Antileishmanial Activity and Synergistic Effects of Amphotericin B Deoxycholate with Allicin and Andrographolide against Leishmania martiniquensis In Vitro

 ,
,
Abstract
:
1. Introduction
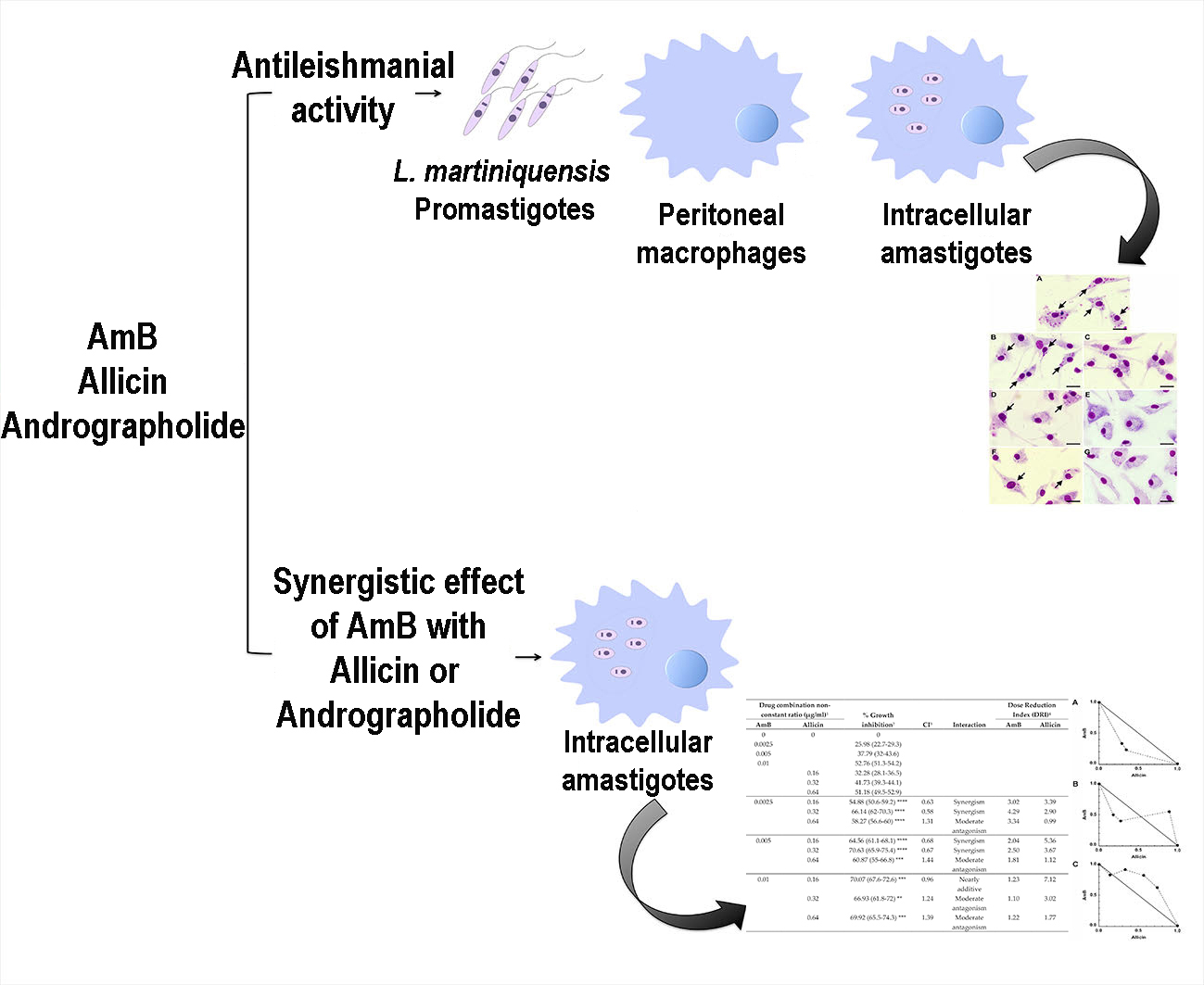
2. Results
2.1. Antileishmanial Activity against Promastigotes
2.2. Cytotoxicity on BALB/c Peritoneal Macrophages
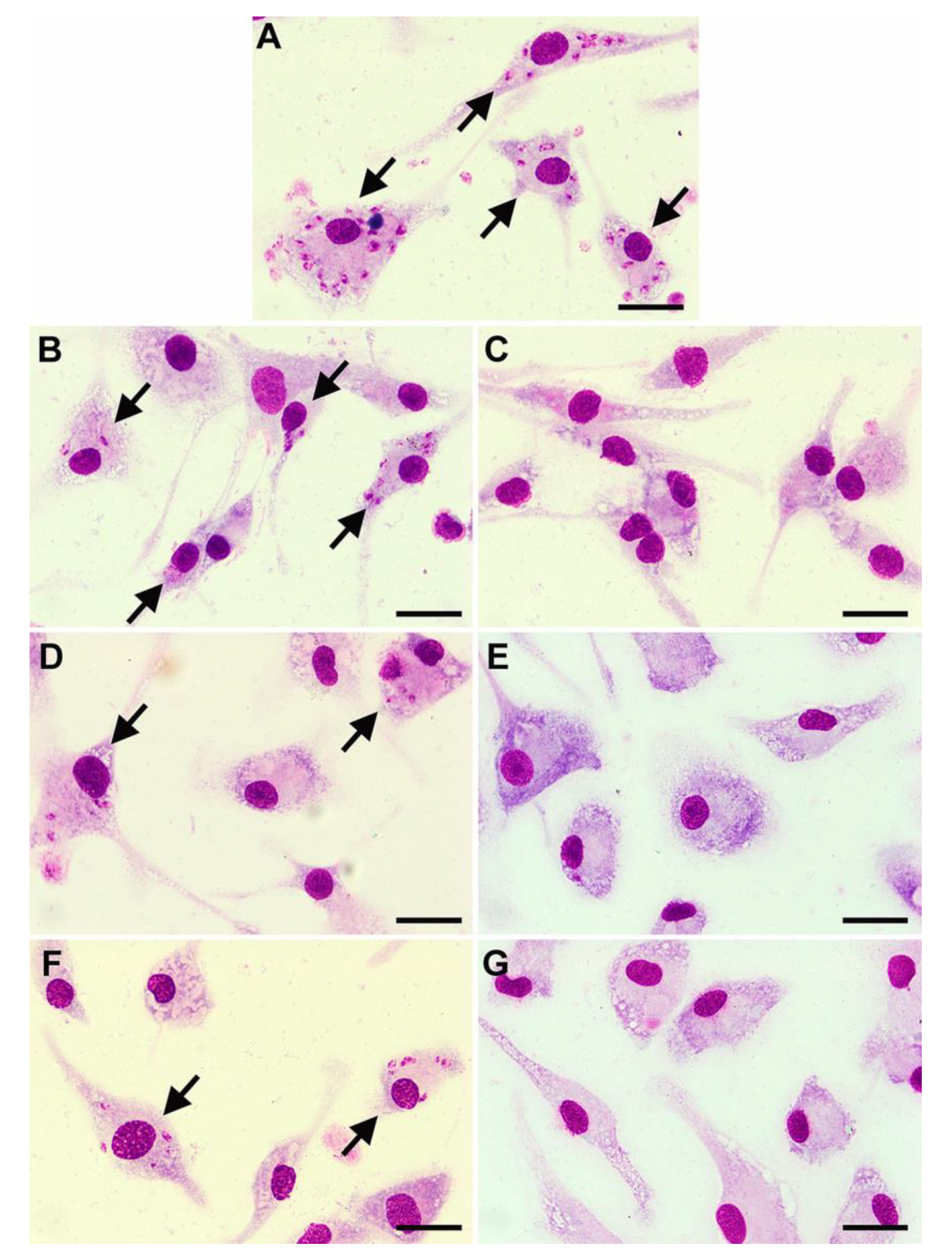
2.3. Antileishmanial Activity against Intracellular Amastigotes
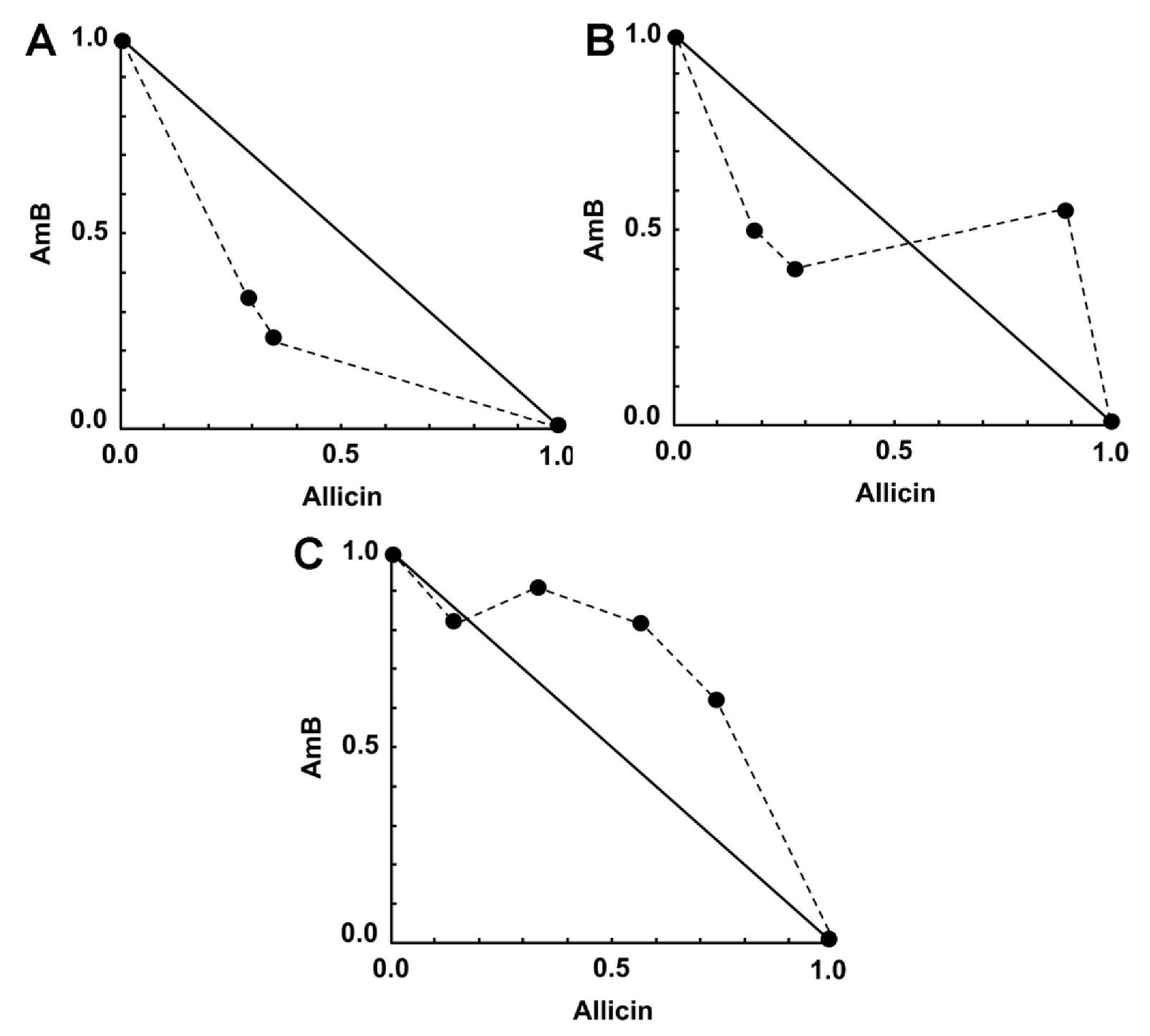
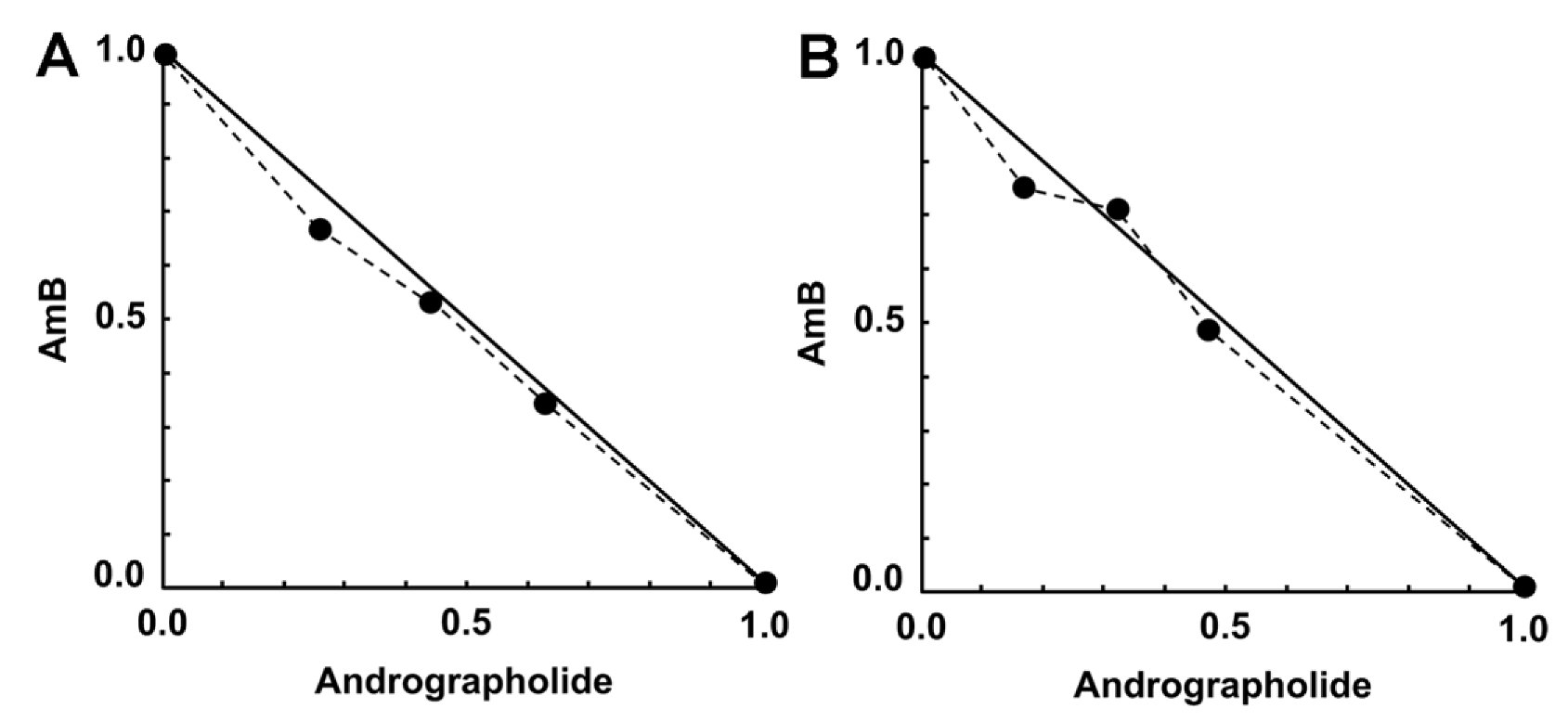
2.4. Activity of Synergistic Combinations against Intracellular Amastigotes
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Parasites and Culture
4.3. Drug and Compounds
4.4. Promastigote Assay
4.5. Cytotoxicity Assay
4.6. Preparation of Promastigotes to Infect Murine Macrophages
4.7. Intracellular Amastigote Assay
4.8. Drug Combination Assay on Intracellular Amastigotes
4.9. Analysis of Interaction
4.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Leelayoova, S.; Siripattanapipong, S.; Manomat, J.; Piyaraj, P.; Tan-Ariya, P.; Bualert, L.; Mungthin, M. Leishmaniasis in Thailand: A review of causative agents and situations. Am. J. Trop. Med. Hyg. 2017, 96, 534–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jariyapan, N.; Daroontum, T.; Jaiwong, K.; Chanmol, W.; Intakhan, N.; Sor-Suwan, S.; Siriyasatien, P.; Somboon, P.; Bates, M.D.; Bates, P.A. Leishmania (Mundinia) orientalis n. sp. (Trypanosomatidae), a parasite from Thailand responsible for localised cutaneous leishmaniasis. Parasit. Vectors 2018, 11, 351. [Google Scholar] [CrossRef] [PubMed]
- Sundar, S.; Singh, O.P.; Chakravarty, J. Visceral leishmaniasis elimination targets in India, strategies for preventing resurgence. Expert Rev. Anti. Infect. Ther. 2018, 16, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Manomat, J.; Leelayoova, S.; Bualert, L.; Tan-ariya, P.; Siripattanapipong, S.; Mungthin, M.; Naaglor, T.; Piyaraj, P. Prevalence and risk factors associated with Leishmania infection in Trang Province, Southern Thailand. PLoS Negl. Trop. Dis. 2017, 11, e00060995. [Google Scholar] [CrossRef] [Green Version]
- Dedet, J.P.; Roche, B.; Pratlong, F.; Cales-Quist, D.; Jouannelle, J.; Benichou, J.C.; Huerre, M. Diffuse cutaneous infection caused by a presumed monoxenous trypanosomatid in a patient infected with HIV. Trans. R. Soc. Trop. Med. Hyg. 1995, 89, 644–646. [Google Scholar] [CrossRef]
- Desbois, N.; Pratlong, F.; Quist, D.; Dedet, J.P. Leishmania (Leishmania) martiniquensis n. sp. (Kinetoplastida: Trypanosomatidae), description of the parasite responsible for cutaneous leishmaniasis in Martinique Island (French West Indies). Parasite 2014, 21, 12. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, O.A.; Serrano, M.G.; Camargo, E.P.; Teixeira, M.M.G.; Shaw, J.J. An appraisal of the taxonomy and nomenclature of trypanosomatids presently classified as Leishmania and Endotrypanum. Parasitology 2018, 145, 430–442. [Google Scholar] [CrossRef]
- Pothirat, T.; Tantiworawit, A.; Chaiwarith, R.; Jariyapan, N.; Wannasan, A.; Siriyasatien, P.; Supparatpinyo, K.; Bates, M.D.; Kwakye-Nuako, G.; Bates, P.A. First isolation of Leishmania from Northern Thailand: Case report, identification as Leishmania martiniquensis and phylogenetic position within the Leishmania enriettii complex. PLoS Negl. Trop. Dis. 2014, 8, e3339. [Google Scholar] [CrossRef]
- Chusri, S.; Hortiwakul, T.; Silpapojakul, K.; Siriyasatien, P. Consecutive cutaneous and visceral leishmaniasis manifestations involving a novel Leishmania species in two HIV patients in Thailand. Am. J. Trop. Med. Hyg. 2012, 87, 76–80. [Google Scholar] [CrossRef]
- Liautaud, B.; Vignier, N.; Miossec, C.; Plumelle, Y.; Kone, M.; Delta, D.; Ravel, C.; Cabié, A.; Desbois, N. First case of visceral leishmaniasis caused by Leishmania martiniquensis. Am. J. Trop. Med. Hyg. 2015, 92, 317–319. [Google Scholar] [CrossRef] [Green Version]
- Osatakul, S.; Mungthin, M.; Siripattanapipong, S.; Hitakarun, A.; Kositnitikul, R.; Naaglor, T.; Leelayoova, S. Recurrences of visceral leishmaniasis caused by Leishmania siamensis after treatment with amphotericin B in a seronegative child. Am. J. Trop. Med. Hyg. 2014, 90, 40–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laniado-Laborín, R.; Cabrales-Vargas, M.N. Amphotericin B: Side effects and toxicity. Rev Iberoam. Micol. 2009, 26, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Sundar, S.; Chakravarty, J. An update on pharmacotherapy for leishmaniasis. Expert Opin. Pharmacother. 2015, 16, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Jesús Corral-Caridad, M.; Moreno, I.; Toraño, A.; Domínguez, M.; Alunda, J.M. Effect of allicin on promastigotes and intracellular amastigotes of Leishmania donovani and L. infantum. Exp. Parasitol. 2012, 132, 475–482. [Google Scholar] [CrossRef]
- Corral, M.J.; González-Sánchez, E.; Cuquerella, M.; Alunda, J.M. In vitro synergistic effect of amphotericin B and allicin on Leishmania donovani and L. infantum. Antimicrob. Agents Chemother. 2014, 58, 1596–1602. [Google Scholar] [CrossRef] [Green Version]
- McClure, D.C.; Noland, L.L.; Zatyrka, S.A. Antileishmanial properties of Allium sativum extracts and derivatives. Acta Hortic. 1996, 426, 183–191. [Google Scholar] [CrossRef]
- Metwally, D.M.; Al-Olayan, E.M.; El-Khadragy, M.F.; Alkathiri, B. Anti-Leishmanial activity (in vitro and in vivo) of allicin and allicin cream using Leishmania major (Sub-strain Zymodeme LON4) and Balb/c mice. PLoS ONE 2016, 11, e0161296. [Google Scholar] [CrossRef]
- Sinha, J.; Mukhopadhyay, S.; Das, N.; Basu, M.K. Targeting of liposomal andrographolide to L. donovani-infected macrophages In Vivo. Drug Deliv. 2000, 7, 209–213. [Google Scholar]
- Mishra, K.; Dash, A.P.; Dey, N. Andrographolide: A novel antimalarial diterpene lactone compound from Andrographis paniculata and its interaction with curcumin and artesunate. J. Trop. Med. 2011, 2011, 579518. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, M.; Parai, D.; Dhar, P.; Roy, M.; Barik, R.; Chattopadhyay, S.; Mukherjee, SK. Andrographolide induces oxidative stress-dependent cell death in unicellular protozoan parasite Trypanosoma brucei. Acta Trop. 2017, 176, 58–67. [Google Scholar]
- Yardley, V.; Croft, S.L. Activity of liposomal Amphotericin B against experimental cutaneous leishmaniasis. Antimicrob. Agents Chemother. 1997, 41, 752–756. [Google Scholar] [CrossRef] [Green Version]
- Al-Mohammed, H.I.; Chance, M.L.; Bates, P.A. Production and characterization of stable Amphotericin-resistant amastigotes and promastigotes of Leishmania mexicana. Antimicrob. Agents Chemother. 2005, 49, 3274–3280. [Google Scholar] [CrossRef] [Green Version]
- Don, R.; Loset, J.R. Screening strategies to identify new chemical diversity for drug development to treat kinetoplastid infection. Parasitology 2014, 141, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Katsuno, K.; Burrows, J.N.; Duncan, K.; Hooft van Huijsduijnen, R.; Kaneko, T.; Kita, K.; Mowbray, C.E.; Schmatz, D.; Warner, P.; Slingsby, B.T. Hit and lead criteria in drug discovery for infectious diseases of the developing world. Nat. Rev. Drug Discov. 2015, 14, 751–758. [Google Scholar] [CrossRef]
- An, M.; Shen, H.; Cao, Y.; Zhang, J.; Cai, Y.; Wang, R.; Jiang, Y. Allicin enhances the oxidative damage effect of amphotericin B against Candida albicans. Int. J. Antimicrob. Agents. 2009, 33, 258–263. [Google Scholar] [CrossRef]
- Corral, M.J.; Benito-Peña, E.; Jiménez-Antón, M.D.; Cuevas, L.; Moreno-Bondi, M.C.; Alunda, J.M. Allicin induces calcium and mitochondrial dysregulation causing necrotic death in Leishmania. PLoS Negl. Trop. Dis. 2016, 10, e0004525. [Google Scholar] [CrossRef] [Green Version]
- Rabinkov, A.; Miron, T.; Konstantinovski, L.; Wilchek, M.; Mirelman, D.; Weiner, L. The mode of action of allicin: Trapping of radicals and interaction with thiol containing proteins. Biochim. Biophys. Acta 1998, 1379, 233–244. [Google Scholar] [CrossRef]
- Krstin, S.; Sobeh, M.; Bruan, M.S.; Wink, M. Antiparasitic activity of Allium sativum and Allium cepa against Trypanosoma b. brucei and Leishmania tarentolae. Medicines 2018, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Miron, T.; Rabinkov, A.; Mirelman, D.; Wilchek, M.; Weiner, L. The mode of action of allicin: Its ready permeability through phospholipid membranes may contribute to its biological activity. Biochim. Biophys. Acta 2000, 1463, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Haase, H.; Hieke, N.; Plum, L.M.; Gruhlke, M.; Slusarenko, A.J.; Rink, L. Impact of allicin on macrophage activity. Food Chem. 2012, 134, 141–148. [Google Scholar] [CrossRef]
- Sundar, S.; Chatterjee, M. Visceral leishmaniasis-current therapeutic modalities. Indian J. Med. Res. 2006, 123, 345–352. [Google Scholar]
- Sundar, S.; Mehta, H.; Chhabra, A.; Singh, V.; Chauhan, V.; Desjeux, P.; Rai, M. Amphotericin B colloidal dispersion for the treatment of Indian visceral leishmaniasis. Clin. Infect. Dis. 2006, 42, 608–613. [Google Scholar] [CrossRef]
- Bao, G.Q.; Shen, B.Y.; Pan, C.P.; Zhang, Y.J.; Shi, M.M.; Peng, C.H. Andrographolide causes apoptosis via inactivation of STAT3 and Akt and potentiates antitumor activity of gemcitabine in pancreatic cancer. Toxicol. Lett. 2013, 222, 23–35. [Google Scholar] [CrossRef]
- Yunos, N.M.; Mutalip, S.S.; Jauri, M.H.; Yu, J.Q.; Huq, F. Anti-proliferative and pro-apoptotic effects from sequenced combinations of andrographolide and cisplatin on ovarian cancer cell lines. Anticancer Res. 2013, 33, 4365–4371. [Google Scholar]
- Guo, H.; Zhang, Z.; Su, Z.; Sun, C.; Zhang, X.; Zhao, X.; Lai, X.; Su, Z.; Li, Y.; Zhan, J.Y. Enhanced anti-tumor activity and reduced toxicity by combination andrographolide and bleomycin in ascitic tumor-bearing mice. Eur. J. Pharmacol. 2016, 776, 52–63. [Google Scholar] [CrossRef]
- Yuan, H.; Sun, B.; Gao, F.; Lan, M. Synergistic anticancer effects of andrographolide and paclitaxel against A549 NSCLC cells. Pharm. Biol. 2016, 54, 2629–2635. [Google Scholar] [CrossRef] [Green Version]
- Alzaharna, M.; Alqouqa, I.; Cheung, H.Y. Taxifolin synergizes Andrographolide-induced cell death by attenuation of autophagy and augmentation of caspase dependent and independent cell death in HeLa cells. PLoS ONE 2017, 12, e0171325. [Google Scholar] [CrossRef] [Green Version]
- Roy, P.; Das, S.; Bera, T.; Mondol, S.; Mukherjee, A. Andrographolide nanoparticles in leishmaniasis: Characterization and in vitro evaluations. Int. J. Nanomed. 2010, 5, 1113–1121. [Google Scholar]
- Chiewchanvit, S.; Tovanabutra, N.; Jariyapan, N.; Bates, M.D.; Mahanupab, P.; Chuamanochan, M.; Tantiworawit, A.; Bates, P.A. Chronic generalized fibrotic skin lesions from disseminated leishmaniasis caused by Leishmania martiniquensis in two patients from northern Thailand infected with HIV. Br. J. Dermatol. 2015, 173, 663–670. [Google Scholar] [CrossRef]
- Van Bockstal, L.; Gielis, J.F.; Delputte, P.; Cos, P.; Maes, L.; Caljon, G.; Hendrickx, S. Impact of primary mouse macrophage cell types on Leishmania infection and in vitro drug susceptibility. Parasitol Res. 2018, 117, 3601–3612. [Google Scholar]
- Zhang, X.; Goncalves, R.; Mosser, D.M. The isolation and characterization of murine macrophages. Curr. Protoc. Immunol. 2008, 14, 14–21. [Google Scholar] [CrossRef]
- Zakai, H.A.; Chance, M.L.; Bates, P.A. In vitro stimulation of metacyclogenesis in Leishmania braziliensis, L. donovani, L. major and L. mexicana. Parasitology 1998, 116, 305–309. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, A.K.; Valero, V.B.; Takahashi, H.K.; Straus, A.H. Inhibition of Leishmania (Leishmania) amazonensis growth and infectivity by aureobasidin A. J. Antimicrob. Chemother. 2007, 59, 487–492. [Google Scholar] [CrossRef]
- Chou, T.C. Preclinical versus clinical drug combination studies. Leuk Lymphoma. 2008, 49, 2059–2080. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug Combination Non-Constant Ratio (μg/mL) 1 | % Growth Inhibition 2 | CI 3 | Interaction | Dose Reduction Index (DRI) 4 | ||
|---|---|---|---|---|---|---|
| AmB | Allicin | AmB | Allicin | |||
| 0 | 0 | 0 | ||||
| 0.0025 | 25.98 (22.7–29.3) | |||||
| 0.005 | 37.79 (32–43.6) | |||||
| 0.01 | 52.76 (51.3–54.2) | |||||
| 0.16 | 32.28 (28.1–36.5) | |||||
| 0.32 | 41.73 (39.3–44.1) | |||||
| 0.64 | 51.18 (49.5–52.9) | |||||
| 0.0025 | 0.16 | 54.88 (50.6–59.2) *** | 0.63 | Synergism | 3.02 | 3.39 |
| 0.0025 | 0.32 | 66.14 (62–70.3) *** | 0.58 | Synergism | 4.29 | 2.90 |
| 0.0025 | 0.64 | 58.27 (56.6–60) *** | 1.31 | Moderate antagonism | 3.34 | 0.99 |
| 0.005 | 0.16 | 64.56 (61.1–68.1) *** | 0.68 | Synergism | 2.04 | 5.36 |
| 0.005 | 0.32 | 70.63 (65.9–75.4) *** | 0.67 | Synergism | 2.50 | 3.67 |
| 0.005 | 0.64 | 60.87 (55–66.8) ** | 1.44 | Moderate antagonism | 1.81 | 1.12 |
| 0.01 | 0.16 | 70.07 (67.6–72.6) ** | 0.96 | Nearly additive | 1.23 | 7.12 |
| 0.01 | 0.32 | 66.93 (61.8–72) * | 1.24 | Moderate antagonism | 1.10 | 3.02 |
| 0.01 | 0.64 | 69.92 (65.5–74.3) ** | 1.39 | Moderate antagonism | 1.22 | 1.77 |
| Drug Combination Non-Constant Ratio (μg/mL) 1 | % Growth Inhibition 2 | CI 3 | Interaction | Dose Reduction Index (DRI) 4 | ||
|---|---|---|---|---|---|---|
| AmB | Andrographolide | AmB | Andrographolide | |||
| 0 | 0 | 0 | ||||
| 0.0025 | 24.5 (21–28) | |||||
| 0.005 | 38.4 (32.3–44.5) | |||||
| 0.01 | 51.72 (48.8–54.7) | |||||
| 0.08 | 4.72 (2.68–6.76) | |||||
| 0.16 | 12.6 (9.88–15.3) | |||||
| 0.32 | 44.88 (38.7–51.1) | |||||
| 0.64 | 53.78 (50.7–56.8) | |||||
| 0.0025 | 0.08 | 31.5 (27.8–35.2) | 0.93 | Nearly additive | 1.51 | 3.79 |
| 0.0025 | 0.16 | 37.8 (33.7–41.9) ** | 0.98 | Nearly additive | 1.87 | 2.25 |
| 0.0025 | 0.32 | 51.18 (47.9–54.5) **** | 0.98 | Nearly additive | 2.87 | 1.58 |
| 0.0025 | 0.64 | 59.05 (55.5–62.6) **** | 1.31 | Moderate antagonism | 3.67 | 0.96 |
| 0.005 | 0.08 | 48.81 (43.8–53.8) * | 0.92 | Nearly additive | 1.33 | 5.95 |
| 0.005 | 0.16 | 50.63 (44.8–56.5) * | 1.03 | Nearly additive | 1.41 | 3.11 |
| 0.005 | 0.32 | 62.44 (60.3–64.6) *** | 0.96 | Nearly additive | 2.05 | 2.10 |
| 0.005 | 0.64 | 51.18 (49–53.3) * | 1.97 | Antagonism | 1.43 | 0.79 |
| 0.01 | 0.08 | 40.16 (37.1–43.2) ** | 2.18 | Antagonism | 0.51 | 4.79 |
| 0.01 | 0.16 | 48.56 (46.1–51.1) | 1.85 | Antagonism | 0.66 | 2.96 |
| 0.01 | 0.32 | 37.20 (30.8–43.7) *** | 3.08 | Antagonism | 0.46 | 1.11 |
| 0.01 | 0.64 | 44.72 (39.9–49.6) * | 3.19 | Antagonism | 0.58 | 0.67 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Intakhan, N.; Chanmol, W.; Somboon, P.; Bates, M.D.; Yardley, V.; Bates, P.A.; Jariyapan, N. Antileishmanial Activity and Synergistic Effects of Amphotericin B Deoxycholate with Allicin and Andrographolide against Leishmania martiniquensis In Vitro. Pathogens 2020, 9, 49. https://doi.org/10.3390/pathogens9010049
Intakhan N, Chanmol W, Somboon P, Bates MD, Yardley V, Bates PA, Jariyapan N. Antileishmanial Activity and Synergistic Effects of Amphotericin B Deoxycholate with Allicin and Andrographolide against Leishmania martiniquensis In Vitro. Pathogens. 2020; 9(1):49. https://doi.org/10.3390/pathogens9010049
Chicago/Turabian StyleIntakhan, Nuchpicha, Wetpisit Chanmol, Pradya Somboon, Michelle D. Bates, Vanessa Yardley, Paul A. Bates, and Narissara Jariyapan. 2020. "Antileishmanial Activity and Synergistic Effects of Amphotericin B Deoxycholate with Allicin and Andrographolide against Leishmania martiniquensis In Vitro" Pathogens 9, no. 1: 49. https://doi.org/10.3390/pathogens9010049