Immunization of Goats with Recombinant Protein 14-3-3 Isoform 2(rHcftt-2) Induced Moderate Protection against Haemonchus contortus Challenge



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
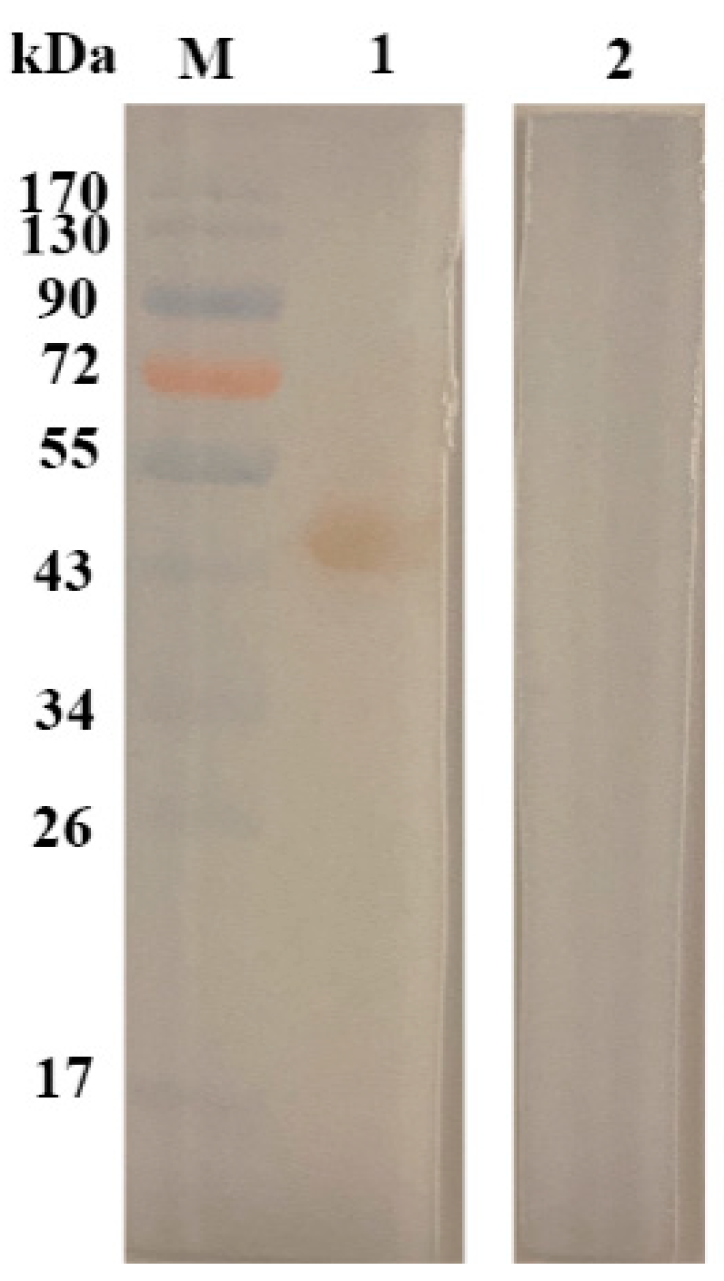
2.1. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis (SDS-PAGE) Profile of rHcftt-2 and Immunoblot
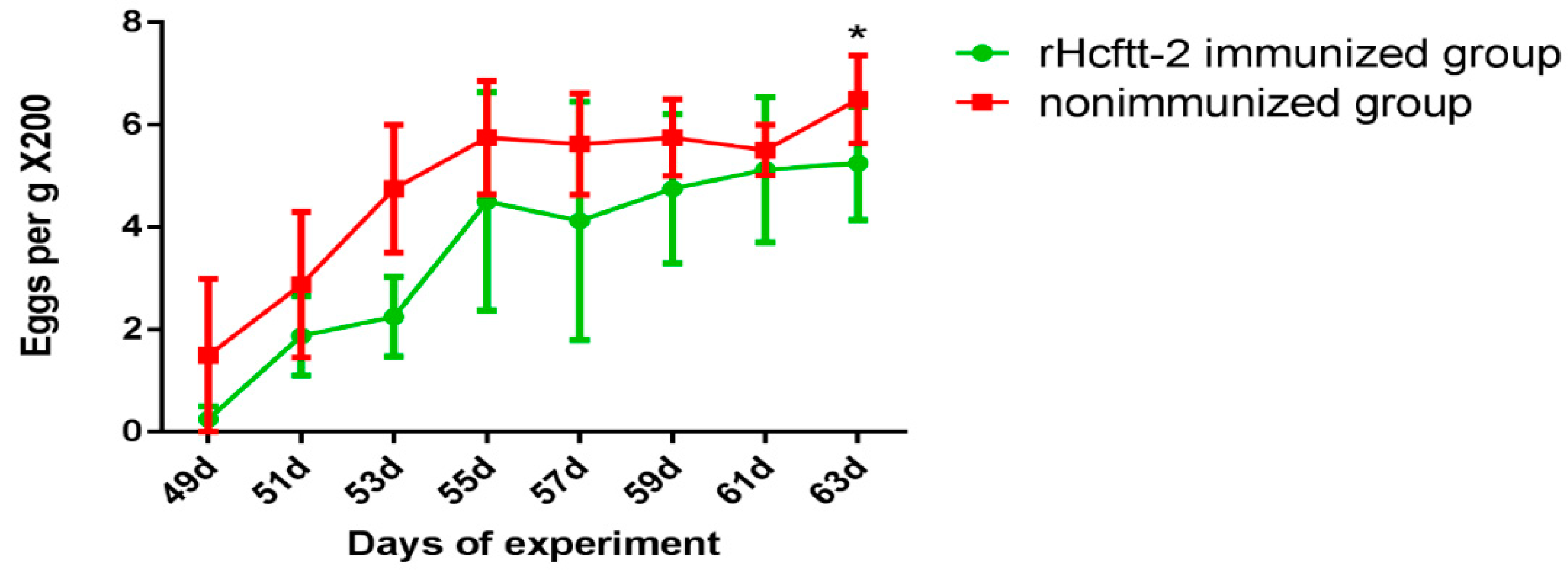
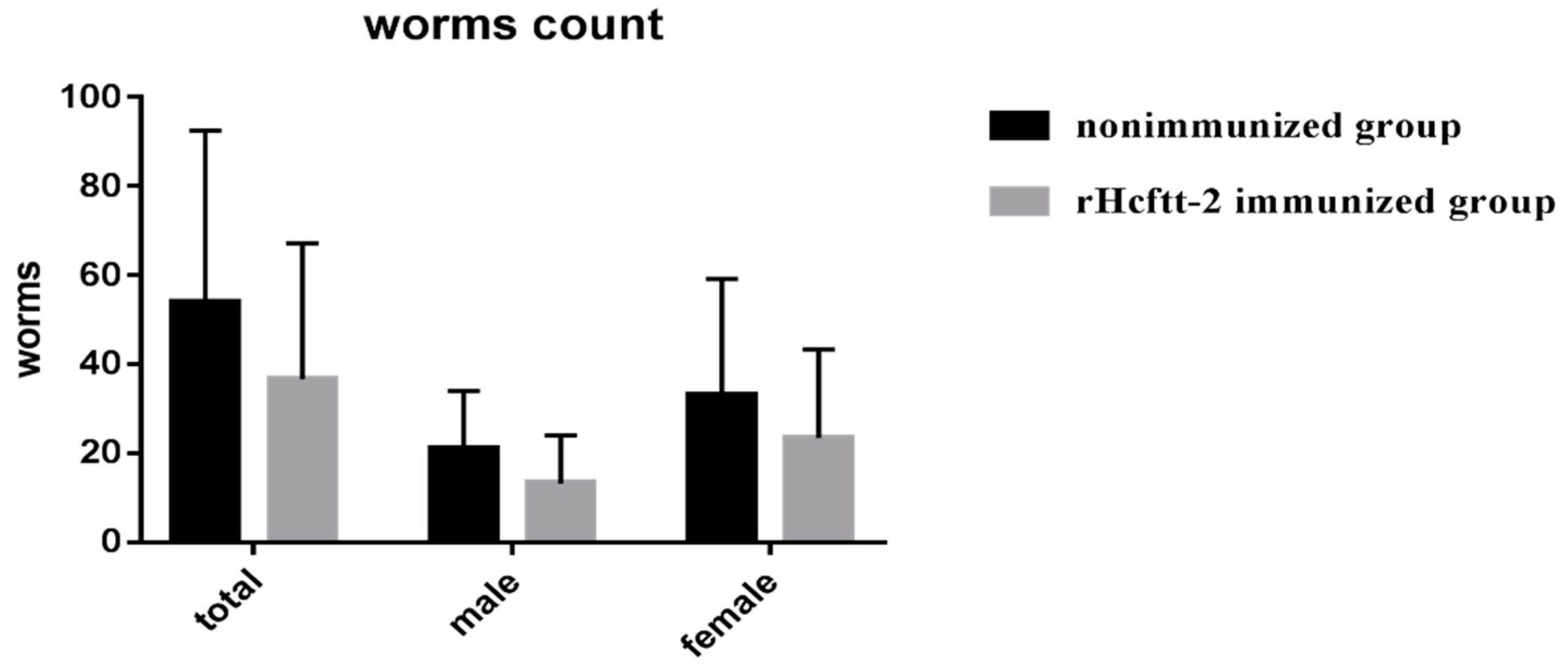
2.2. Fecal Eggs Counts and Worm Burdens
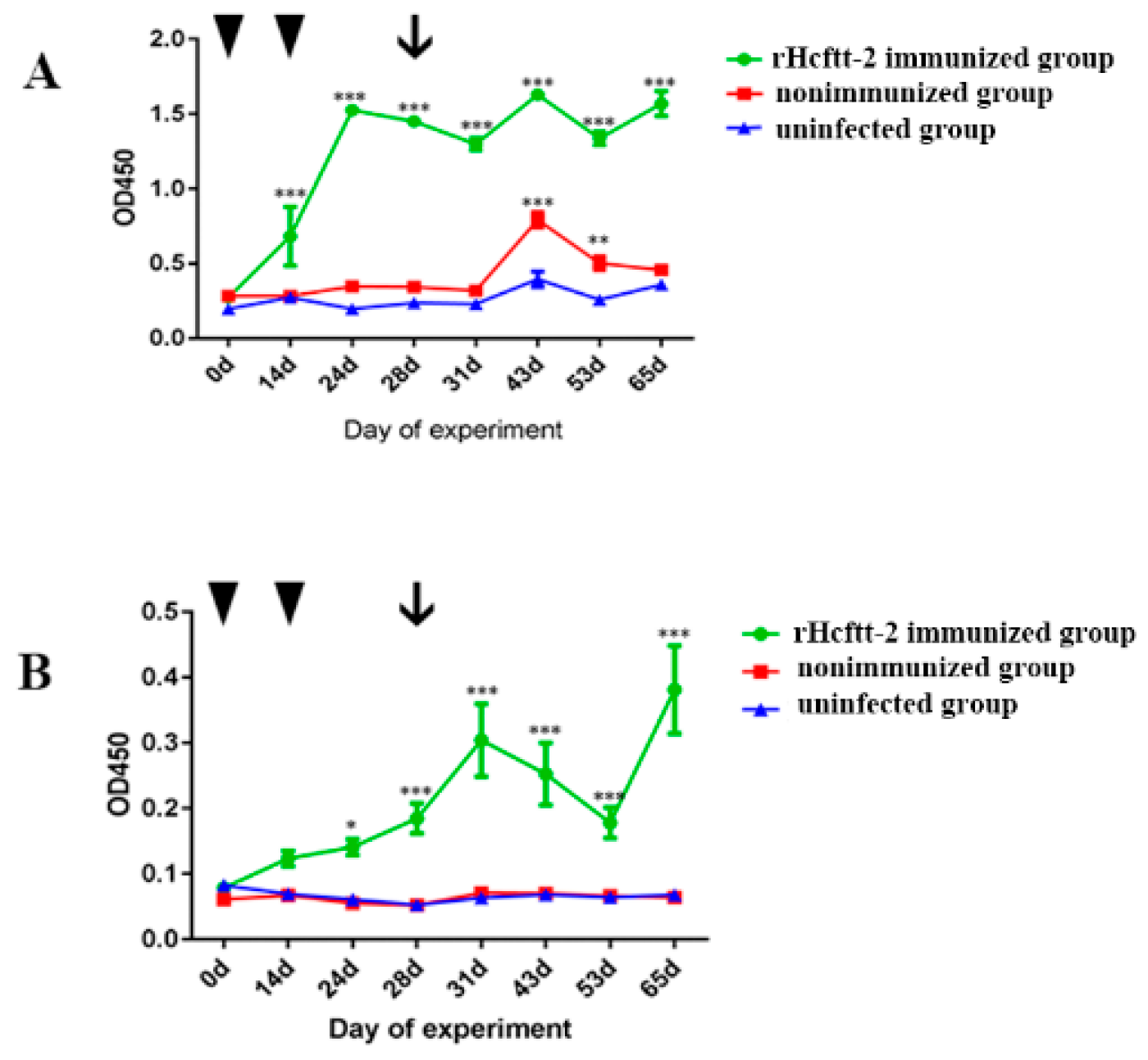
2.3. Serum Antibody Response
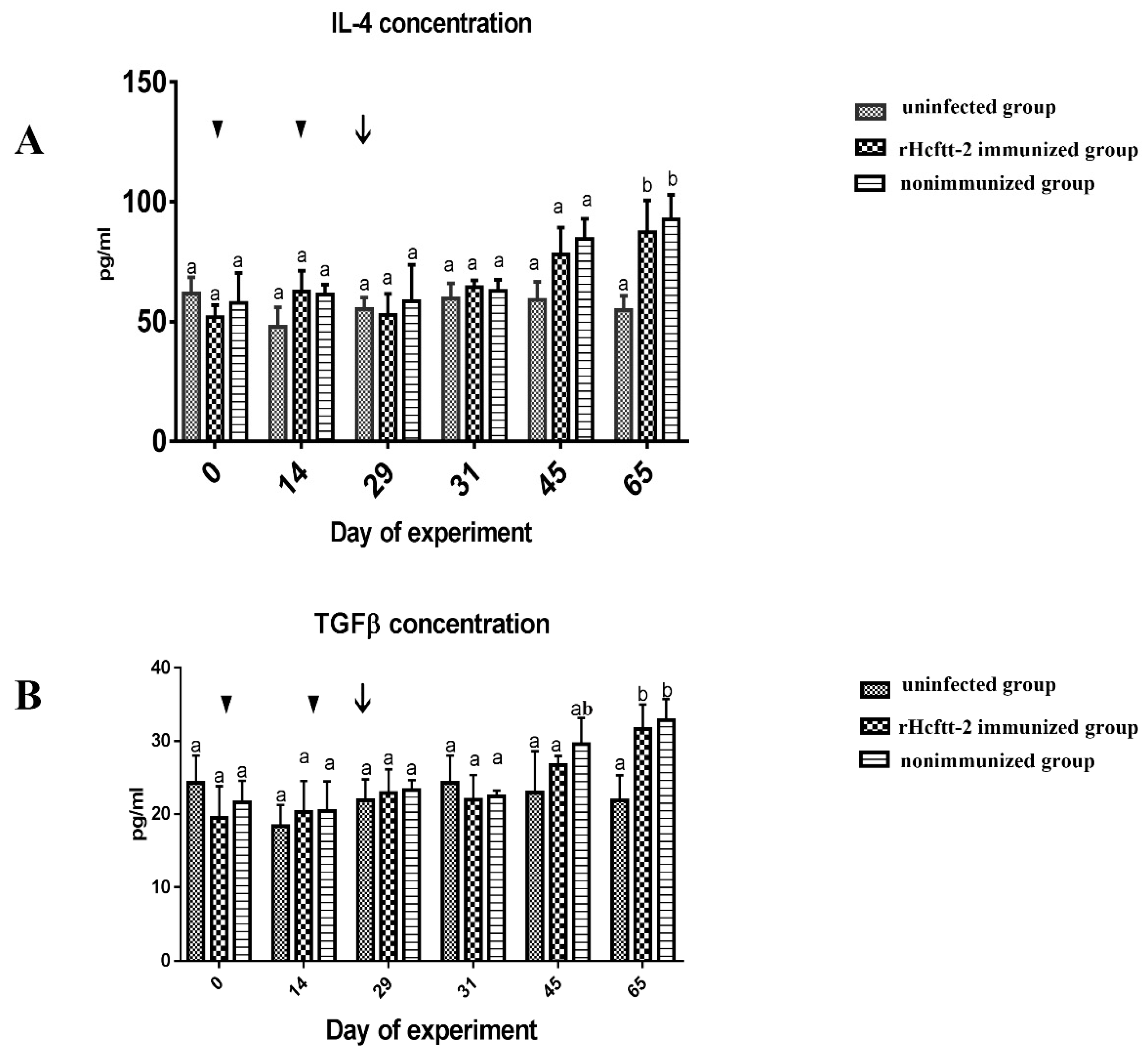
2.4. Concentration of Serum Cytokines
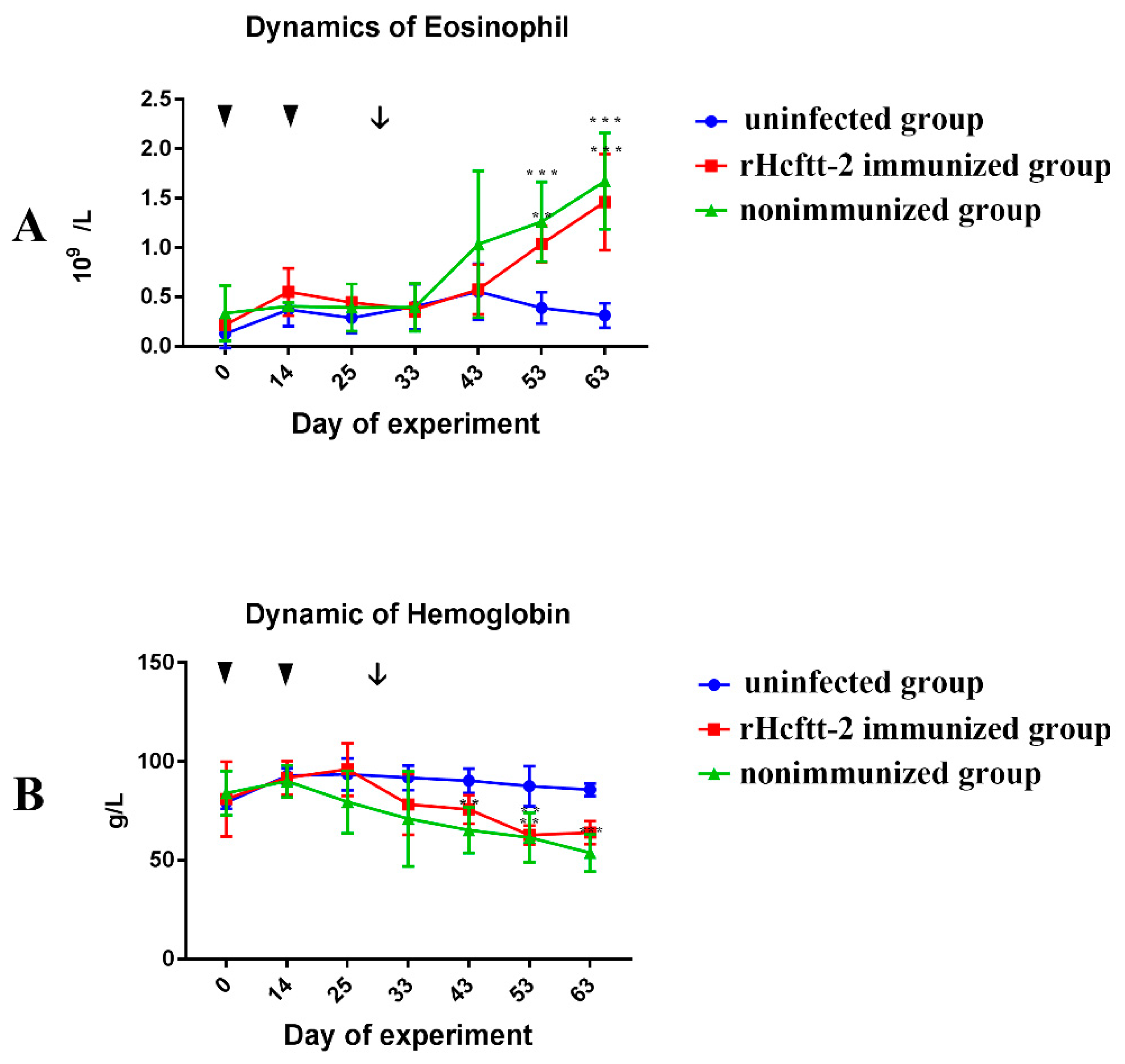
2.5. Differential Cell Counts
3. Discussion
4. Material and Methods
4.1. Infective Larvae for Challenge
4.2. Expression and Purification of rHcftt-2
4.3. Immunoblot Analysis of rHcftt-2 Using Goats Sera
4.4. Animals and Vaccination Protocol
4.5. Parasitological Measurements
4.6. Differential Cell Count
4.7. Detection of Antibodies in Serum
4.8. Determination of Serum Cytokine Concentration
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hoste, H.; Torres-Acosta, J.F.; Quijada, J.; Chan-Perez, I.; Dakheel, M.M.; Kommuru, D.S.; Mueller-Harvey, I.; Terrill, T.H. Interactions Between Nutrition and Infections With Haemonchus contortus and Related Gastrointestinal Nematodes in Small Ruminants. Adv. Parasitol. 2016, 93, 239–351. [Google Scholar]
- Jasmer, D.P.; Lahmers, K.K.; Brown, W.C. Haemonchus contortus intestine: A prominent source of mucosal antigens. Parasite Immunol. 2007, 29, 139–151. [Google Scholar] [CrossRef]
- Kalyanasundaram, A.; Jawahar, S.; Illangopathy, M.; Palavesam, A.; Raman, M. Comparative immunoprophylactic efficacy of Haemonchus contortus recombinant enolase (rHcENO) and Con A purified native glycoproteins in sheep. Exp. Parasitol. 2015, 154, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, F.; Zhang, Z.; Yang, X.; Ahmad, A.A.; Li, X.; Du, A.; Hu, M. Recent Research Progress in China on Haemonchus contortus. Front. Microbiol. 2017, 8, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolstenholme, A.J.; Fairweather, I.; Prichard, R.; Von, S.H.G.; Sangster, N.C. Drug resistance in veterinary helminths. Trends Parasitol. 2004, 20, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Yanming, S.; Ruofeng, Y.; Muleke, C.I.; Guangwei, Z.; Lixin, X.; Xiangrui, L. Vaccination of goats with recombinant galectin antigen induces partial protection against Haemonchus contortus infection. Parasite Immunol. 2007, 29, 319–326. [Google Scholar] [CrossRef]
- Molina, J.M.; Martín, S.; Hernández, Y.I.; González, J.F.; Ferrer, O.; Ruiz, A. Immunoprotective effect of cysteine proteinase fractions from two Haemonchus contortus strains adapted to sheep and goats. Vet. Parasitol. 2012, 188, 53. [Google Scholar] [CrossRef]
- Meier, L.; Torgerson, P.R.; Hertzberg, H. Vaccination of goats against Haemonchus contortus with the gut membrane proteins H11/H-gal-GP. Vet. Parasitol. 2016, 229, 15. [Google Scholar] [CrossRef]
- Fawzi, E.M.; González-Sánchez, M.E.; Corral, M.J.; Alunda, J.M.; Cuquerella, M. Vaccination of lambs with the recombinant protein rHc23 elicits significant protection against Haemonchus contortus challenge. Vet. Parasitol. 2015, 211, 54–59. [Google Scholar] [CrossRef]
- Han, K.; Xu, L.; Yan, R.; Song, X.; Li, X. Vaccination of goats with glyceraldehyde-3-phosphate dehydrogenase DNA vaccine induced partial protection against Haemonchus contortus. Vet. Immunol. Immunopathol. 2012, 149, 177–185. [Google Scholar] [CrossRef]
- Sun, W.; Song, X.; Yan, R.; Xu, L.; Li, X. Vaccination of goats with a glutathione peroxidase DNA vaccine induced partial protection against Haemonchus contortus infection. Vet. Parasitol. 2011, 182, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Yan, R.; Muleke, C.I.; Sun, Y.; Xu, L.; Li, X. Vaccination of goats with DNA vaccines encoding H11 and IL-2 induces partial protection against Haemonchus contortus infection. Vet. J. 2012, 191, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.F.I.M.D.; Nobre, C.O.R.; Monteiro, J.P.; Bevilaqua, C.M.L.; Smith, D.W.; Teixeira, M. Attempt to control Haemonchus contortus in dairy goats with Barbervax ®, a vaccine derived from the nematode gut membrane glycoproteins. Small Rumin. Res. 2017, 151, 1–4. [Google Scholar] [CrossRef]
- Redmond, D.L.; Knox, D.P. Protection studies in sheep using affinity-purified and recombinant cysteine proteinases of adult Haemonchus contortus. Vaccine 2004, 22, 4252–4261. [Google Scholar] [CrossRef] [PubMed]
- Aghazadeh, Y.; Papadopoulos, V. The role of the 14-3-3 protein family in health, disease, and drug development. Drug Discov. Today 2016, 21, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Obsil, T.; Obsilova, V. Structural basis of 14-3-3 protein functions. Semin. Cell Dev. Biol. 2011, 22, 663–672. [Google Scholar] [CrossRef]
- Wang, W.; Shakes, D.C. Molecular evolution of the 14-3-3 protein family. J. Mol. Evol. 1996, 43, 384–398. [Google Scholar] [CrossRef]
- Siles-Lucas, M.; Merli, M.; Gottstein, B. 14-3-3 Proteins in Echinococcus: Their role and potential as protective antigens. Exp. Parasitol. 2008, 119, 516–523. [Google Scholar] [CrossRef]
- Schechtman, D.; Winnen, R.; Tarrabhazdai, R.; Ram, D.; Shinder, V.; Grevelding, C.G.; Kunz, W.; Arnon, R. Expression and immunolocalization of the 14-3-3 protein of Schistosoma mansoni. Parasitology 2001, 123, 573–582. [Google Scholar] [CrossRef]
- Zhao, N.; Gong, P.; Cheng, B.; Li, J.; Yang, Z.; Li, H.; Yang, J.; Zhang, G.; Zhang, X. Eimeria tenella: 14-3-3 protein interacts with telomerase. Parasitol. Res. 2014, 113, 3885. [Google Scholar] [CrossRef]
- Gadahi, J.A.; Ehsan, M.; Wang, S.; Zhang, Z.; Wang, Y.; Yan, R.; Song, X.; Xu, L.; Li, X. Recombinant protein of Haemonchus contortus 14-3-3 isoform 2(rHcftt-2) decreased the production of IL-4 and suppressed the proliferation of goat PBMCs In Vitro. Exp. Parasitol. 2016, 171, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Mcneilly, T.N.; Nisbet, A.J. Immune modulation by helminth parasites of ruminants: Implications for vaccine development and host immune competence. Parasite 2014, 21, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisbet, A.J.; McNeilly, T.N.; Wildblood, L.A.; Morrison, A.A.; Bartley, D.J.; Bartley, Y.; Longhi, C.; McKendrick, I.J.; Palarea-Albaladejo, J.; Matthews, J.B. Successful immunization against a parasitic nematode by vaccination with recombinant proteins. Vaccine 2013, 31, 4017–4023. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wang, S.; Zhang, H.; Yuan, C.; Yan, R.F. Galectin Hco-gal-m fromHaemonchus contortusmodulates goat monocytes and T cell function in different patterns. Parasit Vectors 2014, 7, 342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, M.; He, S.; Zhao, G.; Bai, Y.; Zhou, H.; Cong, H.; Lu, G.; Zhao, Q.; Zhu, X.Q. Evaluation of protective immune responses induced by DNA vaccines encodingToxoplasma gondiisurface antigen 1 (SAG1) and 14-3-3 protein in BALB/c mice. Parasites Vectors 2012, 5, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Taylor, M.G.; Johansen, M.V.; Bickle, Q.D. Vaccination of mice with a cocktail DNA vaccine induces a Th1-type immune response and partial protection against Schistosoma japonicum infection. Vaccine 2001, 20, 724–730. [Google Scholar] [CrossRef]
- Gadahi, J.A.; Wang, S.; Bo, G.; Ehsan, M.; Yan, R.; Song, X.; Xu, L.; Li, X. Proteomic Analysis of the Excretory and Secretory Proteins of Haemonchus contortus (HcESP) Binding to Goat PBMCs In Vivo Revealed Stage-Specific Binding Profiles. PLoS ONE 2016, 11, e0159796. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhu, W.; Huang, J.; Wang, X.; Sun, X.; Zhan, B.; Zhu, X. Partially protective immunity induced by the 14-3-3 protein from Trichinella spiralis. Vet. Parasitol. 2016, 231, 63–68. [Google Scholar] [CrossRef]
- Pourseif, M.M.; Moghaddam, G.; Daghighkia, H.; Nematollahi, A.; Omidi, Y. A novel B- and helper T-cell epitopes-based prophylactic vaccine against Echinococcus granulosus. Bioimpacts Bi 2018, 8, 39–52. [Google Scholar] [CrossRef]
- Ridi, R.E.; Tallima, H. Schistosoma mansoni ex vivo lung-stage larvae excretory–secretory antigens as vaccine candidates against schistosomiasis. Vaccine 2009, 27, 666–673. [Google Scholar] [CrossRef]
- Molina, J.M.; Hernandez, Y.I.; Martin, S.; Ferrer, O.; Rodriguez, F.; Ruiz, A. Immune response in goats vaccinated with thiol-binding proteins from Haemonchus contortus. Parasite Immunol. 2018, 40, e12569. [Google Scholar] [CrossRef] [PubMed]
- Knox, D.P.; Smith, W.D. Vaccination against gastrointestinal nematode parasites of ruminants using gut-expressed antigens. Vet. Parasitol. 2001, 100, 21–32. [Google Scholar] [CrossRef]
- Amarante, A.F.; Bricarello, P.A.; Huntley, J.F.; Mazzolin, L.P.; Gomes, J.C. Relationship of abomasal histology and parasite-specific immunoglobulin A with the resistance to Haemonchus contortus infection in three breeds of sheep. Vet. Parasitol. 2005, 128, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Lacroux, C.; Nguyen, T.H.; Andreoletti, O.; Prevot, F.; Grisez, C.; Bergeaud, J.P.; Gruner, L.; Brunel, J.C.; Francois, D.; Dorchies, P. Haemonchus contortus (Nematoda: Trichostrongylidae) infection in lambs elicits an unequivocal Th2 immune response. Vet. Res. 2006, 37, 607–622. [Google Scholar] [CrossRef] [Green Version]
- Mackinnon, K.M.; Bowdridge, S.A.; Kanevsky-Mullarky, I.; Zajac, A.M.; Notter, D.R. Gene expression profiles of hair and wool sheep reveal importance of Th2 immune mechanisms for increased resistance to. J. Anim. Sci. 2015, 93, 2074–2082. [Google Scholar] [CrossRef]
- Jacobs, J.R.; Greiner, S.P.; Bowdridge, S.A. Serum interleukin-4 (IL-4) production is associated with lower fecal egg count in parasite-resistant sheep. Vet. Parasitol. 2015, 211, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Kooyman, F.N.; Van Kooten, P.J.; Huntley, J.F.; Mackellar, A.; Cornelissen, A.W.; Schallig, H.D. Production of a monoclonal antibody specific for ovine immunoglobulin E and its application to monitor serum IgE responses to Haemonchus contortus infection. Parasitology 1997, 114, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Nehra, A.K.; Gowane, G.R.; Kuriyal, A.; Chaurasiya, A.; Kumar, R.; Bhinsara, D.B.; Parthasarathi, B.C.; Bhawana, K.; Khare, R.K.; Prasad, A. Immune response against subclinical haemonchosis in Himalayan hill goats. Vet. Parasitol. 2019, 267, 47–53. [Google Scholar] [CrossRef]
- Kajdaniuk, D.; Marek, B.; Borgiel-Marek, H.; Kos-Kudła, B. Transforming growth factor β1 (TGFβ1) in physiology and pathology. Endokrynol. Pol. 2013, 64, 384. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.Y.; Youngan, B.; Yun, D.H.; Yang, H.J.; Kong, Y. Experimental murine fascioliasis derives early immune suppression with increased levels of TGF-β and IL-4. Korean J. Parasitol. 2012, 50, 301. [Google Scholar] [CrossRef]
- Huang, L.; Appleton, J.A. Eosinophils in Helminth Infection: Defenders and Dupes. Trends Parasitol. 2016, 32, 798–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terefe, G.; Grisez, C.; Prevot, F.; Bergeaud, J.P.; Dorchies, P.; Brunel, J.C.; François, D.; Fourquaux, I.; Jacquiet, P. In Vitro pre-exposure of Haemonchus contortus L3 to blood eosinophils reduces their establishment potential in sheep. Vet. Res. 2007, 38, 647–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ministry of Agriculture, Fisheries and Food. Manual of Veterinary Laboratory Diagnostic Techniques; No. 418; Her Majesty’s Stationery Office: London, UK, 1989.






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bu, Y.; Jia, C.; Tian, X.; Aimulajiang, K.; Memon, M.A.; Yan, R.; Song, X.; Xu, L.; Li, X. Immunization of Goats with Recombinant Protein 14-3-3 Isoform 2(rHcftt-2) Induced Moderate Protection against Haemonchus contortus Challenge. Pathogens 2020, 9, 46. https://doi.org/10.3390/pathogens9010046
Bu Y, Jia C, Tian X, Aimulajiang K, Memon MA, Yan R, Song X, Xu L, Li X. Immunization of Goats with Recombinant Protein 14-3-3 Isoform 2(rHcftt-2) Induced Moderate Protection against Haemonchus contortus Challenge. Pathogens. 2020; 9(1):46. https://doi.org/10.3390/pathogens9010046
Chicago/Turabian StyleBu, Yongqian, Caiwen Jia, Xiaowei Tian, Kalibixiati Aimulajiang, Muhammad Ali Memon, Ruofeng Yan, Xiaokai Song, Lixin Xu, and Xiangrui Li. 2020. "Immunization of Goats with Recombinant Protein 14-3-3 Isoform 2(rHcftt-2) Induced Moderate Protection against Haemonchus contortus Challenge" Pathogens 9, no. 1: 46. https://doi.org/10.3390/pathogens9010046