Enterococcus faecalis Polymicrobial Interactions Facilitate Biofilm Formation, Antibiotic Recalcitrance, and Persistent Colonization of the Catheterized Urinary Tract

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Results
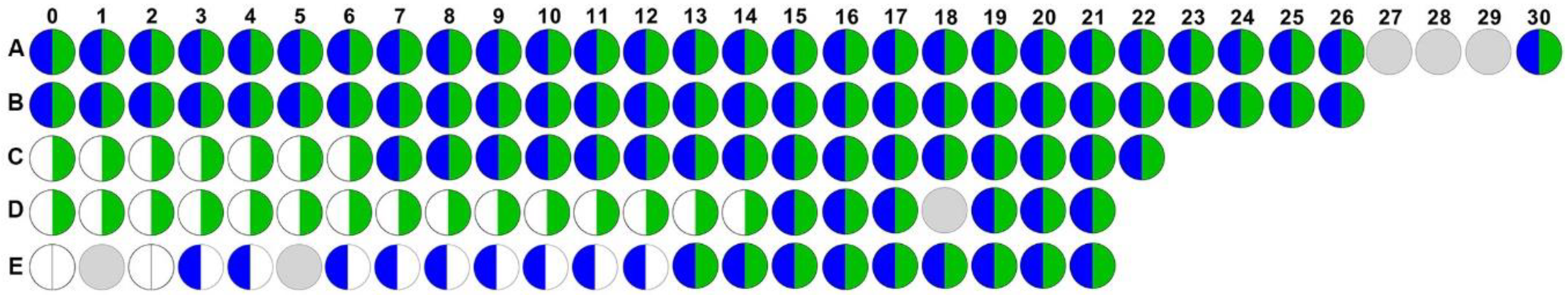
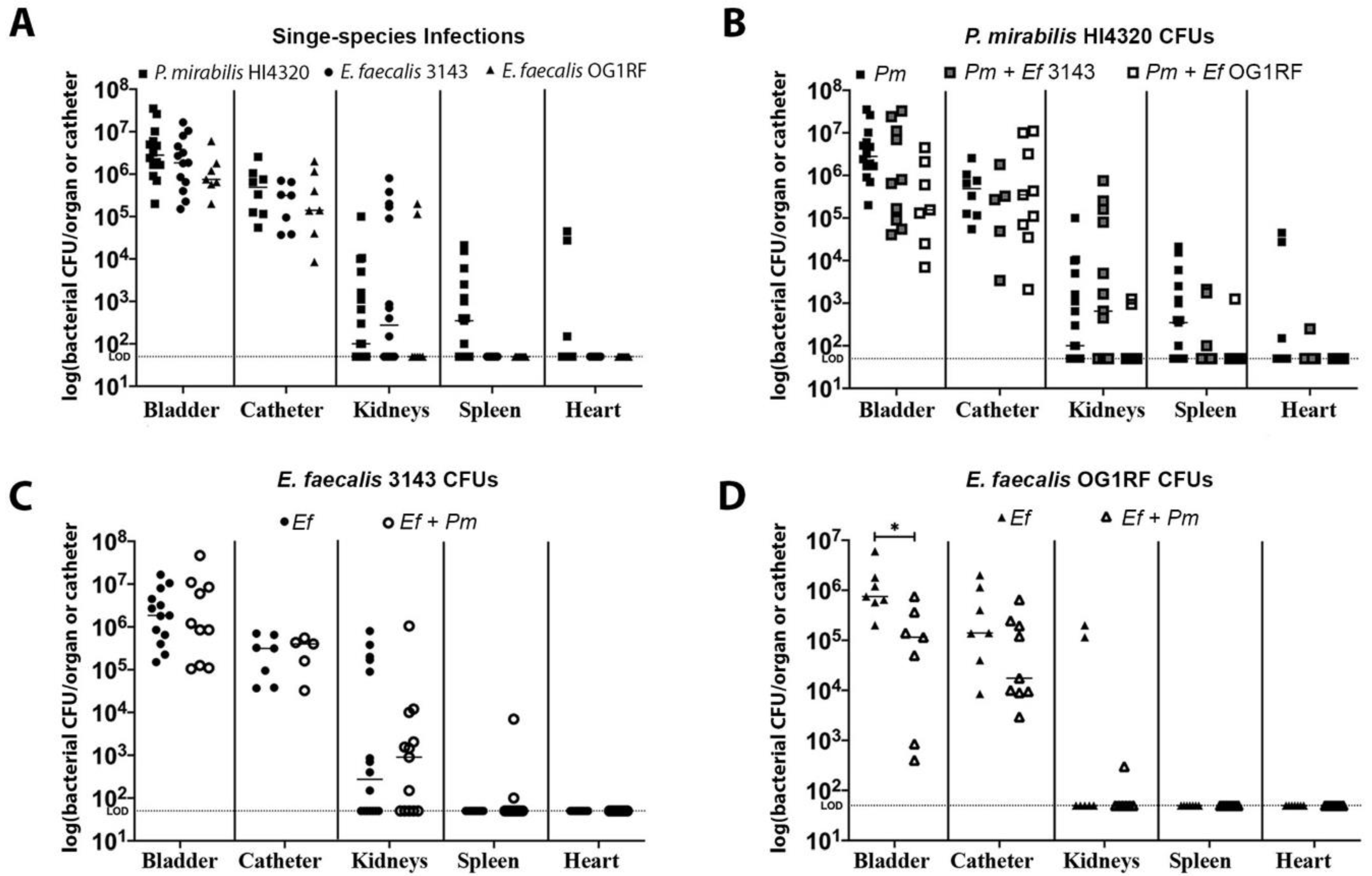
2.1. Proteus mirabilis and Enterococcus faecalis Persistently Co-colonize in Catheterized Nursing Home Residents
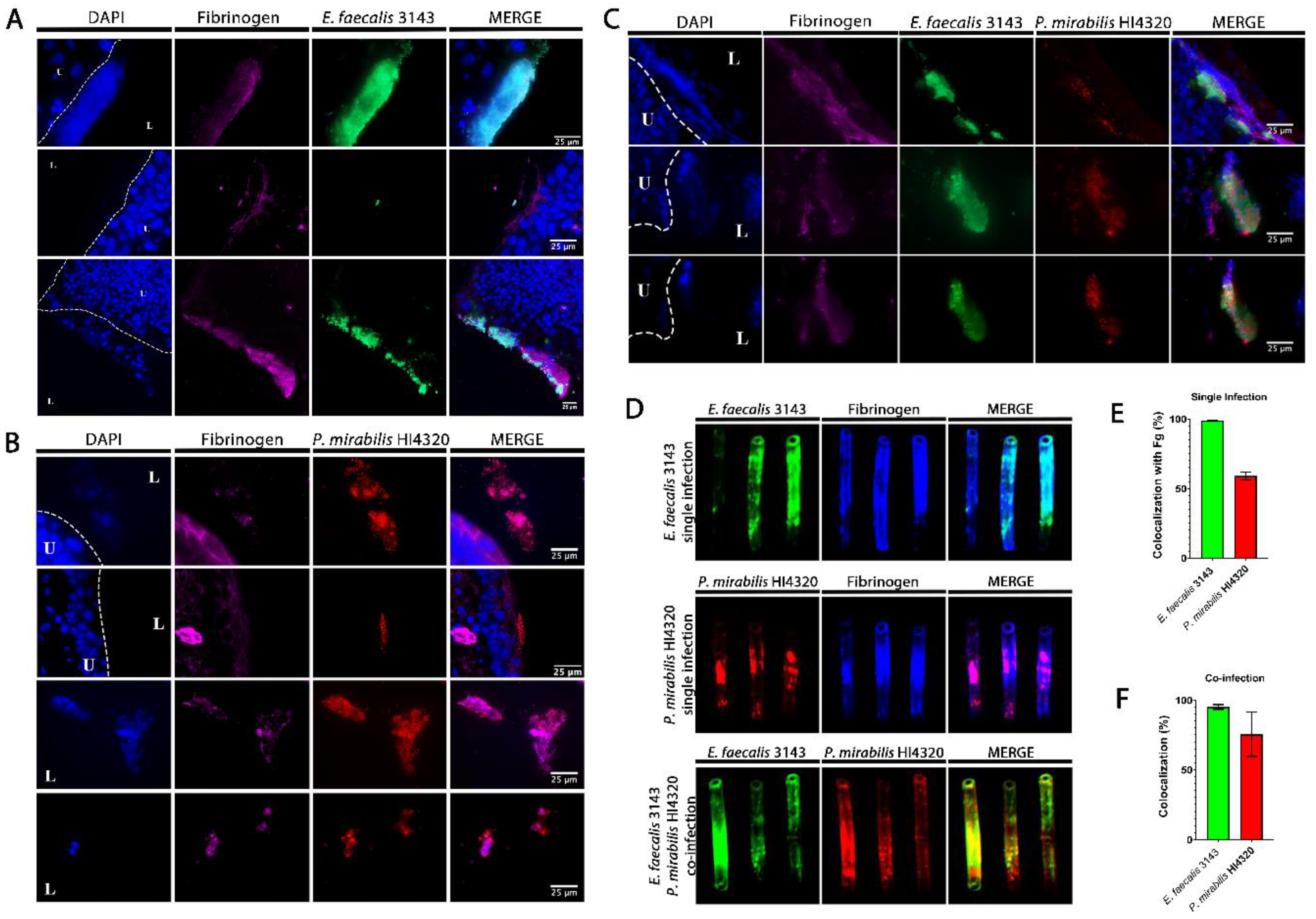
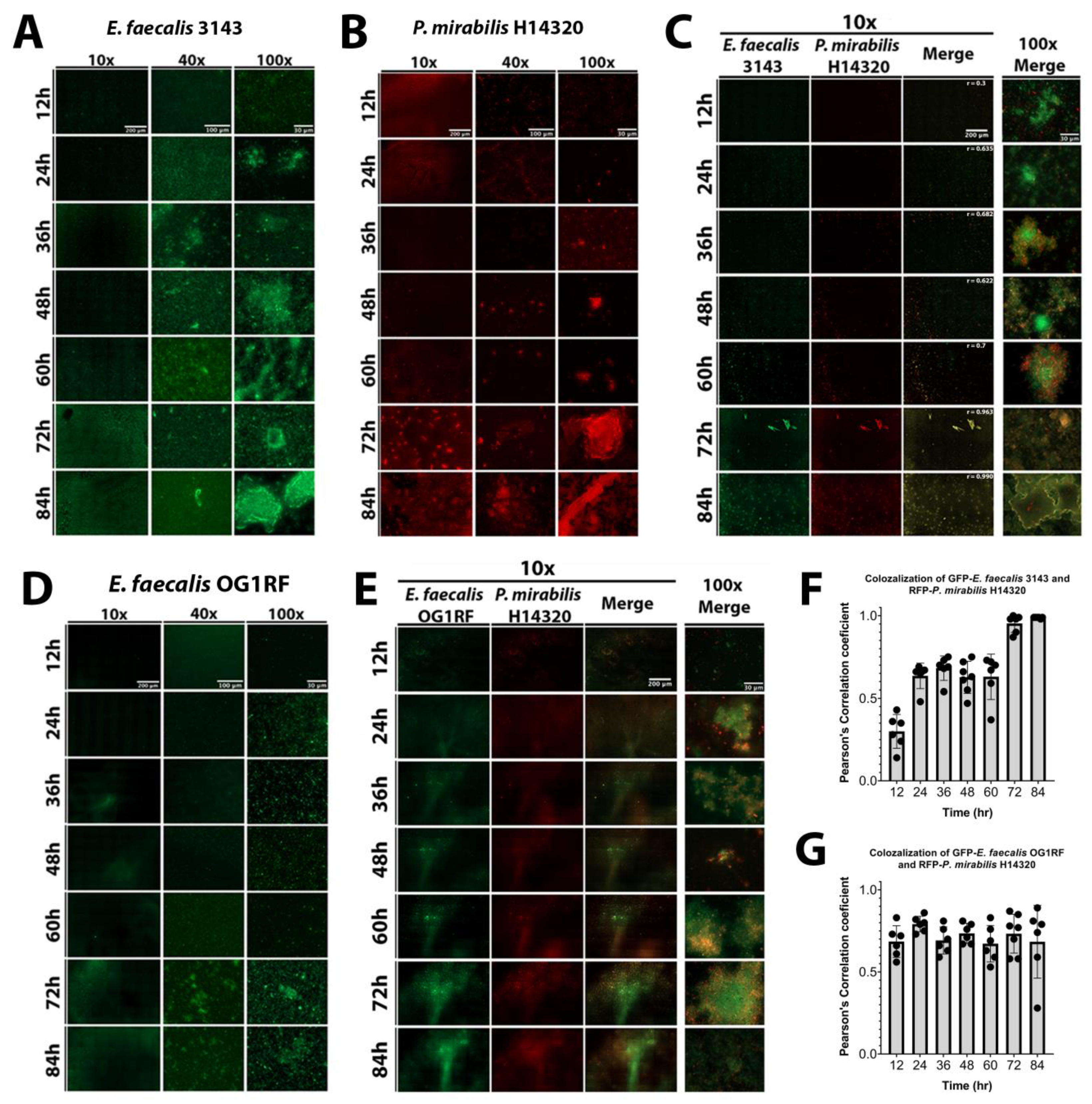
2.2. Enterococcus faecalis and Proteus mirabilis Co-localize during Experimental CAUTI
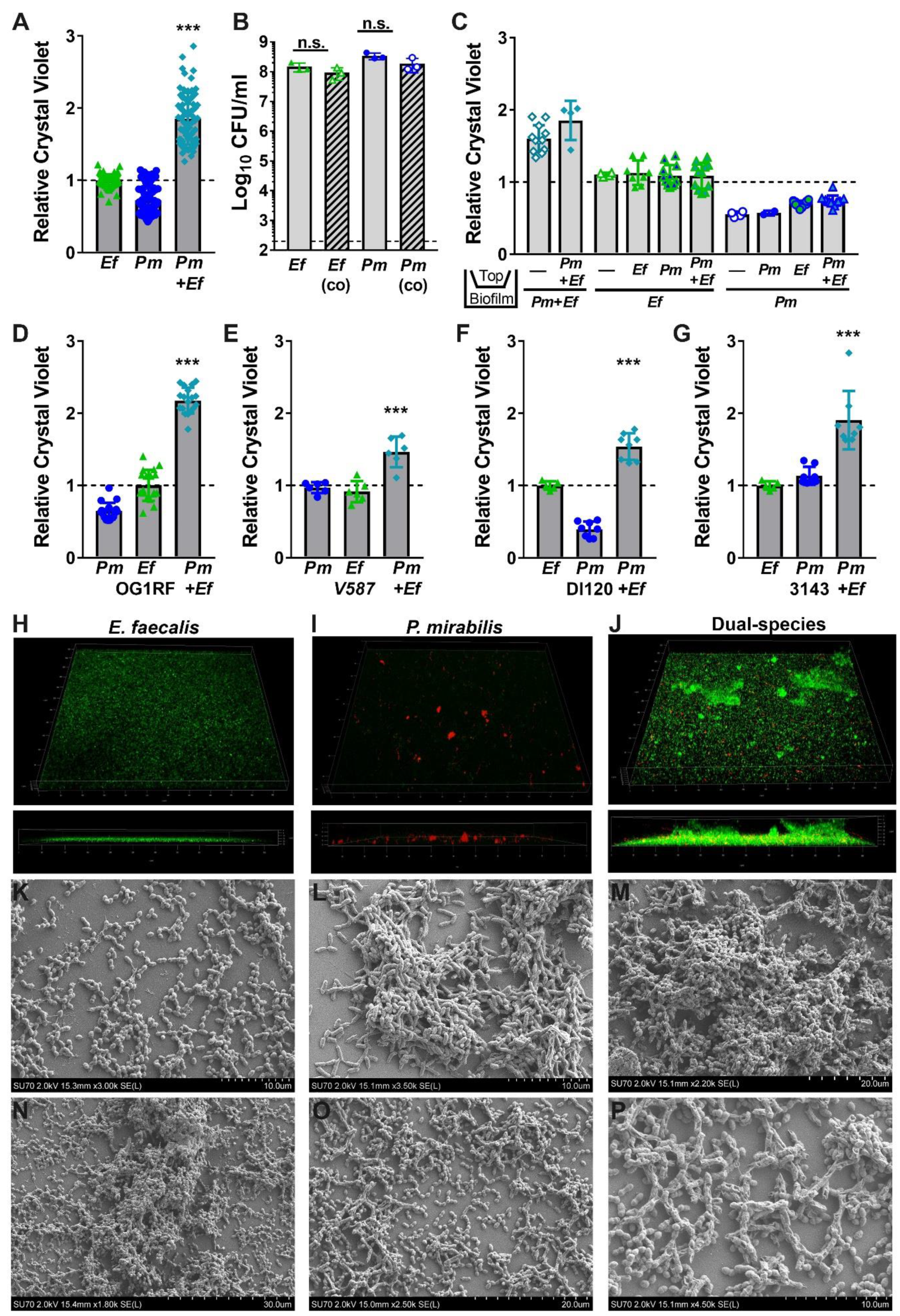
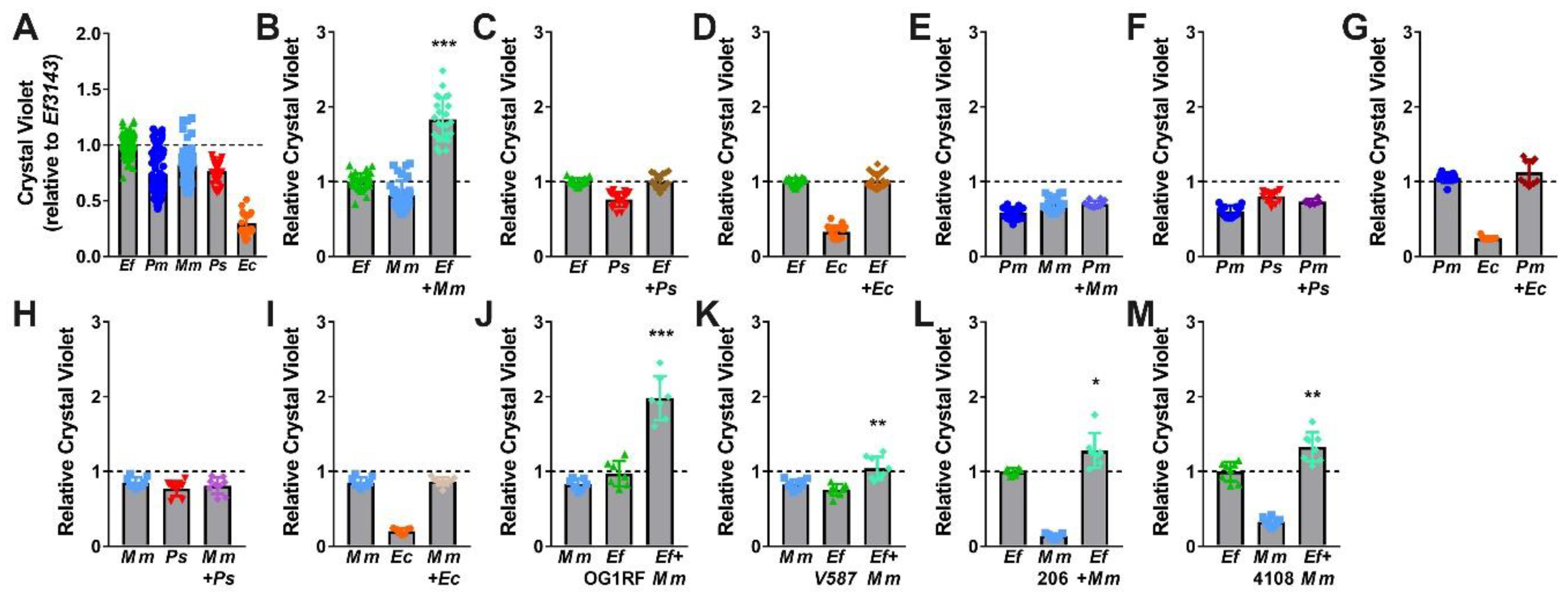
2.3. Co-culture of Enterococcus faecalis with Proteus mirabilis Enhances Biofilm Biomass in a Contact-Dependent Manner
2.4. Enterococcus faecalis and Proteus mirabilis Co-culture Alters Biofilm Architecture
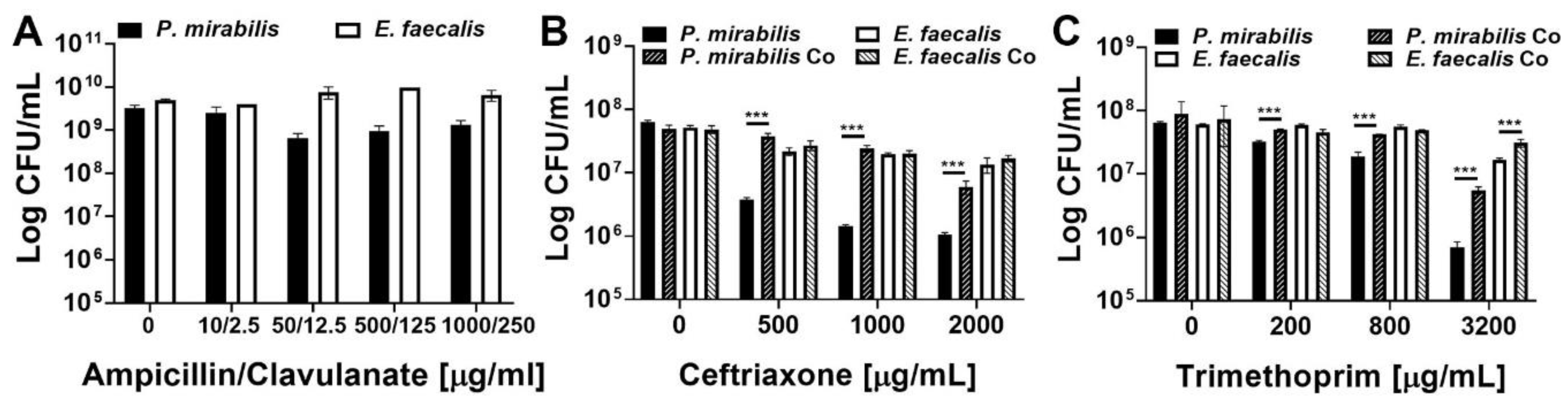
2.5. Enterococcus faecalis and Proteus mirabilis Dual-species Biofilms Exhibit Enhanced Antimicrobial Resistance
2.6. Co-culture of Enterococcus faecalis with Morganella morganii also Enhances Biofilm Biomass
3. Discussion
4. Materials and Methods
4.1. Urine Colonization Study Design and Participants
4.2. Urine Collection for Use in Biofilm Time-Course Microscopy Experiments
4.3. Bacterial Strains
4.4. Bacterial Culture Conditions
4.5. In Vivo Mouse Model
4.6. Antibodies
4.7. Immunofluorescence
4.8. Catheter Imaging
4.9. Biofilm Formation for Determination of Viability
4.10. Crystal Violet Quantification
4.11. Confocal Laser Scanning Microscopy
4.12. Scanning Electron Microscopy
4.13. Time-Course Biofilm Imaging
4.14. Minimum Inhibitory Concentration Assays
4.15. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hooton, T.M.; Bradley, S.F.; Cardenas, D.D.; Colgan, R.; Geerlings, S.E.; Rice, J.C.; Saint, S.; Schaeffer, A.J.; Tambayh, P.A.; Tenke, P.; et al. Diagnosis, Prevention, and Treatment of Catheter-Associated Urinary Tract Infection in Adults: 2009 International Clinical Practice Guidelines from the Infectious Diseases Society of America. Clin. Inf. Dis. 2010, 50, 625–663. [Google Scholar]
- Nicolle, L.E. Urinary Catheter-Associated Infections. Inf. Dis. Clin. N. Am. 2012, 26, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Dudeck, M.A.; Edwards, J.R.; Allen-Bridson, K.; Gross, C.; Malpiedi, P.J.; Peterson, K.D.; Pollock, D.A.; Weiner, L.M.; Sievert, D.M. National Healthcare Safety Network report, data summary for 2013, Device-associated Module. Am. J. Inf. Control 2015, 43, 206–221. [Google Scholar]
- Rogers, M.A.M.; Mody, L.; Kaufman, S.R.; Fries, B.E.; McMahon, L.F.; Saint, S. Use of Urinary Collection Devices in Skilled Nursing Facilities in Five States. J. Am. Geriatr. Soc. 2008, 56, 854–861. [Google Scholar]
- Tsan, L.; Langberg, R.; Davis, C.; Phillips, Y.; Pierce, J.; Hojlo, C.; Gibert, C.; Gaynes, R.; Montgomery, O.; Bradley, S.; et al. Nursing home-associated infections in Department of Veterans Affairs community living centers. Am. J. Inf. Control 2010, 38, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Kunin, C.M.; Douthitt, S.; Dancing, J.; Anderson, J.; Moeschberger, M. The Association between the Use of Urinary Catheters and Morbidity and Mortality among Elderly Patients in Nursing-Homes. Am. J. Epidemiol. 1992, 135, 291–301. [Google Scholar]
- Crnich, C.J.; Drinka, P. Medical device-associated infections in the long-term care setting. Infect. Dis. Clin. N. Am. 2012, 26, 143–164. [Google Scholar]
- Nicolle, L.E. Catheter associated urinary tract infections. Antimicrob. Resist. Infect. Control. 2014, 17, 299–303. [Google Scholar]
- Hofseth, L.J.; Dunn, B.P.; Rosin, M.P. Micronucleus frequencies in urothelial cells of catheterized patients with chronic bladder inflammation. Mutation Res. Fundam. Mol. Mech. Mutagenesis 1996, 352, 65–72. [Google Scholar] [CrossRef]
- Anderson, R.U. Response of bladder and urethral mucosa to catheterization. JAMA 1979, 242, 451–453. [Google Scholar] [CrossRef]
- Armbruster, C.E.; Smith, S.N.; Johnson, A.O.; DeOrnellas, V.; Eaton, K.A.; Yep, A.; Mody, L.; Wu, W.; Mobley, H.L.T. The Pathogenic Potential of Proteus mirabilis is Enhanced by Other Uropathogens During Polymicrobial Urinary Tract Infection. Infect. Immun. 2017, 85, 2. [Google Scholar]
- Nicolle, L.E. Catheter-related urinary tract infection: Practical management in the elderly. Drugs Aging 2014, 31, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nickel, J.C.; Costerton, J.W.; Mclean, R.J.C.; Olson, M. Bacterial Biofilms—Influence on the Pathogenesis, Diagnosis and Treatment of Urinary-Tract Infections. J. Antimicrob. Chemother. 1994, 33, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, B.L.; Aneas, M.A.; Juliano, L.; Palma, M.S.; Lebrun, I.; Portaro, F.C. Development of an operational substrate for ZapA, a metalloprotease secreted by the bacterium Proteus mirabilis. Braz. J. Med. Biol. Res. 2000, 33, 765–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, J.W.; Tenney, J.H.; Hoopes, J.M.; Muncie, H.L.; Anthony, W.C. A Prospective Microbiologic Study of Bacteriuria in Patients with Chronic Indwelling Urethral Catheters. J. Infect. Dis. 1982, 146, 719–723. [Google Scholar] [CrossRef]
- Walker, J.N.; Flores-Mireles, A.L.; Lynch, A.J.L.; Pinkner, C.; Caparon, M.G.; Hultgren, S.J.; Desai, A. High-resolution imaging reveals microbial biofilms on patient urinary catheters despite antibiotic administration. World J. Urol. 2019. [Google Scholar] [CrossRef]
- Nicolle, L.E. Catheter-Related Urinary Tract Infection. Drugs Aging 2005, 22, 627–639. [Google Scholar]
- Armbruster, C.E.; Mobley, H.L.T. Merging mythology and morphology: The multifaceted lifestyle of Proteus mirabilis. Nat. Rev. Microbiol. 2012, 10, 743–754. [Google Scholar]
- Armbruster, C.E.; Prenovost, K.; Mobley, H.L.T.; Mody, L. How Often Do Clinically Diagnosed Catheter-Associated Urinary Tract Infections in Nursing Home Residents Meet Standardized Criteria? J. Am. Geriatr. Soc. 2017, 65, 395–401. [Google Scholar]
- Dedeic-Ljubovic, A.; Hukic, M. Catheter-related urinary tract infection in patients suffering from spinal cord injuries. Bosn. J. Basic Med. Sci. 2009, 9, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Ronald, A. The etiology of urinary tract infection: Traditional and emerging pathogens. Dis. Mon. 2003, 49, 71–82. [Google Scholar] [PubMed]
- Siegman-Igra, Y.; Kulka, T.; Schwartz, D.; Konforti, N. Polymicrobial and monomicrobial bacteraemic urinary tract infection. J. Hosp. Infect. 1994, 28, 49–56. [Google Scholar] [PubMed]
- Kunin, C.M. Blockage of urinary catheters: Role of microorganisms and constituents of the urine on formation of encrustations. J. Clin. Epidemiol. 1989, 42, 835–842. [Google Scholar] [PubMed]
- Rahav, G.; Pinco, E.; Silbaq, F.; Bercovier, H. Molecular epidemiology of catheter-associated bacteriuria in nursing home patients. J. Clin. Microbiol. 1994, 32, 1031–1034. [Google Scholar]
- Mobley, H.L.T.; Warren, J.W. Urease-Positive Bacteriuria and Obstruction of Long-Term Urinary Catheters. J. Clin. Microbiol. 1987, 25, 2216–2217. [Google Scholar]
- Breitenbucher, R.B. Bacterial changes in the urine samples of patients with long-term indwelling catheters. Arch. Intern. Med. 1984, 144, 1585–1588. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Lamont, R.J. Dancing with the Stars: How Choreographed Bacterial Interactions Dictate Nososymbiocity and Give Rise to Keystone Pathogens, Accessory Pathogens, and Pathobionts. Trends Microbiol. 2016, 24, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Tien, B.Y.Q.; Goh, H.M.S.; Chong, K.K.L.; Bhaduri-Tagore, S.; Holec, S.; Dress, R.; Ginhoux, F.; Ingersoll, M.A.; Williams, R.B.H.; Kline, K.A. Enterococcus faecalis Promotes Innate Immune Suppression and Polymicrobial Catheter-Associated Urinary Tract Infection. Infect. Immun. 2017, 85, 12. [Google Scholar]
- Kline, K.A.; Schwartz, D.J.; Gilbert, N.M.; Hultgren, S.J.; Lewis, A.L. Immune modulation by group B Streptococcus influences host susceptibility to urinary tract infection by uropathogenic Escherichia coli. Infect. Immun. 2012, 80, 4186–4194. [Google Scholar]
- Alteri, C.J.; Himpsl, S.D.; Mobley, H.L. Preferential use of central metabolism in vivo reveals a nutritional basis for polymicrobial infection. PLoS Pathog. 2015, 11. [Google Scholar] [CrossRef] [Green Version]
- Armbruster, C.E.; Smith, S.N.; Yep, A.; Mobley, H.L.T. Increased Incidence of Urolithiasis and Bacteremia During Proteus mirabilis and Providencia stuartii Coinfection Due to Synergistic Induction of Urease Activity. J. Infect. Dis. 2014, 209, 1524–1532. [Google Scholar] [PubMed] [Green Version]
- Hjel, E.; Lundell-Etherden, I.; Mardh, P.A. Ascending urinary tract infections in rats induced by Staphylococcus saprophyticus and Proteus mirabilis. Acta Pathol. Microbiol. Immunol. Scand. B 1987, 95, 347–350. [Google Scholar]
- Tsuchimori, N.; Hayashi, R.; Shino, A.; Yamazaki, T.; Okonogi, K. Enterococcus faecalis aggravates pyelonephritis caused by Pseudomonas aeruginosa in experimental ascending mixed urinary tract infection in mice. Infect. Immun. 1994, 62, 4534–4541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Learman, B.S.; Brauer, A.L.; Eaton, K.A.; Armbruster, C.E. A Rare Opportunist, Morganella morganii, Decreases Severity of Polymicrobial Catheter-Associated Urinary Tract Infection. Infect. Immun. 2019, 88. [Google Scholar] [CrossRef]
- Armbruster, C.E.; Brauer, A.L.; Humby, M.S. Prospective assessment of catheter-associated bacteriuria in nursing home residents: Clinical presentation, epidemiology, and dynamics medrxiv. medRxiv 2020. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Bauman, T.M.; Potretzke, A.M.; Schreiber, H.L.; Park, A.M.; Pinkner, J.S.; Caparon, M.G.; Hultgren, S.J.; Desai, A. Fibrinogen Release and Deposition on Urinary Catheters Placed during Urological Procedures. J. Urol. 2016, 196, 416–421. [Google Scholar]
- Flores-Mireles, A.L.; Pinkner, J.S.; Caparon, M.G.; Hultgren, S.J. EbpA vaccine antibodies block binding of Enterococcus faecalis to fibrinogen to prevent catheter-associated bladder infection in mice. Sci. Transl. Med. 2014, 6, 254. [Google Scholar]
- Adler, J.; Parmryd, I. Colocalization Analysis in Fluorescence Microscopy. In Cell Imaging Techniques: Methods and Protocols; Taatjes, D.J., Roth, J., Eds.; Humana Press: Totowa, NJ, USA, 2010; pp. 97–109. [Google Scholar] [CrossRef] [Green Version]
- Stickler, D.; Ganderton, L.; King, J.; Nettleton, J.; Winters, C. Proteus mirabilis biofilms and the encrustation of urethral catheters. Urol. Res. 1993, 21, 407–411. [Google Scholar] [CrossRef]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar]
- Stickler, D.J. Bacterial biofilms in patients with indwelling urinary catheters. Nat. Clin. Pract. Urol. 2008, 5, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Ch’ng, J.-H.; Chong, K.K.L.; Lam, L.N.; Wong, J.J.; Kline, K.A. Biofilm-associated infection by enterococci. Nat. Rev. Microbiol. 2019, 17, 82–94. [Google Scholar] [PubMed]
- Kline, K.A.; Lewis, A.L. Gram-Positive Uropathogens, Polymicrobial Urinary Tract Infection, and the Emerging Microbiota of the Urinary Tract. Urin. Tract Infect. Mol. Pathog. Clin. Manag. 2017. [Google Scholar] [CrossRef] [Green Version]
- Hooton, T.M. Clinical practice. Uncomplicated urinary tract infection. N. Engl. J. Med. 2012, 366, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, C.E.; Mobley, H.L. Proteus species. antimicrobe. E-Sun Technologies, Microbes. Yu, V.L., Ed.; 2015. Available online: http://www.antimicrobe.org/b226.asp. (accessed on 9 September 2020).
- Korytny, A.; Riesenberg, K.; Saidel-Odes, L.; Schlaeffer, F.; Borer, A. Bloodstream infections caused by multi-drug resistant Proteus mirabilis: Epidemiology, risk factors and impact of multi-drug resistance. Infect. Dis. 2016, 48, 428–431. [Google Scholar]
- Kanayama, A.; Kobayashi, I.; Shibuya, K. Distribution and antimicrobial susceptibility profile of extended-spectrum β-lactamase-producing Proteus mirabilis strains recently isolated in Japan. Int. J. Antimicrob. Agents 2015, 45, 113–118. [Google Scholar]
- Wang, J.-T.; Chen, P.-C.; Chang, S.-C.; Shiau, Y.-R.; Wang, H.-Y.; Lai, J.-F.; Huang, I.-W.; Tan, M.-C.; Lauderdale, T.-L.Y. Antimicrobial susceptibilities of Proteus mirabilis: A longitudinal nationwide study from the Taiwan surveillance of antimicrobial resistance (TSAR) program. BMC Infect. Dis. 2014, 14, 486. [Google Scholar] [CrossRef] [Green Version]
- Luzzaro, F.; Ortisi, G.; Larosa, M.; Drago, M.; Brigante, G.; Gesu, G. Prevalence and epidemiology of microbial pathogens causing bloodstream infections: Results of the OASIS multicenter study. Diagn. Microbiol. Infect. Dis. 2011, 69, 363–369. [Google Scholar] [CrossRef]
- Layton, B.A.; Walters, S.P.; Lam, L.H.; Boehm, A.B. Enterococcus species distribution among human and animal hosts using multiplex PCR. J. Appl. Microbiol. 2010, 109, 539–547. [Google Scholar]
- Mobley, H.L.; Green, D.M.; Trifillis, A.L.; Johnson, D.E.; Chippendale, G.R.; Lockatell, C.V.; Jones, B.D.; Warren, J.W. Pyelonephritogenic Escherichia coli and killing of cultured human renal proximal tubular epithelial cells: Role of hemolysin in some strains. Infect. Immun. 1990, 58, 1281–1289. [Google Scholar]
- Sahm, D.F.; Kissinger, J.; Gilmore, M.S.; Murray, P.R.; Mulder, R.; Solliday, J.; Clarke, B. In vitro susceptibility studies of vancomycin-resistant Enterococcus faecalis. Antimicrob Agents Chemother 1989, 33, 1588–1591. [Google Scholar]
- Oliver, D.R.; Brown, B.L.; Clewell, D.B. Analysis of plasmid deoxyribonucleic acid in a cariogenic strain of Streptococcus faecalis: An approach to identifying genetic determinants on cryptic plasmids. J. Bacteriol. 1977, 130, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Gold, O.G.; Jordan, H.V.; van Houte, J. The prevalence of enterococci in the human mouth and their pathogenicity in animal models. Arch. Oral Biol. 1975, 20, 473–477. [Google Scholar]
- Dunny, G.M.; Brown, B.L.; Clewell, D.B. Induced cell aggregation and mating in Streptococcus faecalis: Evidence for a bacterial sex pheromone. Proc. Natl. Acad. Sci. USA 1978, 75, 3479–3483. [Google Scholar] [CrossRef] [Green Version]
- Bevis, B.J.; Glick, B.S. Rapidly maturing variants of the Discosoma red fluorescent protein (DsRed). Nat. Biotechnol. 2002, 20, 83–87. [Google Scholar] [CrossRef]
- Lizier, M.; Sarra, P.G.; Cauda, R.; Lucchini, F. Comparison of expression vectors in Lactobacillus reuteri strains. FEMS Microbiol. Lett. 2010, 308, 8–15. [Google Scholar]
- Monk, I.R.; Shah, I.M.; Xu, M.; Tan, M.W.; Foster, T.J. Transforming the untransformable: Application of direct transformation to manipulate genetically Staphylococcus aureus and Staphylococcus epidermidis. mBio 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Schaffer, J.N.; Norsworthy, A.N.; Sun, T.-T.; Pearson, M.M. Proteus mirabilis fimbriae- and urease-dependent clusters assemble in an extracellular niche to initiate bladder stone formation. Proc. Natl. Acad. Sci. USA 2016, 113, 4494–4499. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar]
- Au-Mah, T.-F. Establishing the Minimal Bactericidal Concentration of an Antimicrobial Agent for Planktonic Cells (MBC-P) and Biofilm Cells (MBC-B). JoVE 2014. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. mirabilis HI4320 | E. faecalis 3143 | |
|---|---|---|
| Ampicillin | 16 | 32 |
| Ceftriaxone | 4 | >64 |
| Daptomycin | >256 | >256 |
| Trimethoprim | 32 | >64 |
| Nitrofurantoin | 128 | 128 |
| Name | Sequence |
|---|---|
| AOJ_424 | ACACTAGGCCCGGTCTCCCAAGAATTAGAAATGAGTAGAT |
| AOJ_425 | ATCGATACCGAGACCTTCTATTTAATCACTTTGACTAGCA |
| AOJ_426 | CCTAGTGTTTTAGGAGACCGAGCCACTATCGACTACGC |
| AOJ_427 | TGATTAAATAGAAGGTCTCGGTATCGATAAGCTTAGTCTA |
| AOJ_428 | CGATAGTGGCTCGGTCTCCTAAAACACTAGGCCCAAGA |
| AOJ_429 | CTGCGGTCTCATGGTCCATGCGAGAGTAGGGAACT |
| AOJ_430 | CGCATGGACCATGAGACCGCAGGTTAGTGACATTAGAA |
| AOJ_431 | CTCATTATTTGGGCAGGAGACCCTTTAGTGAGGGTTAATT |
| AOJ_432 | CTAAAGGGTCTCCTGCCCAAATAATGAGCTAGCCCG |
| AOJ_433 | TCTAATTCTTGGGAGACCGGGCCTAGTGTTTTAGATCC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaston, J.R.; Andersen, M.J.; Johnson, A.O.; Bair, K.L.; Sullivan, C.M.; Guterman, L.B.; White, A.N.; Brauer, A.L.; Learman, B.S.; Flores-Mireles, A.L.; et al. Enterococcus faecalis Polymicrobial Interactions Facilitate Biofilm Formation, Antibiotic Recalcitrance, and Persistent Colonization of the Catheterized Urinary Tract. Pathogens 2020, 9, 835. https://doi.org/10.3390/pathogens9100835
Gaston JR, Andersen MJ, Johnson AO, Bair KL, Sullivan CM, Guterman LB, White AN, Brauer AL, Learman BS, Flores-Mireles AL, et al. Enterococcus faecalis Polymicrobial Interactions Facilitate Biofilm Formation, Antibiotic Recalcitrance, and Persistent Colonization of the Catheterized Urinary Tract. Pathogens. 2020; 9(10):835. https://doi.org/10.3390/pathogens9100835
Chicago/Turabian StyleGaston, Jordan R., Marissa J. Andersen, Alexandra O. Johnson, Kirsten L. Bair, Christopher M. Sullivan, L. Beryl Guterman, Ashely N. White, Aimee L. Brauer, Brian S. Learman, Ana L. Flores-Mireles, and et al. 2020. "Enterococcus faecalis Polymicrobial Interactions Facilitate Biofilm Formation, Antibiotic Recalcitrance, and Persistent Colonization of the Catheterized Urinary Tract" Pathogens 9, no. 10: 835. https://doi.org/10.3390/pathogens9100835