Rotavirus Burden, Genetic Diversity and Impact of Vaccine in Children under Five in Tanzania



Abstract
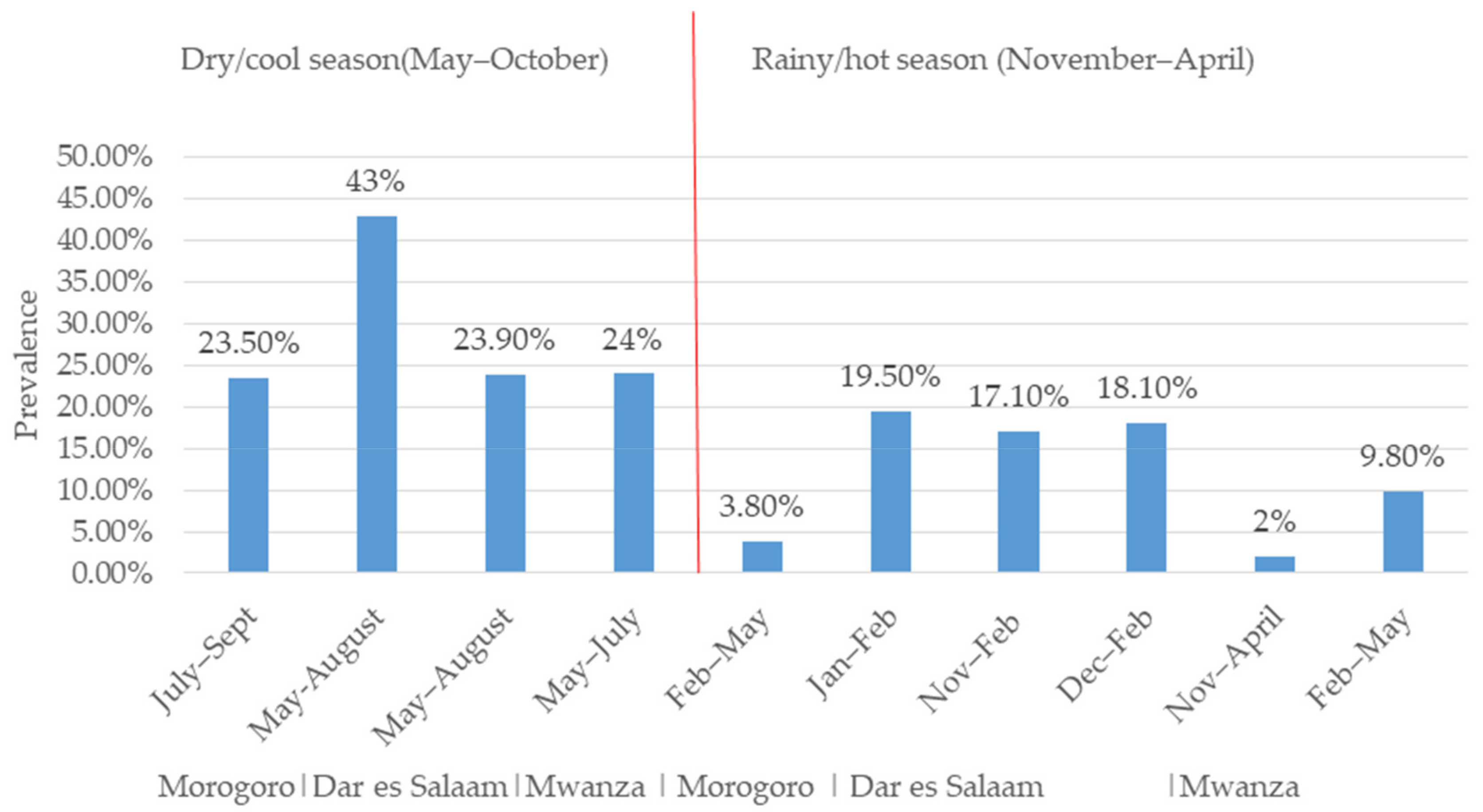
:1. Introduction
2. Trends of Rotavirus Infection and Impact of the Rotavirus Vaccine in Tanzania
3. Trends in Rotavirus Genetic Diversity in Tanzania
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Parashar, U.D.; World Health Organization–Coordinated Global Rotavirus Surveillance Network; Agocs, M.; Serhan, F.; de Oliveira, L.; Mwenda, J.M.; Mihigo, R.; et al. Global, regional, and national estimates of rotavirus mortality in children <5 years of age, 2000–2013. Clin. Infect. Dis. 2016, 62, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Troeger, C.; Forouzanfar, M.; Rao, P.C.; Khalil, I.; Brown, A.; Reiner, R.C., Jr.; Fullman, N.; Thompson, R.L.; Abajobir, A.; Ahmed, M.; et al. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: A systematic analysis for the Global Burden of Disease Study 2015. Lancet. Infect. Dis. 2017, 17, 909–948. [Google Scholar] [CrossRef]
- Fischer, T.K.; Viboud, C.; Parashar, U.; Malek, M.; Steiner, C.; Glass, R.; Simonsen, L. Hospitalizations and deaths from diarrhea and rotavirus among children <5 years of age in the United States, 1993–2003. J. Infect. Dis. 2007, 195, 1117–1125. [Google Scholar] [PubMed]
- WHO. Immunization, Vaccines and Biologicals. Rotavirus. Last Updated on December, 2018. Available online: https://www.who.int/immunization/diseases/rotavirus/en/ (accessed on 20 February 2019).
- Estes, M.K.; Kapikian, A.Z. Rotaviruses. Fields Virol. 2007, 2, 1917–1974. [Google Scholar]
- Gentsch, J.R.; Laird, A.R.; Bielfelt, B.; Griffin, D.D.; Bányai, K.; Ramachandran, M.; Jain, V.; Cunliffe, N.A.; Nakagomi, O.; Kirkwood, C.D.; et al. Serotype diversity and reassortment between human and animal rotavirus strains: Implications for rotavirus vaccine programs. J. Infect. Dis. 2005, 192, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Ciarlet, M.; Heiman, E.; Arijs, I.; Delbeke, T.; McDonald, S.M.; Palombo, E.A.; Iturriza-Gómara, M.; Maes, P.; Patton, J.T.; et al. Full genome-based classification of rotaviruses reveals a common origin between human Wa-Like and porcine rotavirus strains and human DS-1-like and bovine rotavirus strains. J. Virol. 2008, 82, 3204–3219. [Google Scholar] [CrossRef]
- Park, S.I.; Matthijnssens, J.; Saif, L.J.; Kim, H.J.; Park, J.G.; Alfajaro, M.M.; Kim, D.S.; Son, K.Y.; Yang, D.K.; Hyun, B.H.; et al. Reassortment among bovine, porcine and human rotavirus strains results in G8P [7] and G6P [7] strains isolated from cattle in South Korea. Veter Microbiol. 2011, 152, 55–66. [Google Scholar] [CrossRef]
- Arista, S.; Giammanco, G.M.; De Grazia, S.; Ramirez, S.; Biundo, C.L.; Colomba, C.; Cascio, A.; Martella, V. Heterogeneity and temporal dynamics of evolution of G1 human rotaviruses in a settled population. J. Virol. 2006, 80, 10724–10733. [Google Scholar] [CrossRef]
- Zeller, M.; Patton, J.T.; Heylen, E.; De Coster, S.; Ciarlet, M.; Van Ranst, M.; Matthijnssens, J. Genetic analyses reveal differences in the VP7 and VP4 antigenic epitopes between human rotaviruses circulating in Belgium and rotaviruses in Rotarix and RotaTeq. J. Clin. Microbiol. 2012, 50, 966–976. [Google Scholar] [CrossRef]
- Ciarlet, M.; Hoshino, Y.; Liprandi, F. Single point mutations may affect the serotype reactivity of serotype G11 porcine rotavirus strains: A widening spectrum? J. Virol. 1997, 71, 8213–8220. [Google Scholar]
- Ciarlet, M.; Reggeti, F.; Piña, C.I.; Liprandi, F. Equine rotaviruses with G14 serotype specificity circulate among Venezuelan horses. J. Clin. Microbiol. 1994, 32, 2609–2612. [Google Scholar] [PubMed]
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; Van Ranst, M.; Johne, R. VP6-sequence-based cutoff values as a criterion for rotavirus species demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Martella, V.; Van Ranst, M. Genomic evolution, host-species barrier, reassortment and classification of rotaviruses. Futur. Virol. 2010, 5, 385–390. [Google Scholar] [CrossRef]
- Kirkwood, C.D. Genetic and antigenic diversity of human rotaviruses: Potential impact on vaccination programs. J. Infect. Dis. 2010, 202, 43–48. [Google Scholar] [CrossRef] [PubMed]
- RCWG. List of Accepted Genotypes. 2018. Available online: https://rega.kuleuven.be/cev/viralmetagenomics/virus-classification/rcwg (accessed on 9 September 2018).
- Matthijnssens, J.; Bilcke, J.; Ciarlet, M.; Martella, V.; Bányai, K.; Ranst, M.V. Rotavirus disease and vaccination: Impact on genotype diversity. Futur. Microbiol. 2009, 4, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Matthijnssens, J.; Yang, X.; Delbeke, T.; Arijs, I.; Taniguchi, K.; Iturriza-Gómara, M.; Iftekharuddin, N.; Azim, T.; Van Ranst, M. Evolutionary history and global spread of the emerging G12 human rotaviruses. J. Virol. 2007, 81, 2382–2390. [Google Scholar] [CrossRef]
- Santos, N.; Hoshino, Y. Global distribution of rotavirus serotypes/genotypes and its implication for the development and implementation of an effective rotavirus vaccine. Rev. Med. Virol. 2005, 15, 29–56. [Google Scholar] [CrossRef]
- Mwenda, J.M.; Ntoto, K.M.; Abebe, A.; Enweronu-Laryea, C.; Amina, I.; Mchomvu, J.; Kisakye, A.; Mpabalwani, E.M.; Pazvakavambwa, I.; Armah, G.E.; et al. Burden and epidemiology of rotavirus diarrhea in selected African countries: Preliminary results from the African Rotavirus Surveillance Network. J. Infect. Dis. 2010, 202, 5–11. [Google Scholar] [CrossRef]
- Seheri, L.M.; Magagula, N.B.; Peenze, I.; Rakau, K.; Ndadza, A.; Mwenda, J.M.; Weldegebriel, G.; Steele, A.D.; Mphahlele, M.J. Rotavirus strain diversity in Eastern and Southern African countries before and after vaccine introduction. Vaccine 2018, 36, 7222–7230. [Google Scholar] [CrossRef]
- Seheri, M.; Nemarude, L.; Peenze, I.; Netshifhefhe, L.; Nyaga, M.M.; Ngobeni, H.G.; Maphalala, G.; Maake, L.L.; Steele, A.D.; Mwenda, J.M.; et al. Update of rotavirus strains circulating in Africa from 2007 through 2011. Pediatric Infect. Dis. J. 2014, 33, 76–84. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Ciarlet, M.; Rahman, M.; Attoui, H.; Bányai, K.; Estes, M.K.; Gentsch, J.R.; Iturriza-Gómara, M.; Kirkwood, C.D.; Martella, V.; et al. Recommendations for the classification of group A rotaviruses using all 11 genomic RNA segments. Arch. Virol. 2008, 153, 1621–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyaga, M.M.; Stucker, K.M.; Esona, M.D.; Jere, K.C.; Mwinyi, B.; Shonhai, A.; Tsolenyanu, E.; Mulindwa, A.; Chibumbya, J.N.; Adolfine, H.; et al. Whole-genome analyses of DS-1-like human G2P [4] and G8P [4] rotavirus strains from Eastern, Western and Southern Africa. Virus Genes 2014, 49, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Block, S.L.; Vesikari, T.; Goveia, M.G.; Rivers, S.B.; Adeyi, B.A.; Dallas, M.J.; Bauder, J.; Boslego, J.W.; Heaton, P.M. Efficacy, immunogenicity, and safety of a pentavalent human-bovine (WC3) reassortant rotavirus vaccine at the end of shelf life. Pediatrics 2007, 119, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Vesikari, T.; Matson, D.O.; Dennehy, P.; Van Damme, P.; Santosham, M.; Rodriguez, Z.; Dallas, M.J.; Heyse, J.F.; Goveia, M.G.; Black, S.B.; et al. Safety and efficacy of a pentavalent human–bovine (WC3) reassortant rotavirus vaccine. N. Engl. J. Med. 2006, 354, 23–33. [Google Scholar] [CrossRef]
- Afrad, M.H.; Matthijnssens, J.; Moni, S.; Kabir, F.; Ashrafi, A.; Rahman, M.Z.; Faruque, A.S.; Azim, T.; Rahman, M. Genetic characterization of a rare bovine-like human VP4 mono-reassortant G6P [8] rotavirus strain detected from an infant in Bangladesh. Infect. Genet. Evol. 2013, 19, 120–126. [Google Scholar] [CrossRef]
- Liu, L.; Johnson, H.L.; Cousens, S.; Perin, J.; Scott, S.; Lawn, J.E.; Rudan, I.; Campbell, H.; Cibulskis, R.; Li, M.; et al. Global, regional, and national causes of child mortality: An updated systematic analysis for 2010 with time trends since 2000. Lancet 2012, 379, 2151–2161. [Google Scholar] [CrossRef]
- WHO. Global Health Observatory Data Repository, Rotavirus Immunization Coverage Estimates by Country. 2018. Available online: http://apps.who.int/gho/data/node.main.ROTACnfiltertable|resettable (accessed on 2 November 2018).
- Shah, M.P.; Tate, J.E.; Mwenda, J.M.; Steele, A.D.; Parashar, U.D. Estimated reductions in hospitalizations and deaths from childhood diarrhea following implementation of rotavirus vaccination in Africa. Exp. Rev. Vaccines 2017, 16, 987–995. [Google Scholar] [CrossRef]
- Moyo, S.J.; Blomberg, B.; Hanevik, K.; Kommedal, O.; Vainio, K.; Maselle, S.Y.; Langeland, N. Genetic diversity of circulating rotavirus strains in Tanzania prior to the introduction of vaccination. PLoS ONE 2014, 9, 97562. [Google Scholar] [CrossRef]
- WHO. United Republic of Tanzania: WHO and UNICEF Estimates of Immunization Coverage: 2017 Revision. 2017. Available online: http://apps.who.int/immunization_monitoring/globalsummary/wucoveragecountrylist.html (accessed on 13 July 2019).
- Abeid, K.A.; Jani, B.; Cortese, M.M.; Kamugisha, C.; Mwenda, J.M.; Pandu, A.S.; Msaada, K.A.; Mohamed, A.S.; Khamis, A.U.; Parashar, U.D.; et al. Monovalent rotavirus vaccine effectiveness and impact on rotavirus hospitalizations in Zanzibar, Tanzania: Data from the first 3 years after introduction. J. Infect. Dis. 2016, 215, 183–191. [Google Scholar] [CrossRef]
- Brookfield, D.S.K.; Cosgrove, B.P.; Bell, E.J.; Madeley, C.R. Viruses demonstrated in children in Tanzania: Studies in diarrhoea and measles. J. Infect. 1979, 1, 249–255. [Google Scholar] [CrossRef]
- Gachanja, E.; Buza, J.; Petrucka, P. Molecular Detection of Group A Rotavirus in Children under Five in Urban and Peri-Urban Arusha, Tanzania. J. Adv. Med. Med Res. 2016, 12, 1–9. [Google Scholar] [CrossRef]
- Hokororo, A.; Kidenya, B.R.; Seni, J.; Mapaseka, S.; Mphahlele, J.; Mshana, S.E. 2Predominance of rotavirus G1 [P8] genotype among under-five children with gastroenteritis in Mwanza, Tanzania. J. Trop. Pediatric 2014, 60, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Jani, B.; Hokororo, A.; Mchomvu, J.; Cortese, M.M.; Kamugisha, C.; Mujuni, D.; Kallovya, D.; Parashar, U.D.; Mwenda, J.M.; Lyimo, D.; et al. Detection of rotavirus before and after monovalent rotavirus vaccine introduction and vaccine effectiveness among children in mainland Tanzania. Vaccine 2018, 36, 7149–7156. [Google Scholar] [CrossRef] [PubMed]
- Mchaile, D.N.; Philemon, R.N.; Kabika, S.; Albogast, E.; Morijo, K.J.; Kifaro, E.; Mmbaga, B.T. Prevalence and genotypes of Rotavirus among children under 5 years presenting with diarrhoea in Moshi, Tanzania: A hospital-based cross sectional study. BMC Res. Notes 2017, 10, 542. [Google Scholar] [CrossRef]
- Mhalu, F.S.; Myrmel, H.; Msengi, A.; Haukenes, G. Prevalence of infection with rotavirus and enteric adenoviruses among children in Tanzania. NIPH Ann. 1988, 11, 3–7. [Google Scholar]
- Moyo, S.J.; Gro, N.; Kirsti, V.; Matee, M.I.; Kitundu, J.; Maselle, S.Y.; Langeland, N.; Myrmel, H. Prevalence of enteropathogenic viruses and molecular characterization of group A rotavirus among children with diarrhea in Dar es Salaam Tanzania. BMC Public Health 2007, 7, 359. [Google Scholar] [CrossRef]
- Sam, N.E.; Haukenes, G.; Szilvay, A.M.; Mhalu, F. Rotavirus infection in Tanzania: A virological, epidemiological and clinical study among young children. APMIS 1992, 100, 790–796. [Google Scholar] [CrossRef]
- Temu, A.; Kamugisha, E.; Mwizamholya, D.L.; Hokororo, A.; Seni, J.; Mshana, S.E. Prevalence and factors associated with Group A rotavirus infection among children with acute diarrhea in Mwanza, Tanzania. J. Infect. Dev. Ctries. 2012, 6, 508–515. [Google Scholar] [CrossRef]
- Temu, M.M.; Changalucha, J.M.; Mngara, J.T.; Steele, A.D. Prevalence of rotavirus infections and strain types detected among under-five children presenting with diarrhoea at selected MCH clinics in Mwanza City, Tanzania. Tanzan. J. Health Res. 2002, 4, 30–32. [Google Scholar] [CrossRef]
- Vargas, M.; Gascon, J.; Casals, C.; Schellenberg, D.; Urassa, H.; Kahigwa, E.; Ruiz, J.; Vila, J. Etiology of diarrhea in children less than five years of age in Ifakara, Tanzania. Am. J. Trop. Med. Hyg. 2004, 70, 536–539. [Google Scholar] [CrossRef]
- PATH. Rotavirus Disease and Vaccines in Tanzania. 2012. Available online: https://www.path.org/resources/rotavirus-disease-and-vaccines-in-tanzania/ (accessed on 22 September 2018).
- WHO. 2008 Rotavirus Deaths, under 5 Years of Age, as of 31 January 2012. Available online: https://www.who.int/immunization/monitoring_surveillance/burden/estimates/rotavirus/en (accessed on 12 November 2018).
- Platts-Mills, J.A.; Amour, C.; Gratz, J.; Nshama, R.; Walongo, T.; Mujaga, B.; Maro, A.; McMurry, T.L.; Liu, J.; Mduma, E.; et al. Impact of rotavirus vaccine introduction and postintroduction etiology of diarrhea requiring hospital admission in Haydom, Tanzania, a rural African setting. Clin. Infect. Dis. 2017, 65, 1144–1151. [Google Scholar] [CrossRef] [PubMed]
- Bar-Zeev, N.; Jere, K.C.; Bennett, A.; Pollock, L.; Tate, J.E.; Nakagomi, O.; Iturriza-Gomara, M.; Costello, A.; Mwansambo, C.; Parashar, U.D.; et al. Population impact and effectiveness of monovalent rotavirus vaccination in urban Malawian children 3 years after vaccine introduction: Ecological and case-control analyses. Clin. Infect. Dis. 2016, 62, S213–S219. [Google Scholar] [CrossRef] [PubMed]
- Bar-Zeev, N.; Kapanda, L.; Tate, J.E.; Jere, K.C.; Iturriza-Gomara, M.; Nakagomi, O.; Mwansambo, C.; Costello, A.; Parashar, U.D.; Heyderman, R.S.; et al. Effectiveness of a monovalent rotavirus vaccine in infants in Malawi after programmatic roll-out: An observational and case-control study. Lancet Infect. Dis. 2015, 15, 422–428. [Google Scholar] [CrossRef]
- Ngabo, F.; Tate, J.E.; Gatera, M.; Rugambwa, C.; Donnen, P.; Lepage, P.; Mwenda, J.M.; Binagwaho, A.; Parashar, U.D. Effect of pentavalent rotavirus vaccine introduction on hospital admissions for diarrhoea and rotavirus in children in Rwanda: A time-series analysis. Lancet Glob. Health 2016, 4, 129–136. [Google Scholar] [CrossRef]
- Levy, K.; Hubbard, A.E.; Eisenberg, J.N. Seasonality of rotavirus disease in the tropics: A systematic review and meta-analysis. Int. J. Epidemiol. 2009, 38, 1487–1496. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.E.; Panozzo, C.A.; Payne, D.C.; Patel, M.M.; Cortese, M.M.; Fowlkes, A.L.; Parashar, U.D. Decline and Change in Seasonality of US Rotavirus Activity After the Introduction of Rotavirus Vaccine. Am. Acad. Pediatrics 2009, 123, 465–471. [Google Scholar] [CrossRef]
- National Bureau of Statistics. 2015–16 Tanzania Demographic and Health Survey and Malaria Indicator Survey (TDHS-MIS) Report. Available online: https://www.nbs.go.tz/index.php/en/census-surveys/health-statistics/demographic-and-health-survey-dhs (accessed on 21 July 2019).
- Beres, L.K.; Tate, J.E.; Njobvu, L.; Chibwe, B.; Rudd, C.; Guffey, M.B.; Stringer, J.S.; Parashar, U.D.; Chilengi, R.A. preliminary assessment of rotavirus vaccine effectiveness in Zambia. Clin. Infect. Dis. 2016, 62, 175–182. [Google Scholar] [CrossRef]
- Gastañaduy, P.A.; Steenhoff, A.P.; Mokomane, M.; Esona, M.D.; Bowen, M.D.; Jibril, H.; Pernica, J.M.; Mazhani, L.; Smieja, M.; Tate, J.E.; et al. Effectiveness of monovalent rotavirus vaccine after programmatic implementation in Botswana: A multisite prospective case-control study. Clin. Infect. Dis. 2016, 62, 161–167. [Google Scholar] [CrossRef]
- Groome, M.J.; Page, N.; Cortese, M.M.; Moyes, J.; Zar, H.J.; Kapongo, C.N.; Mulligan, C.; Diedericks, R.; Cohen, C.; Fleming, J.A.; et al. Effectiveness of monovalent human rotavirus vaccine against admission to hospital for acute rotavirus diarrhoea in South African children: A case-control study. Lancet Infect. Dis. 2014, 14, 1096–1104. [Google Scholar] [CrossRef]
- Bányai, K.; Gentsch, J.R.; Schipp, R.; Jakab, F.; Meleg, E.; Mihály, I.; Szücs, G. Dominating prevalence of P [8], G1 and P [8], G9 rotavirus strains among children admitted to hospital between 2000 and 2003 in Budapest, Hungary. J. Med. Virol. 2005, 76, 414–423. [Google Scholar] [CrossRef]
- Sharma, S.; Paul, V.K.; Bhan, M.K.; Ray, P. Genomic characterization of nontypeable rotaviruses and detection of a rare G8 strain in Delhi, India. J. Clin. Microbiol. 2009, 47, 3998–4005. [Google Scholar] [CrossRef] [PubMed]
- Cortese, M.M.; Parashar, U.D. Prevention of rotavirus gastroenteritis among infants and children: Recommendations of the Advisory Committee on Immunization Practices (ACIP). Morb. Mortal. Wkly. Rep. Recomm. Rep. 2009, 58, 1–25. [Google Scholar]
- Steele, A.D.; Neuzil, K.M.; Cunliffe, N.A.; Madhi, S.A.; Bos, P.; Ngwira, B.; Witte, D.; Todd, S.; Louw, C.; Kirsten, M.; et al. Human rotavirus vaccine Rotarix™ provides protection against diverse circulating rotavirus strains in African infants: A randomized controlled trial. BMC Infect. Dis. 2012, 12, 213. [Google Scholar] [CrossRef] [PubMed]
- De Grazia, S.; Ramirez, S.; Giammanco, G.M.; Colomba, C.; Martella, V.; Biundo, C.L.; Mazzola, R.; Arista, S. Diversity of human rotaviruses detected in Sicily, Italy, over a 5-year period (2001–2005). Arch. Virol. 2007, 152, 833–837. [Google Scholar] [CrossRef]
- Martella, V.; Bányai, K.; Matthijnssens, J.; Buonavoglia, C.; Ciarlet, M. Zoonotic aspects of rotaviruses. Vet. Microbiol. 2010, 140, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Matthijnssens, J.; Goegebuer, T.; De Leener, K.; Vanderwegen, L.; van der Donck, I.; Van Hoovels, L.; De Vos, S.; Azim, T.; Van Ranst, M. Predominance of rotavirus G9 genotype in children hospitalized for rotavirus gastroenteritis in Belgium during 1999–2003. J. Clin. Virol. 2005, 33, 1–6. [Google Scholar] [CrossRef]
- Ward, R.L.; Bernstein, D.I.; Plotkin, S. Rotarix: A rotavirus vaccine for the world. Clin. Infect. Dis. 2009, 48, 222–228. [Google Scholar] [CrossRef]
- Banyai, K.; Mijatovic-Rustempasic, S.; Hull, J.J.; Esona, M.D.; Freeman, M.M.; Frace, A.M.; Bowen, M.D.; Gentsch, J.R. Sequencing and phylogenetic analysis of the coding region of six common rotavirus strains: Evidence for intragenogroup reassortment among co-circulating G1P [8] and G2P [4] strains from the United States. J. Med. Virol. 2011, 83, 532–539. [Google Scholar] [CrossRef]
- Martella, V.; Bányai, K.; Ciarlet, M.; Iturriza-Gómara, M.; Lorusso, E.; De Grazia, S.; Arista, S.; Decaro, N.; Elia, G.; Cavalli, A.; et al. Relationships among porcine and human P [6] rotaviruses: Evidence that the different human P [6] lineages have originated from multiple interspecies transmission events. Virology 2006, 344, 509–519. [Google Scholar] [CrossRef]
- Papp, H.; László, B.; Jakab, F.; Ganesh, B.; De Grazia, S.; Matthijnssens, J.; Ciarlet, M.; Martella, V.; Bányai, K. Review of group A rotavirus strains reported in swine and cattle. Vet. Microbiol. 2013, 165, 190–199. [Google Scholar] [CrossRef]
- Assenga, J.A.; Matemba, L.E.; Muller, S.K.; Mhamphi, G.G.; Kazwala, R.R. Predominant leptospiral serogroups circulating among humans, livestock and wildlife in Katavi-Rukwa ecosystem, Tanzania. PLoS Negl. Trop. Dis. 2015, 9, e0003607. [Google Scholar] [CrossRef] [PubMed]
- Bishop, R.F. Natural history of human rotavirus infection. Arch. Virol. Suppl. 1996, 12, 119–128. [Google Scholar] [PubMed]
- Ansari, S.A.; Springthorpe, V.S.; Sattar, S.A. Survival and vehicular spread of human rotaviruses: Possible relation to seasonality of outbreaks. Rev. Infect. Dis. 1991, 13, 448–461. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; De Grazia, S.; Piessens, J.; Heylen, E.; Zeller, M.; Giammanco, G.M.; Bányai, K.; Buonavoglia, C.; Ciarlet, M.; Martella, V.; et al. Multiple reassortment and interspecies transmission events contribute to the diversity of feline, canine and feline/canine-like human group A rotavirus strains. Infect. Genet. Evol. 2011, 11, 1396–1406. [Google Scholar] [CrossRef]
- Komoto, S.; Adah, M.I.; Ide, T.; Yoshikawa, T.; Taniguchi, K. Whole genomic analysis of human and bovine G8P [1] rotavirus strains isolated in Nigeria provides evidence for direct bovine-to-human interspecies transmission. Infect. Genet. Evol. 2016, 43, 424–433. [Google Scholar] [CrossRef]
- Medici, M.C.; Tummolo, F.; Bonica, M.B.; Heylen, E.; Zeller, M.; Calderaro, A.; Matthijnssens, J. Genetic diversity in three bovine-like human G8P [14] and G10P [14] rotaviruses suggests independent interspecies transmission events. J. Gen. Virol. 2015, 96, 1161–1168. [Google Scholar] [CrossRef]
- Tacharoenmuang, R.; Komoto, S.; Guntapong, R.; Ide, T.; Singchai, P.; Upachai, S.; Fukuda, S.; Yoshida, Y.; Murata, T.; Yoshikawa, T.; et al. Characterization of a G10P [14] rotavirus strain from a diarrheic child in Thailand: Evidence for bovine-to-human zoonotic transmission. Infect. Genet. Evol. 2018, 63, 43–57. [Google Scholar] [CrossRef]
- Dóró, R.; Farkas, S.L.; Martella, V.; Bányai, K. Zoonotic transmission of rotavirus: Surveillance and control. Expert Rev. Anti-Infect. Ther. 2015, 13, 1337–1350. [Google Scholar] [CrossRef]
- Medici, M.C.; Abelli, L.A.; Vito, M.; Monica, M.; Eleonora, L.; Canio, B.; Giuseppe, D.; Carlo, C. Characterization of inter-genogroup reassortant rotavirus strains detected in hospitalized children in Italy. J. Med. Virol. 2007, 79, 1406–1412. [Google Scholar] [CrossRef]
- Iturriza-Gómara, M.; Cubitt, D.; Desselberger, U.; Gray, J. Amino acid substitution within the VP7 protein of G2 rotavirus strains associated with failure to serotype. J. Clin. Microbiol. 2001, 39, 3796–3798. [Google Scholar] [CrossRef]
- Iturriza-Gómara, M.; Kang, G.; Gray, J. Rotavirus genotyping: Keeping up with an evolving population of human rotaviruses. J. Clin. Virol. 2004, 31, 259–265. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Region | Reference | Diagnostic Technique | Settings, Year of Study | Rotavirus Vaccine Dosage Comparison | VE (95% CI) |
|---|---|---|---|---|---|
| Zanzibar | [1] | EIA | Single hospital, December 2012–2015 | 2 doses vs. 0 doses | 57 (14–78) |
| Manyara | [47] | EIA | Single hospital, August–December 2015 | 2 doses vs. 0 doses 2 doses vs. 0 doses | 74.8 (−8.2 to 94.1) 85.1 (26.5–97.0) |
| Mwanza | [28] | RT-PCR | Multiple hospitals, May 2015 | 2 doses vs. 0 doses ≥1 dose vs. 0 doses | 49 (−30 to 80) 53 (−14 to 81) |
| Region | Ref | Year | n | G1P [8] | G2P [4] | G9P [8] | G12P [6] | G12P [8] | G1P [4] | G8P [4] | G8P [6] | G4P [6] | G4P [4] | G3P [6] | G3P [8] | G8P [8] | G1P [6] | Untypeable | Mixed genotypes |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dar es Salaam | [48] | 2005–2006 | 49 | 10% | 0 | 90.7% | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5% | 10% | 0 |
| [47] | 2010–2011 | 211 | 64.7% | 0 | 0.5% | 11.1% | 0 | 3.7% | 14.2% | 3.2% | 1.1% | 1.1% | 0 | 0 | 0 | 0.5% | 0 | 0 | |
| Tanga | [49] | 2007–2008 | 32 | 34% | 0 | 3% | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3% | 28% | 31% |
| Mwanza | [25] | 2010–2011. | 100 | 24% | 2% | 0 | 0 | 0 | 0 | 7% | 4% | 0 | 0 | 0 | 0 | 6% | 17% | 0 | 29% |
| Zanzibar | [1] | 2010–2015 | 101 | 52% | 8% | 0 | 0 | 2% | 0 | 0 | 3% | 0 | 0 | 16% | 0 | 0 | 1% | 7% | 12% |
| ND | [28] | 2009–2015 | 32 | 15.63% | 50% | 0 | 0 | 0 | 3.1% | 0 | 0 | 0 | 0 | 15.6% | 3.1% | 0 | 3.1% | 6.3% | 9.3% |
| ND | [62] | 2010 | – | 31.4% | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| ND | [62] | 2011 | – | 25.8% | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| ND | [62] | 2014 | – | 41.1% | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| ND | [62] | 2015 | – | 75.1% | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| East Africa | [63] | 2007–2011 | – | 23% | 8% | 12% | 0 | 4% | 0 | 4% | 5% | 0 | 0 | 4% | 0 | 0 | 4% | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malakalinga, J.J.; Misinzo, G.; Msalya, G.M.; Kazwala, R.R. Rotavirus Burden, Genetic Diversity and Impact of Vaccine in Children under Five in Tanzania. Pathogens 2019, 8, 210. https://doi.org/10.3390/pathogens8040210
Malakalinga JJ, Misinzo G, Msalya GM, Kazwala RR. Rotavirus Burden, Genetic Diversity and Impact of Vaccine in Children under Five in Tanzania. Pathogens. 2019; 8(4):210. https://doi.org/10.3390/pathogens8040210
Chicago/Turabian StyleMalakalinga, Joseph J., Gerald Misinzo, George M. Msalya, and Rudovick R. Kazwala. 2019. "Rotavirus Burden, Genetic Diversity and Impact of Vaccine in Children under Five in Tanzania" Pathogens 8, no. 4: 210. https://doi.org/10.3390/pathogens8040210