Effect of Meglumine Antimoniate with Paromomycin, Miltefosine or Allopurinol on in Vitro Susceptibility of Leishmania tropica Resistant Isolate. Iran. J. Parasitol. 2013, 8, 396–401. [Google Scholar] [PubMed]
- Passero, L.F.D.; Cruz, L.A.; Santos-Gomes, G.; Rodrigues, E.; Laurenti, M.D.; Lago, J.H.G. Conventional versus natural alternative treatments for leishmaniasis: A review. Curr. Top. Med. Chem. 2018. [Google Scholar] [CrossRef]
- Kashyap, D.; Tuli, H.S.; Sharma, A.K. Ursolic acid (UA): A metabolite with promising therapeutic potential. Life Sci. 2016, 146, 201–213. [Google Scholar] [CrossRef]
- Chen, H.; Gao, Y.; Wang, A.; Zhou, X.; Zheng, Y.; Zhou, J. Evolution in medicinal chemistry of ursolic acid derivatives as anticancer agents. Eur. J. Med. Chem. 2015, 92, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Jesus, J.A.; Fragoso, T.N.; Yamamoto, E.S.; Laurenti, M.D.; Silva, M.S.; Ferreira, A.F.; Lago, J.H.G.; Santos-Gomes, G.; Passero, L.F.D. Corrigendum to “Therapeutic effect of ursolic acid in experimental visceral leishmaniasis”. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 250. [Google Scholar] [CrossRef]
- Yamamoto, E.S.; Campos, B.L.S.; Jesus, J.A.; Laurenti, M.D.; Ribeiro, S.P.; Kallás, E.G.; Rafael-Fernandes, M.; Santos-Gomes, G.; Silva, M.S.; Sessa, D.P.; et al. The Effect of Ursolic Acid on Leishmania (Leishmania) amazonensis Is Related to Programed Cell Death and Presents Therapeutic Potential in Experimental Cutaneous Leishmaniasis. PLoS ONE 2015, 10, e0144946. [Google Scholar] [CrossRef] [Green Version]
- Kazakova, O.B.; Giniyatullina, G.V.; Yamansarov, E.Y.; Tolstikov, G.A. Betulin and ursolic acid synthetic derivatives as inhibitors of Papilloma virus. Bioorg. Med. Chem. Lett. 2010, 20, 4088–4090. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-X.; Lin, C.-J.; Jia, Z.-J. Triterpenoids and Sesquiterpenes from Mulgedium tataricum. Planta Med. 2006, 72, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Begum, S.; Ayub, A.; Qamar Zehra, S.; Shaheen Siddiqui, B.; Iqbal Choudhary, M.; Samreen. Leishmanicidal Triterpenes from Lantana camara. Chem. Biodivers. 2014, 11, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, E.S.; Campos, B.L.S.; Laurenti, M.D.; Lago, J.H.G.; Grecco, S.D.S.; Corbett, C.E.P.; Passero, L.F.D. Treatment with triterpenic fraction purified from Baccharis uncinella leaves inhibits Leishmania (Leishmania) amazonensis spreading and improves Th1 immune response in infected mice. Parasitol. Res. 2014, 113, 333–339. [Google Scholar] [CrossRef]
- Mahato, S.B.; Kundu, A.P. 13C NMR Spectra of pentacyclic triterpenoids—A compilation and some salient features. Phytochemistry 1994, 37, 1517–1575. [Google Scholar] [CrossRef]
- Hou, W.; Li, Y.; Zhang, Q.; Wei, X.; Peng, A.; Chen, L.; Wei, Y. Triterpene acids isolated from Lagerstroemia speciosa leaves as α -glucosidase inhibitors. Phyther. Res. 2009, 23, 614–618. [Google Scholar] [CrossRef]
- Verdi, L.G.; Brighente, I.M.C.; Pizzolatti, M.G. Gênero Baccharis (Asteraceae): Aspectos químicos, econômicos e biológicos. Quim. Nova 2005, 28, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Mostafavi, M.; Sharifi, I.; Farajzadeh, S.; Khazaeli, P.; Sharifi, H.; Pourseyedi, E.; Kakooei, S.; Bamorovat, M.; Keyhani, A.; Parizi, M.H.; et al. Niosomal formulation of amphotericin B alone and in combination with glucantime: In vitro and in vivo leishmanicidal effects. Biomed. Pharmacother. 2019, 116, 108942. [Google Scholar] [CrossRef]
- Yeon Kang, J.; Gao, J.; Hwan Shin, D.; Alvarez, C.; Zhong, W.; Kwon, G. Pharmacokinetics and Renal Toxicity of Monomeric Amphotericin B in Rats after a Multiple Dose Regimen. Pharm. Nanotechnol. 2016, 4, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Trinconi, C.T.; Reimão, J.Q.; Coelho, A.C.; Uliana, S.R.B. Efficacy of tamoxifen and miltefosine combined therapy for cutaneous leishmaniasis in the murine model of infection with Leishmania amazonensis. J. Antimicrob. Chemother. 2016, 71, 1314–1322. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.F.; Schubach, A.O.; Martins, M.M.; Passos, S.L.; Oliveira, R.V.; Marzochi, M.C.; Andrade, C.A. Systematic review of the adverse effects of cutaneous leishmaniasis treatment in the New World. Acta Trop. 2011, 118, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.I.S.; Arruda, V.O.; Alves, E.V.C.; de Azevedo, A.P.S.; Monteiro, S.G.; Pereira, S.R.F. Genotoxic effects of the antileishmanial drug glucantime®. Arch. Toxicol. 2010, 84, 227–232. [Google Scholar] [CrossRef]
- Moreira, V.R.; de Jesus, L.C.L.; Soares, R.-E.P.; Silva, L.D.M.; Pinto, B.A.S.; Melo, M.N.; Paes, A.M.D.A.; Pereira, S.R.F. Meglumine Antimoniate (Glucantime) Causes Oxidative Stress-Derived DNA Damage in BALB/c Mice Infected by Leishmania (Leishmania) infantum. Antimicrob. Ag. Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunn, P.T.; Montes de Oca, M.; de Labastida Rivera, F.; Kumar, R.; Ng, S.S.; Edwards, C.L.; Faleiro, R.J.; Sheel, M.; Amante, F.H.; Frame, T.C.M.; et al. Distinct Roles for CD4 + Foxp3 + Regulatory T Cells and IL-10–Mediated Immunoregulatory Mechanisms during Experimental Visceral Leishmaniasis Caused by Leishmania donovani. J. Immunol. 2018, 201, 3362–3372. [Google Scholar] [CrossRef] [Green Version]
- Dayakar, A.; Chandrasekaran, S.; Kuchipudi, S.V.; Kalangi, S.K. Cytokines: Key Determinants of Resistance or Disease Progression in Visceral Leishmaniasis: Opportunities for Novel Diagnostics and Immunotherapy. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banchereau, J.; Pascual, V.; O’Garra, A. From IL-2 to IL-37: The expanding spectrum of anti-inflammatory cytokines. Nat. Immunol. 2012, 13, 925–931. [Google Scholar] [CrossRef]
- Kima, P.E.; Soong, L. Interferon Gamma in Leishmaniasis. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Hamill, R.J. Amphotericin B Formulations: A Comparative Review of Efficacy and Toxicity. Drugs 2013, 73, 919–934. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hoffmann, K.F.; Mendez, S.; Kamhawi, S.; Udey, M.C.; Wynn, T.A.; Sacks, D.L. The Role of Interleukin (IL)-10 in the Persistence of Leishmania major in the Skin after Healing and the Therapeutic Potential of Anti–IL-10 Receptor Antibody for Sterile Cure. J. Exp. Med. 2001, 194, 1497–1506. [Google Scholar] [CrossRef] [Green Version]
- Sacks, D.; Noben-Trauth, N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat. Rev. Immunol. 2002, 2, 845–858. [Google Scholar] [CrossRef]
- Castellano, L.R.; Filho, D.C.; Argiro, L.; Dessein, H.; Prata, A.; Dessein, A.; Rodrigues, V. Th1/Th2 immune responses are associated with active cutaneous leishmaniasis and clinical cure is associated with strong interferon-γ production. Hum. Immunol. 2009, 70, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Himmelrich, H.; Launois, P.; Maillard, I.; Biedermann, T.; Tacchini-Cottier, F.; Locksley, R.M.; Röcken, M.; Louis, J.A. In BALB/c Mice, IL-4 Production During the Initial Phase of Infection with Leishmania major Is Necessary and Sufficient to Instruct Th2 Cell Development Resulting in Progressive Disease. J. Immunol. 2000, 164, 4819–4825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corral, M.J.; Serrano, D.R.; Moreno, I.; Torrado, J.J.; Dominguez, M.; Alunda, J.M. Efficacy of low doses of amphotericin B plus allicin against experimental visceral leishmaniasis. J. Antimicrob. Chemother. 2014, 69, 3268–3274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, I.A.C.; de Paula, R.C.; Alves, C.L.; Faria, K.F.; de Oliveira, M.M.; Mendes, G.G.; Dias, E.M.F.A.; Ribeiro, R.R.; de Oliveira, A.B.; da Silva, S.M. Efficacy of lapachol on treatment of cutaneous and visceral leishmaniasis. Exp. Parasitol. 2019, 199, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Godinho, J.L.P.; Simas-Rodrigues, C.; Silva, R.; Ürmenyi, T.P.; de Souza, W.; Rodrigues, J.C.F. Efficacy of miltefosine treatment in Leishmania amazonensis-infected BALB/c mice. Int. J. Antimicrob. Ag. 2012, 39, 326–331. [Google Scholar] [CrossRef] [Green Version]
- Fernández-García, R.; Lalatsa, A.; Statts, L.; Bolás-Fernández, F.; Ballesteros, M.P.; Serrano, D.R. Transferosomes as nanocarriers for drugs across the skin: Quality by design from lab to industrial scale. Int. J. Pharm. 2020, 573, 118817. [Google Scholar] [CrossRef]
- Lalatsa, A.; Statts, L.; Adriana de Jesus, J.; Adewusi, O.; Auxiliadora Dea-Ayuela, M.; Bolas-Fernandez, F.; Dalastra Laurenti, M.; Felipe Domingues Passero, L.; Serrano, D.R. Topical buparvaquone nano-enabled hydrogels for cutaneous leishmaniasis. Int. J. Pharm. 2020, 588, 119734. [Google Scholar] [CrossRef]
- Henao, H.H.; Osorio, Y.; Saravia, N.G.; Gómez, A.; Travi, B. Efficacy and toxicity of pentavalent antimonials (Glucantime and Pentostam) in an American cutaneous leishmaniasis animal model: Luminometry application. Biomedica 2004, 24, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Buffet, P.A.; Garin, Y.J.F.; Sulahian, A.; Nassar, N.; Derouin, F. Therapeutic effect of reference antileishmanial agents in murine visceral leishmaniasis due to Leishmania infantum. Ann. Trop. Med. Parasitol. 1996, 90, 295–302. [Google Scholar] [CrossRef]
- Bordon, M.L.A.C.; Laurenti, M.D.; Ribeiro, S.P.; Toyama, M.H.; Toyama, D.D.O.; Passero, L.F.D. Effect of phospholipase A2 inhibitors during infection caused by Leishmania (Leishmania) amazonensis. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 21. [Google Scholar] [CrossRef]
- Lafuse, W.P.; Story, R.; Mahylis, J.; Gupta, G.; Varikuti, S.; Steinkamp, H.; Oghumu, S.; Satoskar, A.R. Leishmania donovani Infection Induces Anemia in Hamsters by Differentially Altering Erythropoiesis in Bone Marrow and Spleen. PLoS ONE 2013, 8, e59509. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jesus, J.A.; Fragoso da Silva, T.N.; Yamamoto, E.S.; G. Lago, J.H.; Dalastra Laurenti, M.; Passero, L.F.D. Ursolic Acid Potentializes Conventional Therapy in Experimental Leishmaniasis. Pathogens 2020, 9, 855. https://doi.org/10.3390/pathogens9100855
Jesus JA, Fragoso da Silva TN, Yamamoto ES, G. Lago JH, Dalastra Laurenti M, Passero LFD. Ursolic Acid Potentializes Conventional Therapy in Experimental Leishmaniasis. Pathogens. 2020; 9(10):855. https://doi.org/10.3390/pathogens9100855
Chicago/Turabian StyleJesus, Jéssica Adriana, Thays Nicolli Fragoso da Silva, Eduardo Seiji Yamamoto, João Henrique G. Lago, Márcia Dalastra Laurenti, and Luiz Felipe Domingues Passero. 2020. "Ursolic Acid Potentializes Conventional Therapy in Experimental Leishmaniasis" Pathogens 9, no. 10: 855. https://doi.org/10.3390/pathogens9100855