Essential Gene Clusters Involved in Copper Tolerance Identified in Acinetobacter baumannii Clinical and Environmental Isolates

 , ,
, ,
Abstract
:1. Introduction
2. Materials and methods
2.1. Bacterial Strains and Growth Condition
2.2. Antibiotic Susceptibility Testing and PCR Detection of Cabarpenemase Encoding Genes
2.3. Minimum Inhibitory Concentration (MIC) of Copper
2.4. Detections Of Copper-Related Genes in Collections of A. baumannii
2.5. Exposure of A. baumannii Cells to Heavy Metals and Imipenem, and Transmission Electron Microscopy (TEM) Analysis
2.6. Total RNA Isolation, cDNA Synthesis, and Quantitative Real-Time PCR (qPCR)
2.7. Whole Genome Sequencing and Genome Assembly
2.8. Bioinformatic Analysis
2.9. Construction of Chromosomal Circular Map and Genomic Island Comparisons
2.10. Data, Statistical, and Cluster Analysis
3. Results
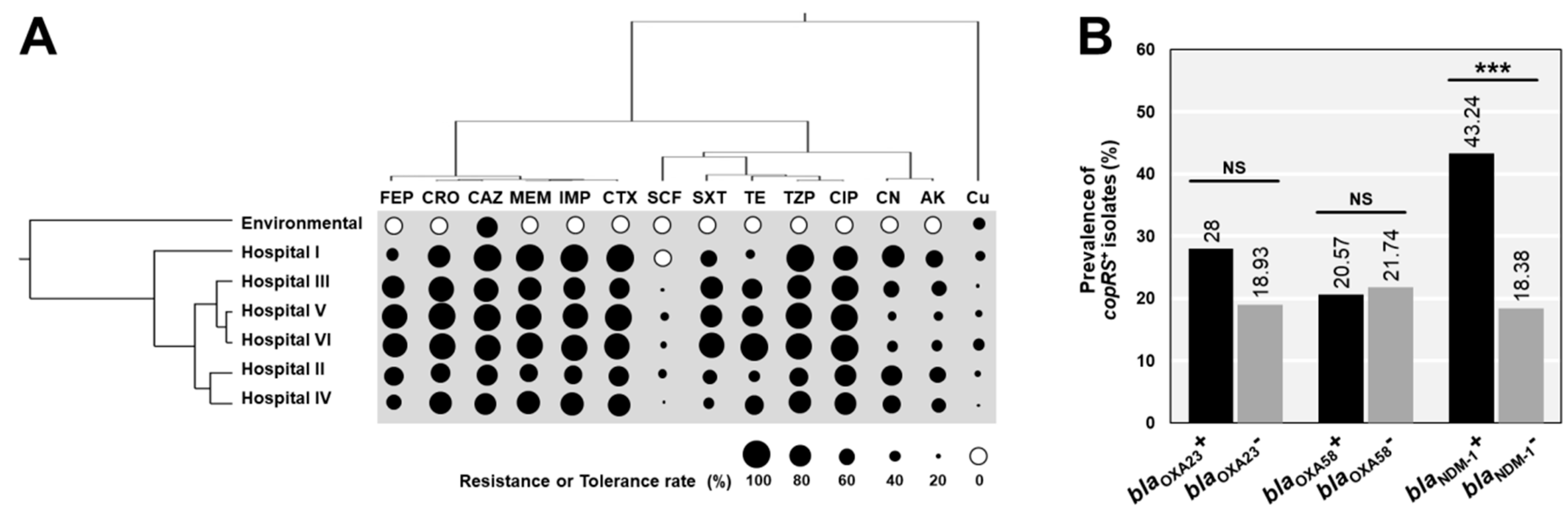
3.1. Prevalence of Copper-Related Genes in A. baumannii
3.2. Copper Susceptibility Profiles of A. baumannii and Their Association with Copper-Related Genes
3.3. Association between Copper Tolerance and Antibiotic Resistances in A. baumannii
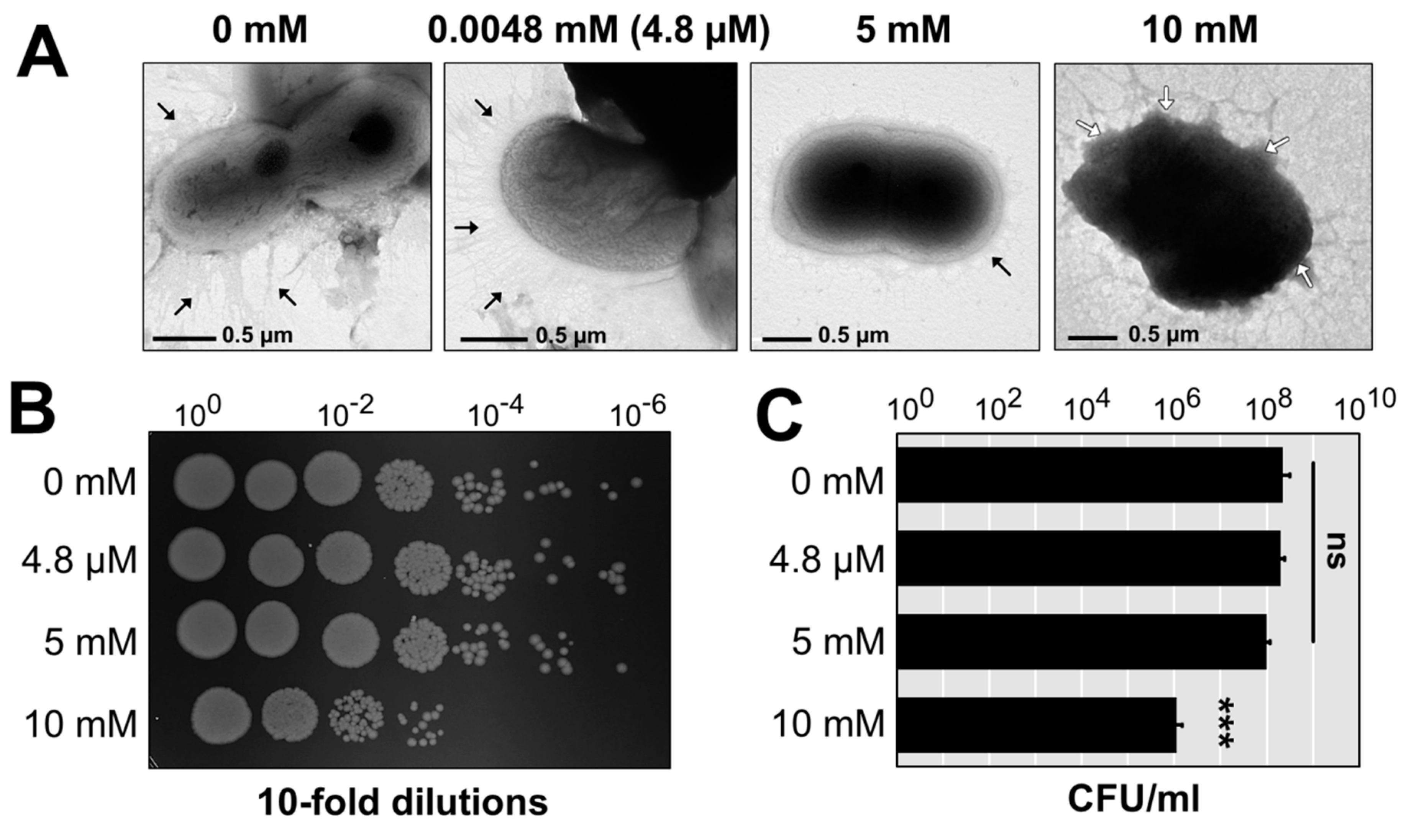
3.4. Visualization of A. baumannii AB003 Challenged with Copper
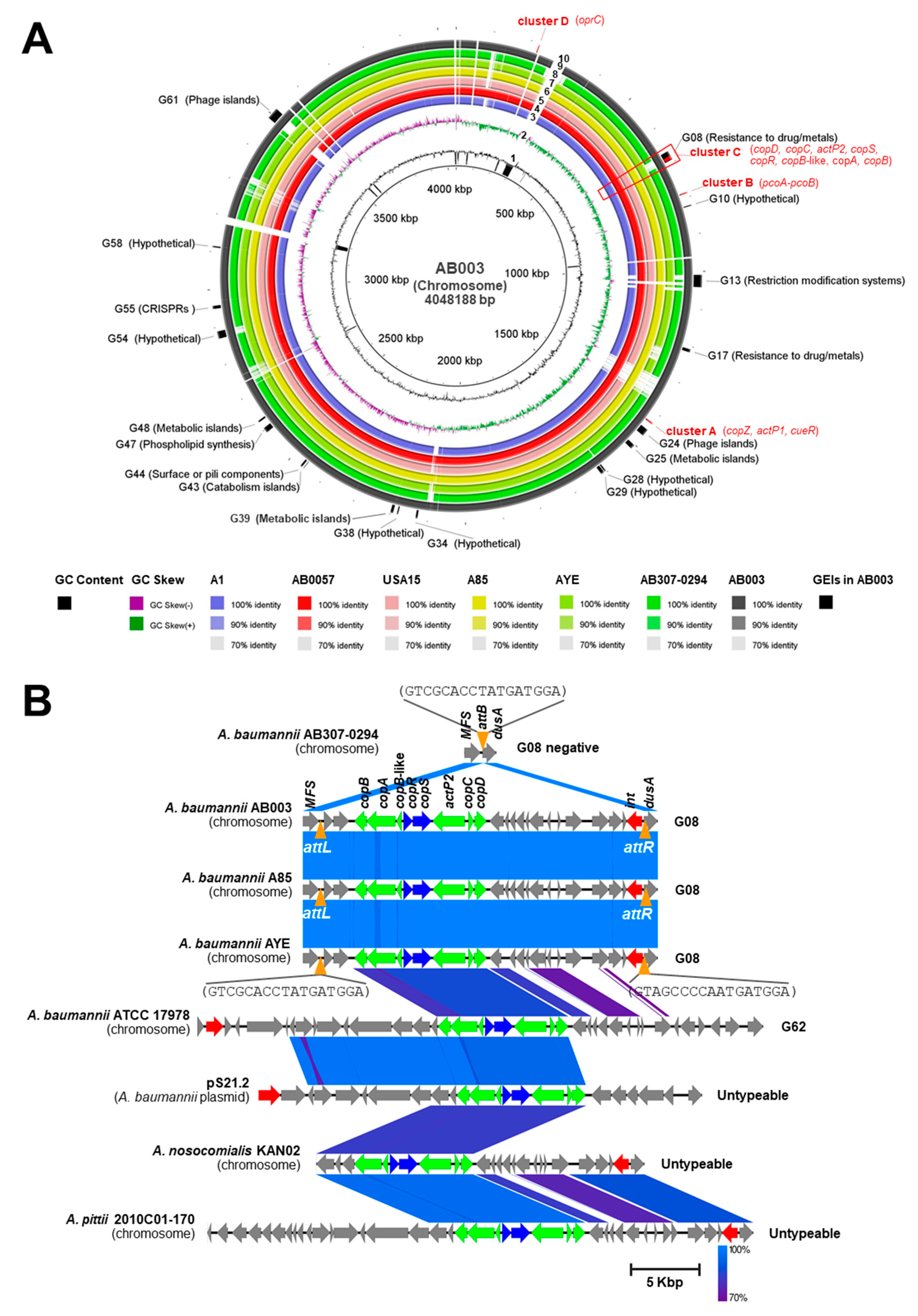
3.5. Whole Genome Sequencing (WGS) and Computational Analysis of AB003
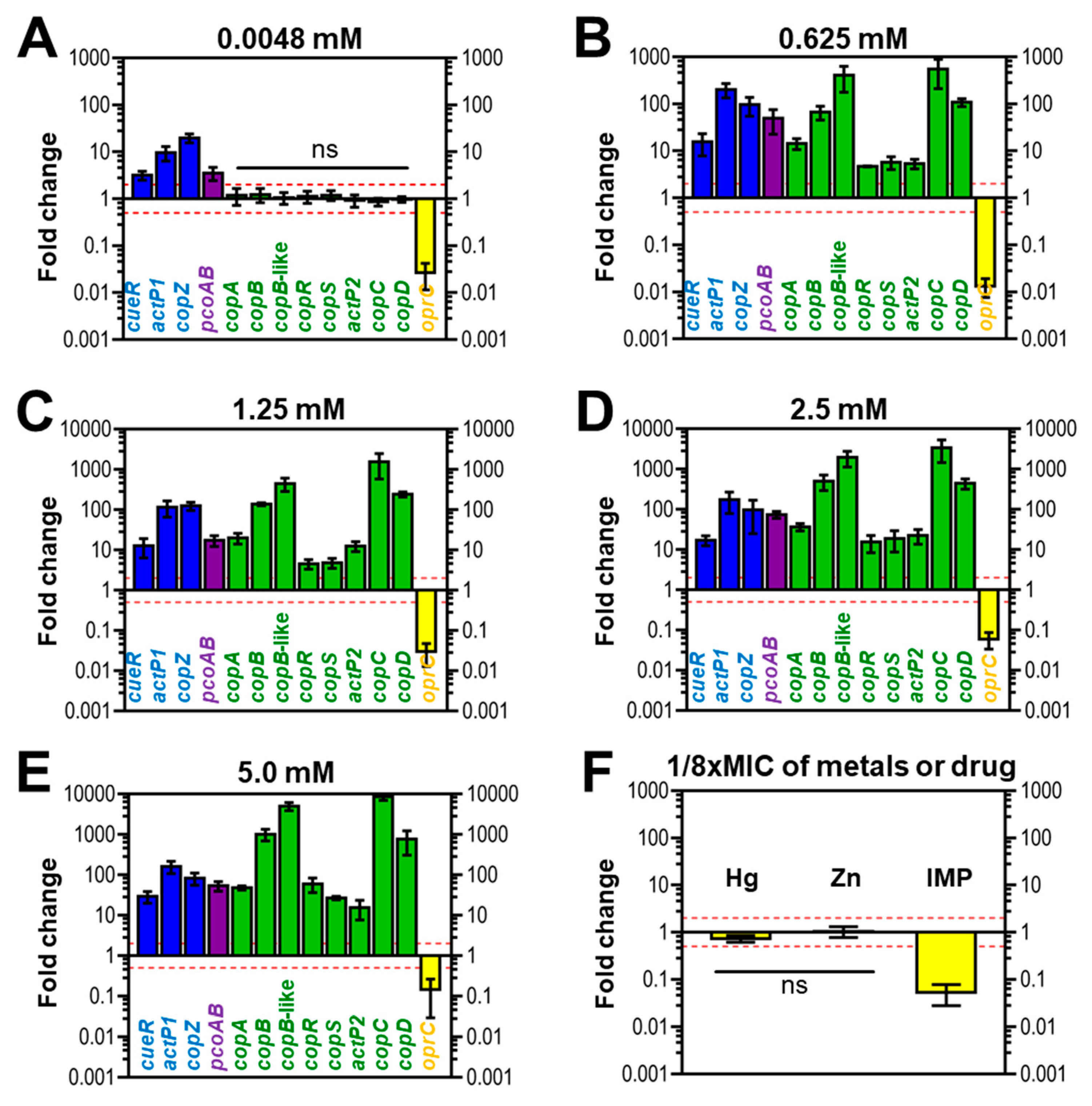
3.6. Expression of Copper-Related Genes in Response to Copper of AB003
4. Discussions and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Borkow, G.; Gabbay, J. Copper, an ancient remedy returning to fight microbial, fungal and viral infections. Curr. Chem. Biol. 2009, 3, 272–278. [Google Scholar]
- Rensing, C.; Grass, G. Escherichia coli mechanisms of copper homeostasis in a changing environment. FEMS Microbiol. Rev. 2003, 27, 197–213. [Google Scholar] [CrossRef] [Green Version]
- Bondarczuk, K.; Piotrowska-Seget, Z. Molecular basis of active copper resistance mechanisms in Gram-negative bacteria. Cell Biol. Toxicol. 2013, 29, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Outten, F.W.; Huffman, D.L.; Hale, J.A.; O’Halloran, T.V. The independent cue and cus systems confer copper tolerance during aerobic and anaerobic growth in Escherichia coli. J. Biol. Chem. 2001, 276, 30670–30677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladomersky, E.; Petris, M.J. Copper tolerance and virulence in bacteria. Metallomics 2015, 7, 957–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaturvedi, K.S.; Hung, C.S.; Crowley, J.R.; Stapleton, A.E.; Henderson, J.P. The siderophore yersiniabactin binds copper to protect pathogens during infection. Nat. Chem. Biol. 2012, 8, 731–736. [Google Scholar] [CrossRef] [Green Version]
- Subashchandrabose, S.; Hazen, T.H.; Brumbaugh, A.R.; Himpsl, S.D.; Smith, S.N.; Ernst, R.D.; Rasko, D.A.; Mobley, H.L. Host-specific induction of Escherichia coli fitness genes during human urinary tract infection. Proc. Natl. Acad. Sci. USA 2014, 111, 18327–18332. [Google Scholar] [CrossRef] [Green Version]
- Cisneros, J.M.; Rodríguez-Baño, J. Nosocomial bacteremia due to Acinetobacter baumannii: epidemiology, clinical features and treatment. Clin. Microbiol. Infect. 2002, 8, 687–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, A.; O’Donoghue, M.; Feeney, A.; Sleator, R.D. Acinetobacter baumannii: An emerging opportunistic pathogen. Virulence 2012, 3, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.E.; Richet, H. The epidemiology and control of Acinetobacter baumannii in health care facilities. Clin. Infect. Dis. 2006, 42, 692–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, C.L.; Neu, H.M.; Gilbreath, J.J.; Michel, S.L.; Zurawski, D.V.; Merrell, D.S. Copper resistance of the emerging pathogen Acinetobacter baumannii. Appl. Environ. Microbiol. 2016, 82, 6174–6188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, K.A.; Pederick, V.G.; Elbourne, L.D.H.; Paulsen, I.T.; Paton, J.C.; McDevitt, C.A.; Eijkelkamp, B.A. Zinc stress induces copper depletion in Acinetobacter baumannii. BMC Microbiol. 2017, 17, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Jabri, Z.; Zamudio, R.; Horvath-Papp, E.; Ralph, J.D.; Al-Muharrami, Z.; Rajakumar, K.; Oggioni, M.R. Integrase-controlled excision of metal-resistance genomic islands in Acinetobacter baumannii. Genes 2018, 9, 366. [Google Scholar] [CrossRef] [Green Version]
- Alquethamy, S.F.; Khorvash, M.; Pederick, V.G.; Whittall, J.J.; Paton, J.C.; Paulsen, I.T.; Hassan, K.A.; McDevitt, C.A.; Eijkelkamp, B.A. The role of the CopA copper efflux system in Acinetobacter baumannii virulence. Int. J. Mol. Sci. 2019, 20, 575. [Google Scholar] [CrossRef] [Green Version]
- Di Nocera, P.P.; Rocco, F.; Giannouli, M.; Triassi, M.; Zarrilli, R. Genome organization of epidemic Acinetobacter baumannii strains. BMC Microbiol. 2011, 11, 224. [Google Scholar] [CrossRef] [Green Version]
- Leungtongkam, U.; Thummeepak, R.; Tasanapak, K.; Sitthisak, S. Acquisition and transfer of antibiotic resistance genes in association with conjugative plasmid or class 1 integrons of Acinetobacter baumannii. PLoS ONE 2018, 13, e0208468. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Insitute. Performance Standards for Antimicrobial Susceptibility Testing. In CLSI Document M100-S27; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Álvarez-Fraga, L.; Pérez, A.; Rumbo-Feal, S.; Merino, M.; Vallejo, J.A.; Ohneck, E.J.; Edelmann, R.E.; Beceiro, A.; Vázquez-Ucha, J.C.; Valle, J.; et al. Analysis of the role of the LH92_11085 gene of a biofilm hyper-producing Acinetobacter baumannii strain on biofilm formation and attachment to eukaryotic cells. Virulence 2016, 7, 443–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, N.A.; Fass, J.N. No Title. Sick. A Sliding-Window, Adapt. Qual. Trimming Tool FastQ Files. (Version 1.33) [Software]. 2011. Available online: https//github.com/najoshi/sickle (accessed on 20 March 2019).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 9, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Assefa, S.; Keane, T.M.; Otto, T.D.; Newbold, C.; Berriman, M. ABACAS: Algorithm-based automatic contiguation of assembled sequences. Bioinformatics 2009, 25, 1968–1969. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, H.; Nakae, T. Protein C (OprC) of the outer membrane of Pseudomonas aeruginosa is a copper-regulated channel protein. Microbiology 1996, 142, 2137–2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D.G.J. BacMet: Antibacterial biocide and metal resistance genes database. Nucleic Acids Res. 2014, 42, 737–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Bagos, P.G.; Liakopoulos, T.D.; Spyropoulos, I.C.; Hamodrakas, S.J. PRED-TMBB: A web server for predicting the topology of beta-barrel outer membrane proteins. Nucleic Acids Res. 2004, 32, W400–W404. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Garcia-Vallvé, S.; Palau, J.; Romeu, A. Horizontal gene transfer in glycosyl hydrolases inferred from codon usage in Escherichia coli and Bacillus subtilis. Mol. Biol. Evol. 1999, 16, 1125–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.E.; Agouri, S.R.; Tyrrell, J.M.; Walsh, T.R. Heavy metal resistance genes are associated with blaNDM-1 and blaCTX-M-15-carrying Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, e02642. [Google Scholar] [CrossRef] [Green Version]
- Dweba, C.C.; Zishiri, O.T.; El Zowalaty, M.E. Isolation and molecular identification of virulence, antimicrobial and heavy metal resistance genes in livestock-associated methicillin-resistant Staphylococcus aureus. Pathogens 2019, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Yadav, K.K.; Mandal, A.K.; Chakraborty, R. Copper susceptibility in Acinetobacter junii BB1A is related to the production of extracellular polymeric substances. Antonie Leeuwenhoek 2013, 104, 261–269. [Google Scholar] [CrossRef]
- Espírito-Santo, C.; Lam, E.W.; Elowsky, C.G.; Quaranta, D.; Domaille, D.W.; Chang, C.J.; Grass, G. Bacterial killing by dry metallic copper surfaces. Appl. Environ. Microbiol. 2011, 77, 794–802. [Google Scholar] [CrossRef] [Green Version]
- Farrugia, D.N.; Elbourne, L.D.; Mabbutt, B.C.; Paulsen, I.T. A novel family of integrases associated with prophages and genomic islands integrated within the tRNA-dihydrouridine synthase A (dusA) gene. Nucleic Acids Res. 2015, 43, 4547–4557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, E.; Freitas, A.R.; Antunes, P.; Barros, M.; Campos, J.; Coque, T.M.; Peixe, L.; Novais, C. Co-transfer of resistance to high concentrations of copper and first-line antibiotics among Enterococcus from different origins (humans, animals, the environment and foods) and clonal lineages. J. Antimicrob. Chemother. 2014, 6, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Li, L.G.; Xia, Y.; Zhang, T. Co-occurrence of antibiotic and metal resistance genes revealed in complete genome collection. ISME J. 2017, 11, 651–662. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, S.; Rasooli, I.; Mousavi-Gargari, S.L. The role of TonB-dependent copper receptor in virulence of Acinetobacter baumannii. Infect. Genet. Evol. 2018, 60, 181–190. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
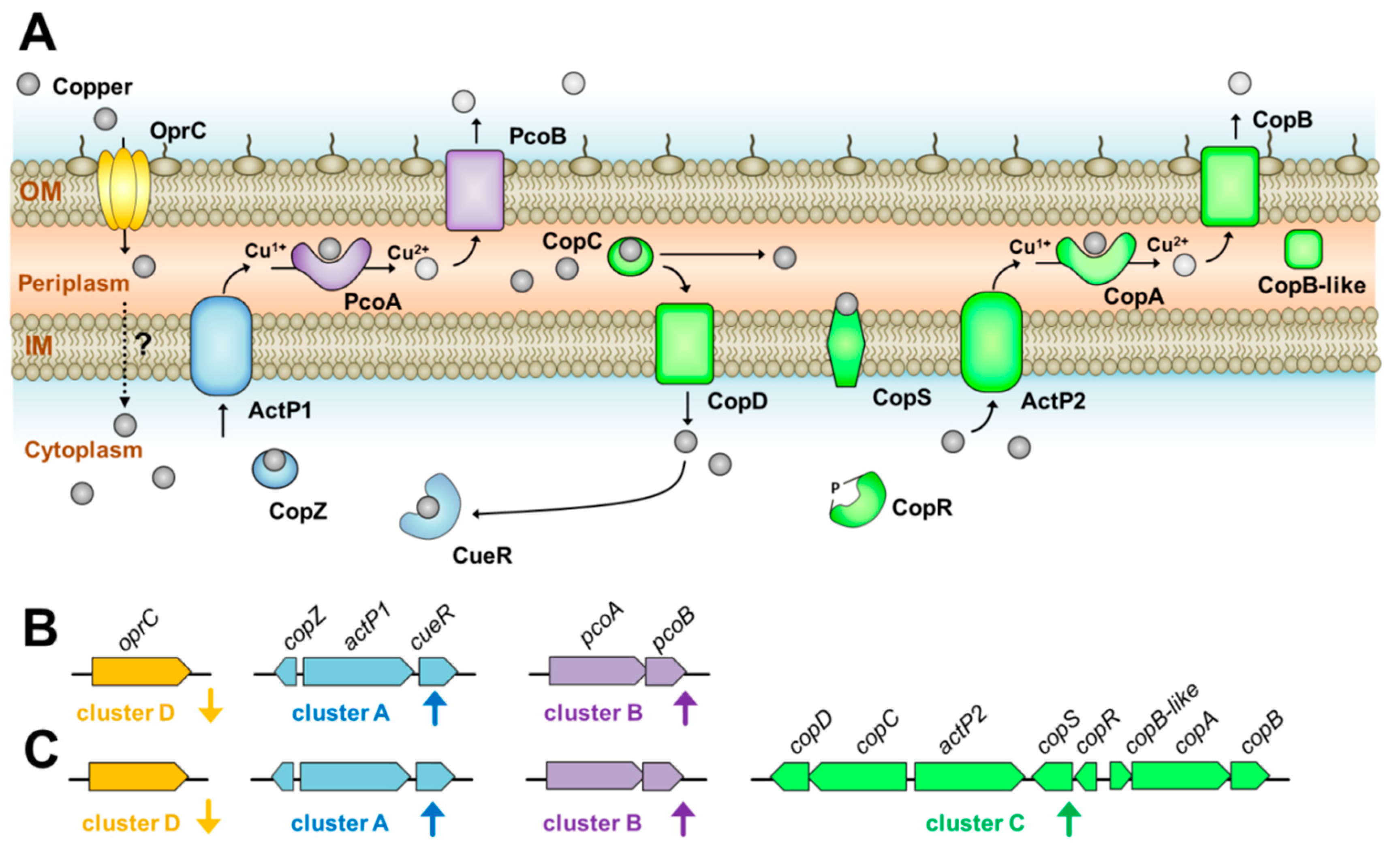{kind=link}
| Patterns of Copper-Related Genes a | Total Isolates (%) | Copper Susceptible Isolates (MIC = 5 mM) (%) | Copper-Tolerant Isolates b (MIC = 10 mM) (%) |
|---|---|---|---|
| All isolates | n = 407 (100) | 311 (100) | 96 (100) |
| cueR+/pcoAB+/copRS-/oprC+ | 323 (79.4) | 311 (100) | 12 (12.5) |
| cueR+/pcoAB+/copRS+/oprC+ | 84 (20.6) | 0 (0) | 84 (87.5) * |
| Environmental isolates | n = 13 (100) | 7 (100) | 6 (100) |
| cueR+/pcoAB+/copRS-/oprC+ | 7 (53.8) | 7 (100) | 0 (0) |
| cueR+/pcoAB+/copRS+/oprC+ | 6 (46.2) | 0 (0) | 6 (100) * |
| Clinical isolates | n = 394 (100) | 304 (100) | 90 (100) |
| cueR+/pcoAB+/copRS-/oprC+ | 316 (80.2) | 304 (100) | 12 (13.3) |
| cueR+/pcoAB+/copRS+/oprC+ | 78 (19.8) | 0 (0) | 78 (86.7) * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thummeepak, R.; Pooalai, R.; Harrison, C.; Gannon, L.; Thanwisai, A.; Chantratita, N.; Millard, A.D.; Sitthisak, S. Essential Gene Clusters Involved in Copper Tolerance Identified in Acinetobacter baumannii Clinical and Environmental Isolates. Pathogens 2020, 9, 60. https://doi.org/10.3390/pathogens9010060
Thummeepak R, Pooalai R, Harrison C, Gannon L, Thanwisai A, Chantratita N, Millard AD, Sitthisak S. Essential Gene Clusters Involved in Copper Tolerance Identified in Acinetobacter baumannii Clinical and Environmental Isolates. Pathogens. 2020; 9(1):60. https://doi.org/10.3390/pathogens9010060
Chicago/Turabian StyleThummeepak, Rapee, Renuka Pooalai, Christian Harrison, Lucy Gannon, Aunchalee Thanwisai, Narisara Chantratita, Andrew D. Millard, and Sutthirat Sitthisak. 2020. "Essential Gene Clusters Involved in Copper Tolerance Identified in Acinetobacter baumannii Clinical and Environmental Isolates" Pathogens 9, no. 1: 60. https://doi.org/10.3390/pathogens9010060