Transstadial Transmission and Replication Kinetics of West Nile Virus Lineage 1 in Laboratory Reared Ixodes ricinus Ticks

 , and
, and
Abstract
:1. Introduction
2. Results
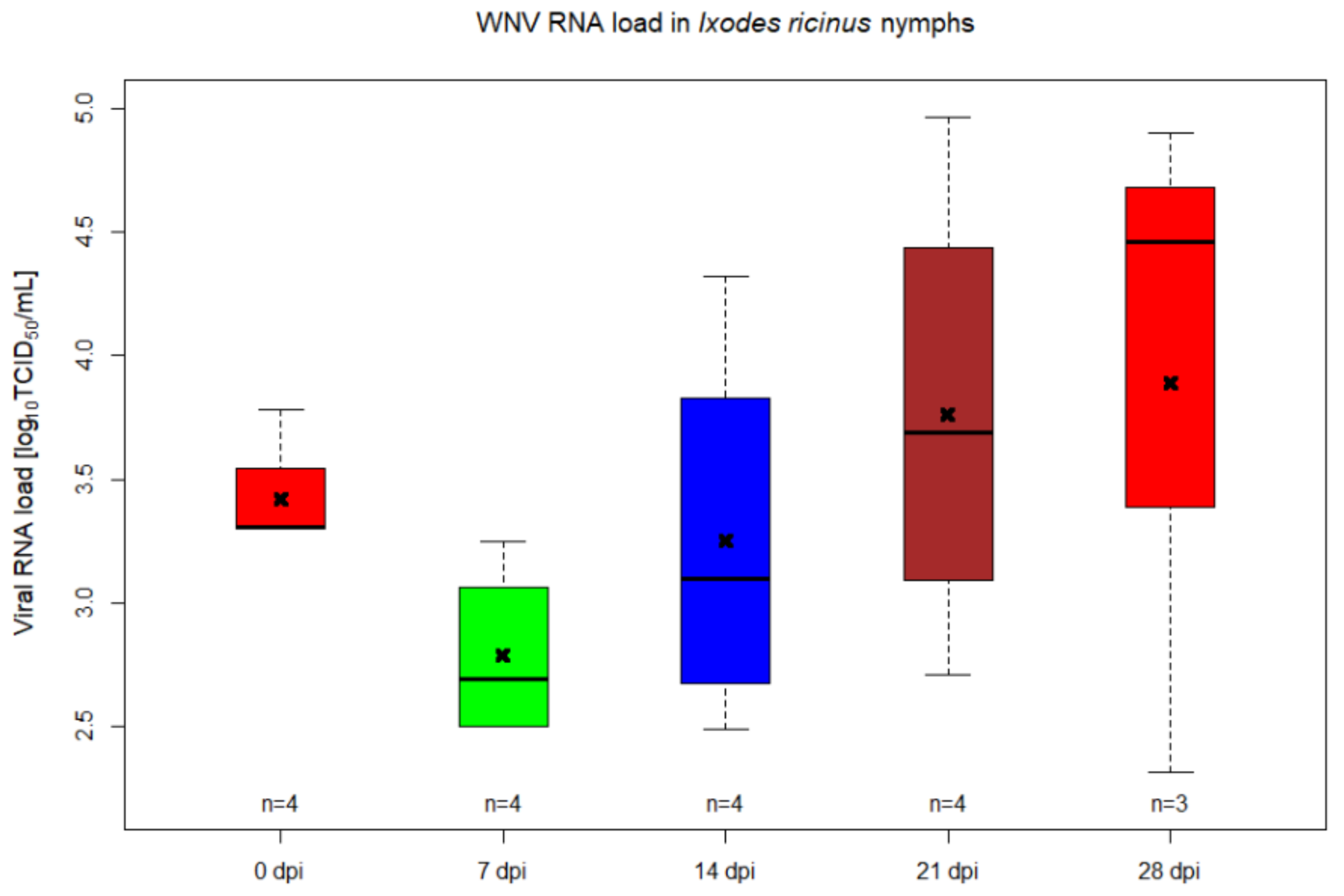
2.1. WNV RNA Kinetics in Ixodes ricinus Injected Nymphs
2.2. Transstadial Transmission of WNV
2.3. Vector Competence of Ixodes ricinus Females
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Tick Colony Rearing
4.3. Virus
4.4. Infection of Ticks by Percoxal Injection
4.5. In Vitro Feeding System
4.6. Artificial Tick Feeding
4.7. Ticks and Blood Samples Processing
4.8. Molecular Detection of WNV RNA
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. Biomed. Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, G.L.; Marfin, A.A.; Lanciotti, R.S.; Gubler, D.J. West Nile virus. Lancet Infect. Dis. 2002, 2, 519–529. [Google Scholar] [CrossRef]
- Kolodziejek, J.; Marinov, M.; Kiss, B.J.; Alexe, V.; Nowotny, N. The complete sequence of a West Nile virus lineage 2 strain detected in a Hyalomma marginatum marginatum tick collected from a song thrush (Turdus philomelos) in eastern Romania in 2013 revealed closest genetic relationship to strain Volgograd 2007. PLoS ONE 2014, 9, e109905. [Google Scholar] [CrossRef] [Green Version]
- Hoogstraal, H.; Clifford, C.M.; Keirans, J.E.; Kaiser, M.N.; Evans, D.E. The Ornithodoros (Alectorobius) capensis group (Acarina: Ixodoidea: Argasidae) of the palearctic and oriental regions. O. (A.) maritimus: Identity, marine bird hosts, virus infections, and distribution in western Europe and northwestern Africa. J. Parasitol. 1976, 62, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Lvov, D.K.; Timopheeva, A.A.; Smirnov, V.A.; Gromashevsky, V.L.; Sidorova, G.A.; Nikiforov, L.P.; Sazonov, A.A.; Andreev, A.P.; Skvortzova, T.M.; Beresina, L.K.; et al. Ecology of tick-borne viruses in colonies of birds in the USSR. Med.Biol. 1975, 53, 325–330. [Google Scholar]
- Lwande, O.W.; Lutomiah, J.; Obanda, V.; Gakuya, F.; Mutisya, J.; Mulwa, F.; Michuki, G.; Chepkorir, E.; Fischer, A.; Venter, M.; et al. Isolation of tick and mosquito-borne arboviruses from ticks sampled from livestock and wild animal hosts in Ijara District, Kenya. Vector Borne Zoonotic Dis. 2013, 13, 637–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platonov, A.E. West Nile encephalitis in Russia 1999-2001: Were we ready? Are we ready? Ann. N. Y. Acad. Sci. 2001, 951, 102–116. [Google Scholar] [CrossRef]
- Flores, F.S.; Zanluca, C.; Guglielmone, A.A.; Duarte Dos Santos, C.N.; Labruna, M.B.; Diaz, A. Vector competence for West Nile virus and St. Louis Encephalitis virus (Flavivirus) of three tick species of the genus Amblyomma (Acari: Ixodidae). Am. J. Trop. Med. Hyg. 2019, 100, 1230–1235. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.F.; Main, A.J.; Andreadis, T.G.; Wikel, S.K.; Vossbrinck, C.R. Transstadial transfer of West Nile virus by three species of ixodid ticks (Acari: Ixodidae). J. Med. Entomol. 2003, 40, 528–533. [Google Scholar] [CrossRef]
- Reisen, W.K.; Brault, A.C.; Martinez, V.M.; Fang, Y.; Simmons, K.; Garcia, S.; Omi-Olsen, E.; Lane, R.S. Ability of transstadially infected Ixodes pacificus (Acari: Ixodidae) to transmit West Nile virus to song sparrows or western fence lizards. J. Med. Entomol. 2007, 44, 320–327. [Google Scholar] [CrossRef]
- Formosinho, P.; Santos-Silva, M.M. Experimental infection of Hyalomma marginatum ticks with West Nile virus. Acta Virol. 2006, 50, 175–180. [Google Scholar] [PubMed]
- Lawrie, C.H.; Uzcátegui, N.Y.; Gould, E.A.; Nuttall, P.A. Ixodid and argasid tick species and West Nile virus. Emerg. Infect. Dis. 2004, 10, 653–657. [Google Scholar] [CrossRef]
- Whitman, L.; Aitken, T.H. Potentiality of Ornithodoros moubata Murray (Acarina, Argasidae) as a reservoir vector of West Nile virus. Ann. Trop. Med. Parasitol. 1960, 54, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Špitalská, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef]
- Randolph, S.E. Tick-borne disease systems emerge from the shadows: The beauty lies in molecular detail, the message in epidemiology. Parasitology 2009, 136, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada- Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Hai, V.V.; Almeras, L.; Socolovschi, C.; Raoult, D.; Parola, P.; Pagès, F. Monitoring human tick-borne disease risk and tick bite exposure in Europe: Available tools and promising future methods. Ticks Tick Borne Dis. 2014, 5, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Gritsun, T.S.; Lashkevich, V.A.; Gould, E.A. Tick-borne encephalitis. Antivir. Res. 2003, 57, 129–146. [Google Scholar] [CrossRef]
- Rizzoli, A.; Hauffe, H.; Carpi, G.; Vourc’h, G.I.; Neteler, M.; Rosà, R. Lyme borreliosis in Europe. EuroSurveillance 2011, 16, 19906. [Google Scholar]
- Rockstroh, A.; Moges, B.; Berneck, B.S.; Sattler, T.; Revilla- Fernández, S.; Schmoll, F.; Pacenti, M.; Sinigaglia, A.; Barzon, L.; Schmidt-Chanasit, J.; et al. Specific detection and differentiation of tick-borne encephalitis and West Nile virus induced IgG antibodies in humans and horses. Transbound. Emerg. Dis. 2019, 66, 1701–1708. [Google Scholar] [CrossRef]
- Norte, A.C.; de Carvalho, I.L.; Ramos, J.A.; Gonçalves, M.; Gern, L.; Núncio, M.S. Diversity and seasonal patterns of ticks parasitizing wild birds in western Portugal. Exp. Appl. Acarol. 2012, 58, 327–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camp, J.V.; Nowotny, N. The knowns and unknowns of West Nile virus in Europe: What did we learn from the 2018 outbreak? Expert Rev. Anti Infect. Ther. 2020, 18, 145–154. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control (ECDC). Epidemiological Update: West Nile Virus Transmission Season in Europe. 2019. Available online: https://www.ecdc.europa.eu/en/news-events/epidemiological-update-west-nile-virus-transmission-season-europe-2019 (accessed on 18 September 2020).
- Ziegler, U.; Lühken, R.; Keller, M.; Cadar, D.; van der Grinten, E.; Michel, F.; Albrecht, K.; Eiden, M.; Rinder, M.; Lachmann, L.; et al. West Nile virus epizootic in Germany, 2018. Antiviral. Res. 2019, 162, 39–43. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Tick Maps [Internet]. Available online: https://ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/tick-maps (accessed on 19 August 2020).
- Rechav, Y.; Zyzak, M.; Fielden, L.J.; Childs, J.E. Comparison of methods for introducing and producing artificial infection of ixodid ticks (Acari: Ixodidae) with Ehrlichia chaffeensis. J. Med. Entomol. 1999, 36, 414–419. [Google Scholar] [CrossRef]
- Ličková, M.; Fumacova Havlíková, S.; Sláviková, M.; Slovák, M.; Drexler, J.F.; Klempa, B. Dermacentor reticulatus is a vector of tick-borne encephalitis virus. Ticks Tick Borne Dis. 2020, 11, 101414. [Google Scholar] [CrossRef]
- Vimonish, R.; Johnson, W.C.; Mousel, M.R.; Brayton, K.A.; Scoles, G.A.; Noh, S.M.; Ueti, M.W. Quantitative analysis of Anaplasma marginale acquisition and transmission by Dermacentor andersoni fed in vitro. Sci. Rep. 2020, 10, 470. [Google Scholar] [CrossRef] [Green Version]
- Körner, S.; Makert, G.R.; Mertens-Scholz, K.; Henning, K.; Pfeffer, M.; Starke, A.; Nijhof, A.M.; Ulbert, S. Uptake and fecal excretion of Coxiella burnetii by Ixodes ricinus and Dermacentor marginatus ticks. Parasites Vectors 2020, 13, 75. [Google Scholar] [CrossRef] [Green Version]
- Tajeri, S.; Razmi, G.; Haghparast, A. Establishment of an artificial tick feeding system to study Theileria lestoquardi infection. PLoS ONE 2016, 11, e0169053. [Google Scholar] [CrossRef] [Green Version]
- Bouwknegt, C.; van Rijn, P.A.; Schipper, J.J.; Hölzel, D.; Boonstra, J.; Nijhof, A.M.; van Rooij, E.M.; Jongejan, F. Potential role of ticks as vectors of bluetongue virus. Exp. Appl. Acarol. 2010, 52, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valcárcel, F.; González, J.; González, M.G.; Sánchez, M.; Tercero, J.M.; Elhachimi, L.; Carbonell, J.D.; Olmeda, A.S. Comparative Ecology of Hyalomma lusitanicum and Hyalomma marginatum Koch, 1844 (Acarina: Ixodidae). Insects 2020, 11, 303. [Google Scholar] [CrossRef] [PubMed]
- Kotti, B.K.; Shaposhnikova, L.I.; Evchenko Iu, M.; Levchenko, B.I.; Surkhaev, D.B.; Korzhov, P.N.; Tokhov Iu, M. [Hyalomma marginatum Koch in Stavropol’ region]. Zhurnal Mikrobiol. Epidemiol. Immunobiol. 2001, 6, 105–108. [Google Scholar]
- Nuttall, P.A. Tick-Borne Viruses; Oxford University Press: New York, NY, USA, 2014; Volume 2. [Google Scholar]
- Rückert, C.; Bell-Sakyi, L.; Fazakerley, J.K.; Fragkoudis, R. Antiviral responses of arthropod vectors: An update on recent advances. Virus Dis. 2014, 25, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajdušek, O.; Šíma, R.; Ayllón, N.; Jalovecká, M.; Perner, J.; de la Fuente, J.; Kopáček, P. Interaction of the tick immune system with transmitted pathogens. Front. Cell. Infect. Microbiol. 2013, 3, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garmendia, A.E.; Van Kruiningen, H.J.; French, R.A.; Anderson, J.F.; Andreadis, T.G.; Kumar, A.; West, A.B. Recovery and identification of West Nile virus from a hawk in winter. J. Clin. Microbiol. 2000, 38, 3110–3111. [Google Scholar] [CrossRef] [Green Version]
- Odelola, H.A.; Oduye, O.O. West Nile virus infection of adult mice by oral route. Arch. Virol. 1977, 54, 251–253. [Google Scholar] [CrossRef]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef]
- Chernesky, M.A.; McLean, D.M. Localization of Powassan virus in Dermacentor andersoni ticks by immunofluorescence. Can. J. Microbiol. 1969, 15, 1399–1408. [Google Scholar] [CrossRef]
- Řeháček, J. Development of animal viruses and rickettsiae in ticks and mites. Annu. Rev. Entomol. 1965, 10, 1–24. [Google Scholar] [CrossRef]
- Ramakrishnan, M.A. Determination of 50% endpoint titer using a simple formula. World J. Virol. 2016, 5, 85–86. [Google Scholar] [CrossRef]
- Kröber, T.; Guerin, P.M. In vitro feeding assays for hard ticks. Trends Parasitol. 2007, 23, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Krull, C.; Böhme, B.; Clausen, P.H.; Nijhof, A.M. Optimization of an artificial tick feeding assay for Dermacentor reticulatus. Parasites Vectors 2017, 10, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiden, M.; Vina-Rodriguez, A.; Hoffmann, B.; Ziegler, U.; Groschup, M.H. Two new real-time quantitative reverse transcription polymerase chain reaction assays with unique target sites for the specific and sensitive detection of lineages 1 and 2 West Nile virus strains. J. Vet. Diagn. Investig. 2010, 22, 748–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; R Studio, Inc.: Boston, MA, USA, 2019. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FUs | Feeding Status | Engorged Ticks | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D1 | D2 | D3 | D4 | D5 | D6 | D7 | D8 | D9 | D10 | D11 | D12 | D13 | D14 | D15 | ||
| FU1-1 | N | N | N | N | F | F | F | F | F | N | X | X | X | X | X | 0 |
| FU1-2 | N | N | N | N | N | N | N | N | N | X | X | X | X | X | X | 0 |
| FU1-3 | N | N | N | F | F | F | F | F | F | F | F | F | H | 1 a | ||
| FU1-4 | F | F | F | F | F | F | F | F | N | N | N | N | H | 1 a | ||
| FU2-1 | F | F | F | F | F | F | F | F | N | N | N | N | H | 1 b | ||
| FU2-2 | N | X | X | X | X | X | X | X | X | X | X | X | X | X | X | 0 |
| FU2-3 | N | N | F | F | F | F | N | F | F | F | F | F | F | N | H | 1 a |
| FU2-4 | N | N | N | F | F | F | F | F | F | F | X | X | X | X | X | 0 |
| FU2-5 | N | F | F | F | F | F | F | F | F | H | 1 b | |||||
| FU2-6 | F | N | X | X | X | X | X | X | X | X | X | X | X | X | X | 0 |
| FU3-1 | F | F | F | F | F | F | F | F | F | F | H | 1 a | ||||
| FU3-2 | N | N | N | N | N | F | F | F | F | F | N | N | H | 1 b | ||
| FU3-3 | N | N | N | N | N | N | F | N | N | X | X | X | X | X | X | 0 |
| FU3-4 | F | F | F | F | X | X | X | X | X | X | X | X | X | X | X | 0 |
| FU3-5 | N | N | F | F | F | X | X | X | X | X | X | X | X | X | X | 0 |
| FU3-6 | F | F | F | X | X | X | X | X | X | X | X | X | X | X | X | 0 |
| FU3-7 | N | N | N | F | F | F | F | F | F | X | X | X | X | X | X | 0 |
| FU4-1 | F | F | F | X | X | X | X | X | X | X | X | X | X | X | X | 0 |
| FU4-2 | F | F | F | F | F | F | F | N | N | N | H | 1 a | ||||
| FU4-3 | N | N | N | N | N | N | N | N | N | X | X | X | X | X | X | 0 |
| FU4-4 | N | N | N | F | F | N | X | X | X | X | X | X | X | X | X | 0 |
| FU4-5 | N | N | N | N | X | X | X | X | X | X | X | X | X | X | X | 0 |
| FU4-6 | N | N | F | F | F | F | F | F | F | F | H | 1 | ||||
| Total | 8 | 8 | 11 | 13 | 13 | 12 | 12 | 10 | 9 | 6 | 2 | 2 | 1 | 0 | 0 | 9 |
| Neg. control * | 3 | 3 | 3 | 4 | 4 | 5 | 4 | 5 | 5 | 5 | 3 | 3 | 1 | 1 | H | 3 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Răileanu, C.; Tauchmann, O.; Vasić, A.; Neumann, U.; Tews, B.A.; Silaghi, C. Transstadial Transmission and Replication Kinetics of West Nile Virus Lineage 1 in Laboratory Reared Ixodes ricinus Ticks. Pathogens 2020, 9, 780. https://doi.org/10.3390/pathogens9100780
Răileanu C, Tauchmann O, Vasić A, Neumann U, Tews BA, Silaghi C. Transstadial Transmission and Replication Kinetics of West Nile Virus Lineage 1 in Laboratory Reared Ixodes ricinus Ticks. Pathogens. 2020; 9(10):780. https://doi.org/10.3390/pathogens9100780
Chicago/Turabian StyleRăileanu, Cristian, Oliver Tauchmann, Ana Vasić, Ulrike Neumann, Birke Andrea Tews, and Cornelia Silaghi. 2020. "Transstadial Transmission and Replication Kinetics of West Nile Virus Lineage 1 in Laboratory Reared Ixodes ricinus Ticks" Pathogens 9, no. 10: 780. https://doi.org/10.3390/pathogens9100780