
Characterization of a Novel Species of Legionella Isolated from a Healthcare Facility: Legionella resiliens sp. nov
 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Isolation of Bacterial Strains and Growth Conditions
2.2. Identification by Serological Test and MALDI–TOF MS Technique
2.3. Physiology and Chemotaxonomy
2.4. DNA Extraction and Gene Sequencing
2.5. Phylogenetic Analyses Based on Gene Sequences
2.6. Whole Genome Sequencing and Genome Features
2.7. Core Genome
- Total genes (0% ≤ strains ≤ 100%);
- Core genes (99% ≤ strains ≤ 100%);
- Soft core genes 95% ≤ strains < 99%);
- Shell genes (15% ≤ strains < 95%);
- Cloud genes (0% ≤ strains < 15%).
2.8. Plasmid, Virulence, Pathogenicity, and Antibiotic Resistance Gene Analysis
2.9. Antibiotic Susceptibility Test
3. Results and Discussion
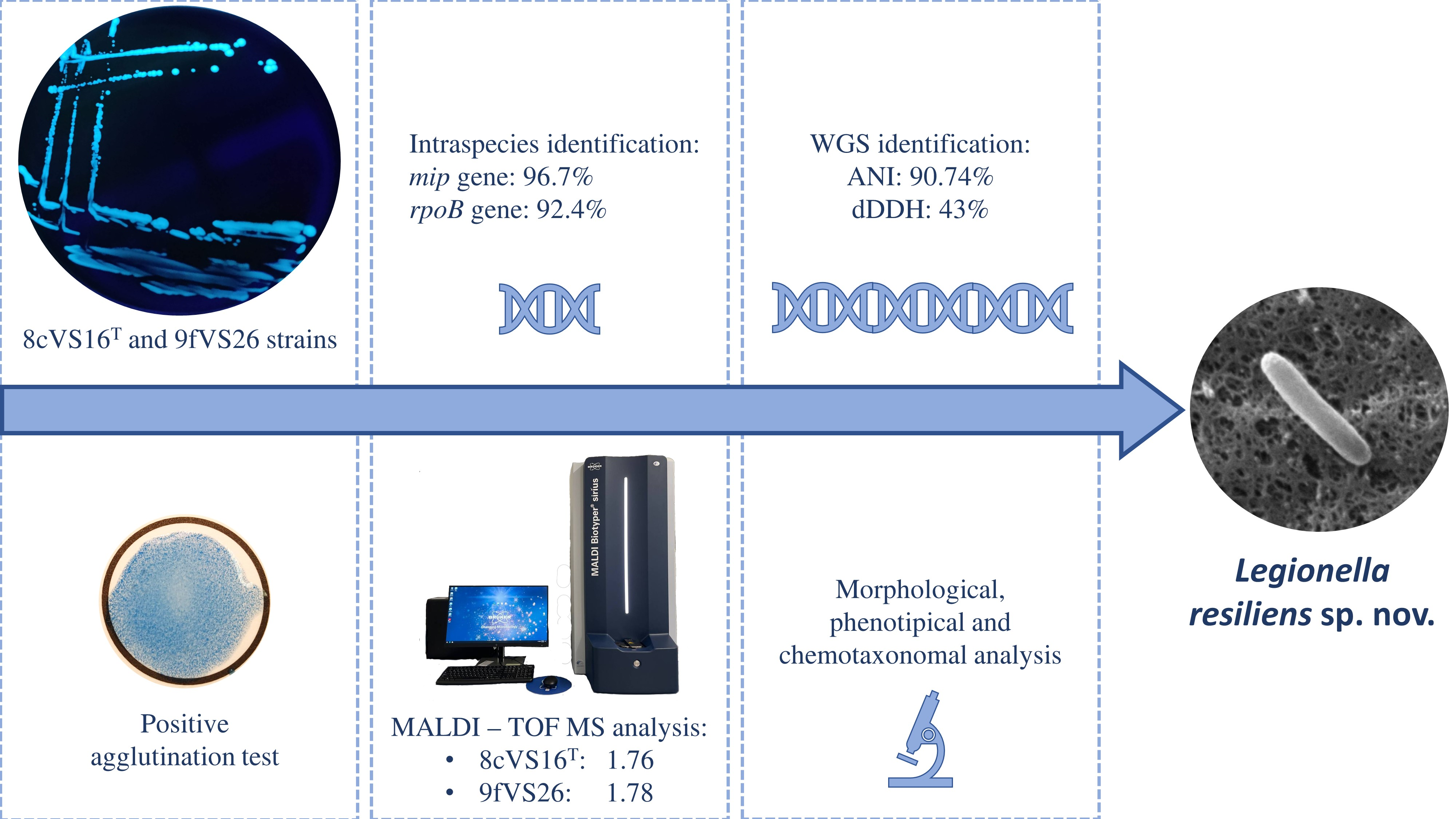
3.1. Isolation of Bacterial Strains, Growth Conditions, and Identification
3.2. Physiological, Biochemical, and Morphological Features
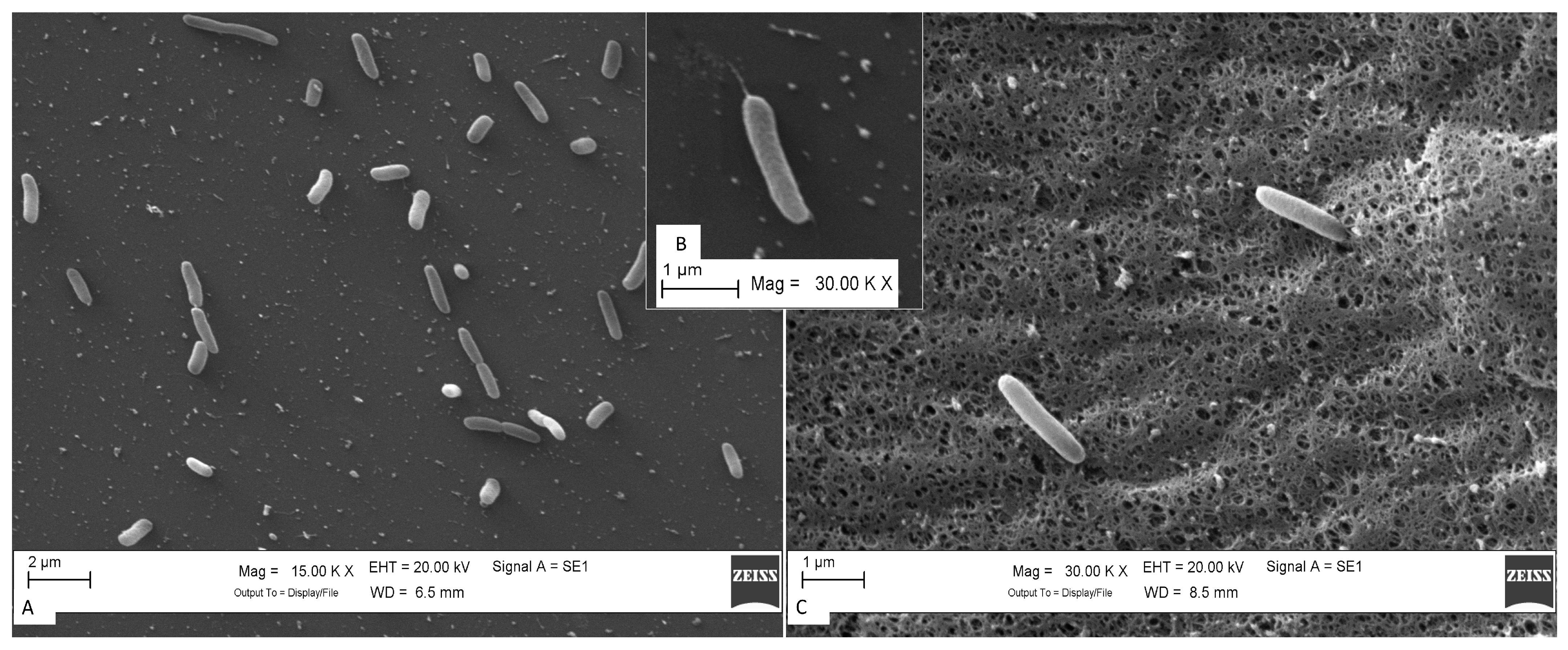
Scanning Electron Microscopy (SEM) and Motility Gene Patterns
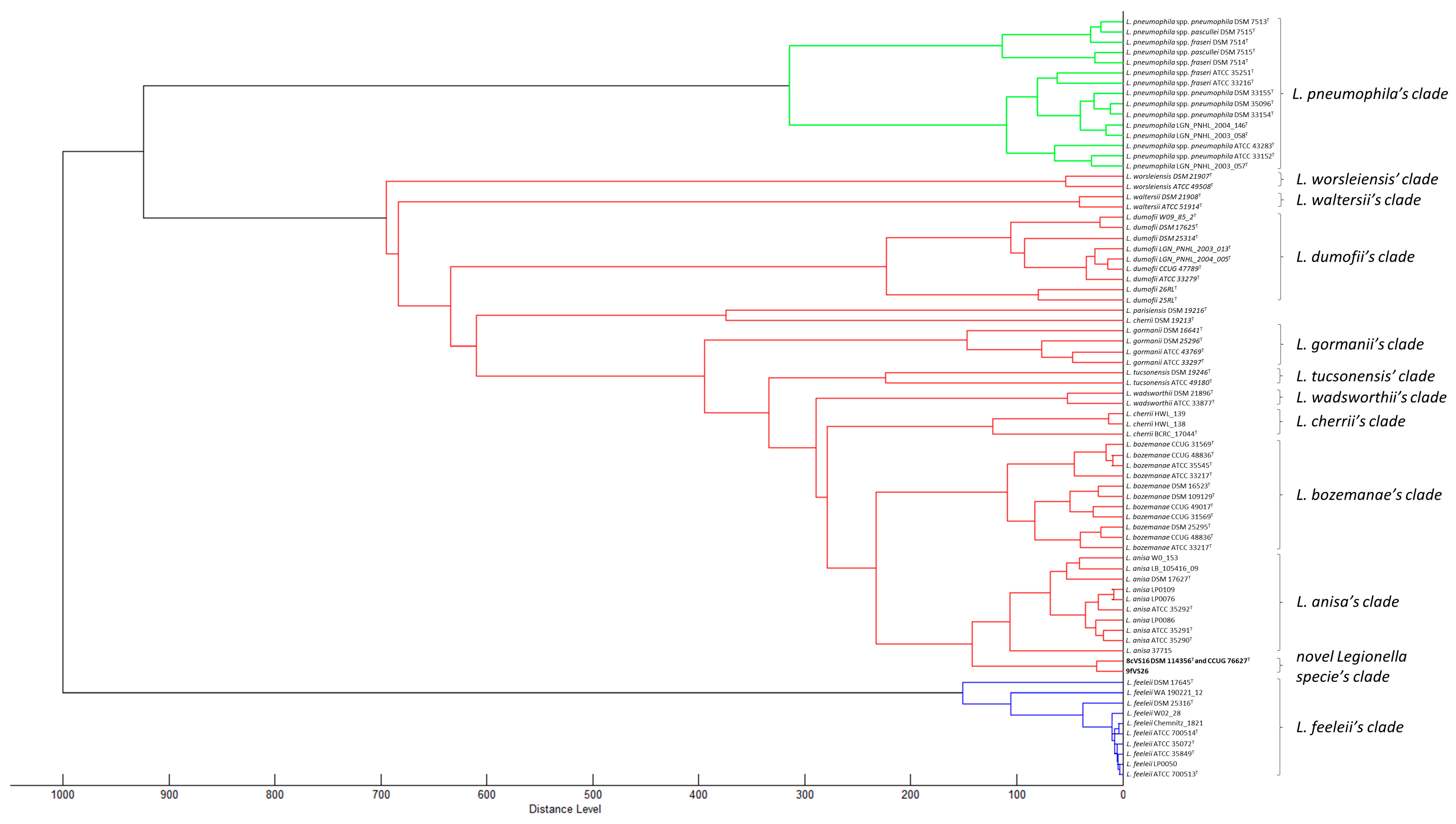
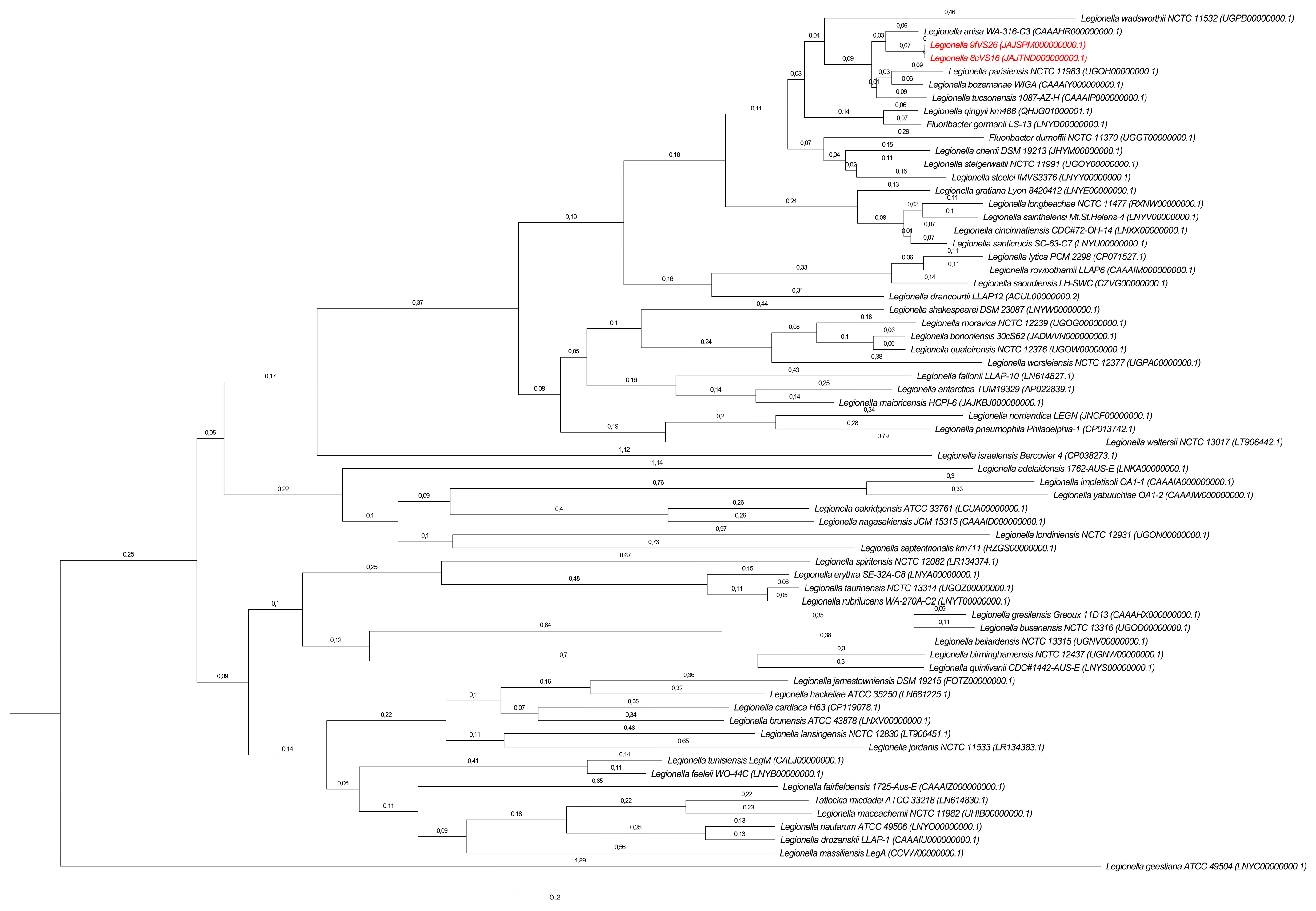
3.3. Phylogenetic and Sequencing Analyses
- Regarding 16SrRNA, using 1537 bp obtained by WGS, the range of similarity with respect to the top ten Legionella species was between 97.34 and 99.29%;
- Considering the classification scheme targeting the mip sequence for the identification of novel Legionella isolates, using 611 bp, the range was 88.93–96.73%;
- In relation to the new classification scheme targeting the rpoB sequencing for a deep-resolution identification of the novel Legionella isolate, using 329 bp, the range was between 83.07 and 92.40%. In addition, using the entire rpoB gene (4107 bp) obtained by WGS, the range of similarity was 80.32–95.13%.
- mip: MW052957.1 and MW052913.1;
- rpoB: MZ367138 and MZ367095;
- 16S rRNAs: OL804581.1 and OL889882.1.
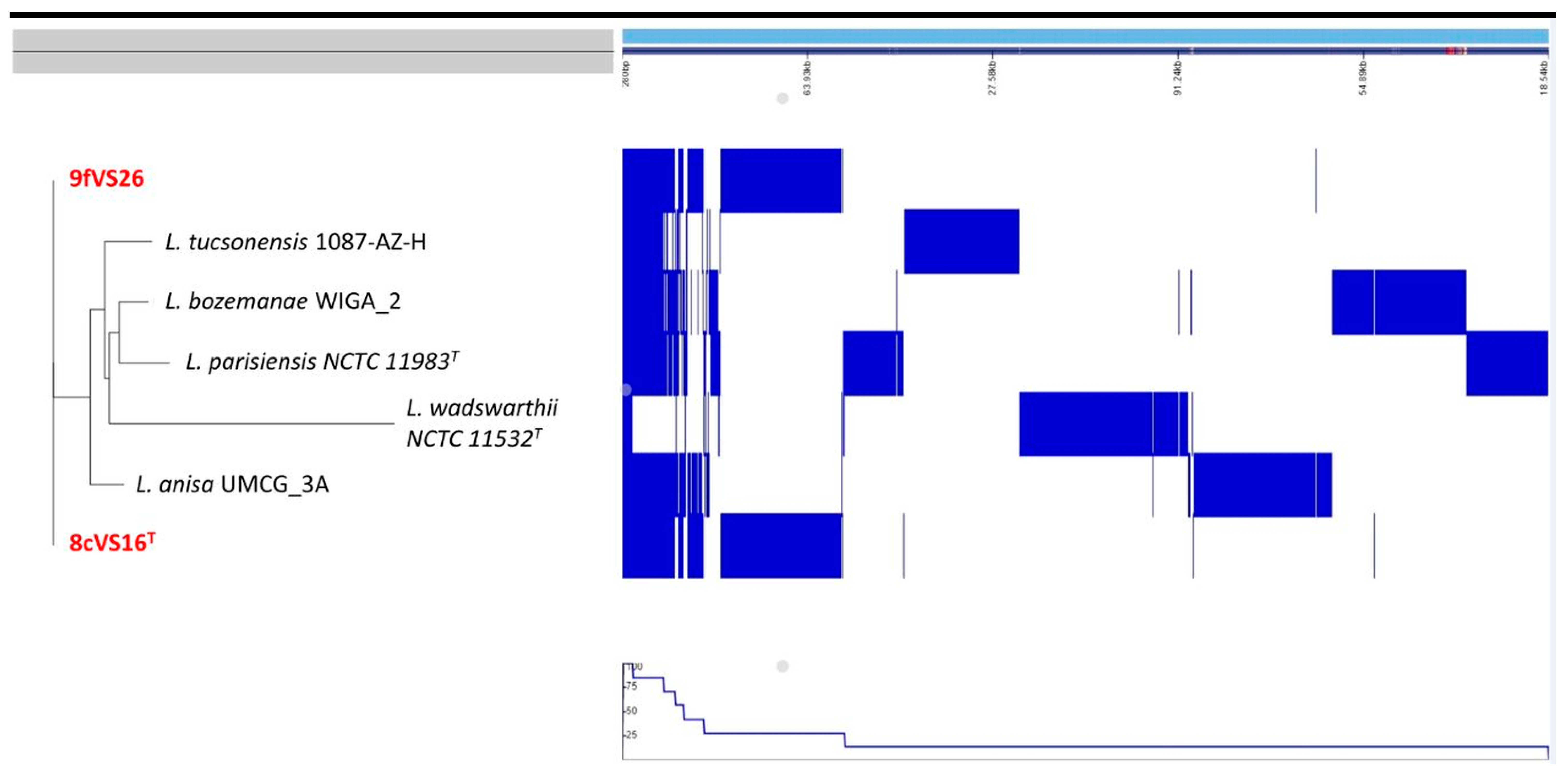
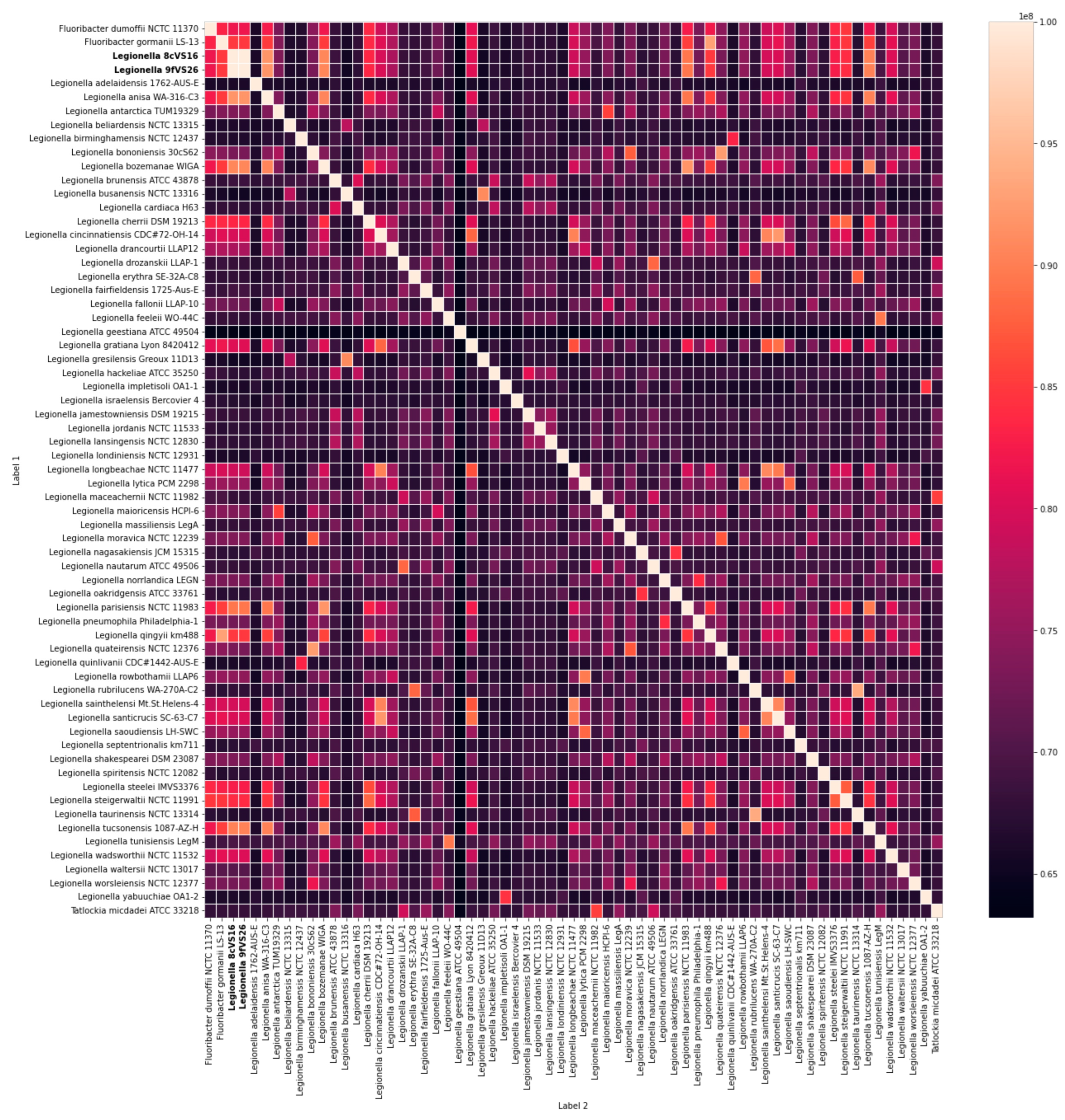
Whole Genome Sequencing (WGS) and Comparative Analysis
3.4. Core Genome
- Total genes: 174,828;
- Core genes: 3;
- Soft core genes: 7;
- Shell genes: 484;
- Cloud genes: 174,334.
3.5. Plasmid, Virulence, Pathogenicity, and Antibiotic Resistance Gene Results
3.6. Antibiotic Susceptibility Test Results
4. Conclusions
Description of Legionella resiliens sp. nov.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fields, B.S.; Benson, R.F.; Besser, R.E. Legionella and Legionnaires’ Disease: 25 Years of Investigation. J. Clin. Microbiol. 2002, 15, 506–526. [Google Scholar] [CrossRef] [PubMed]
- Harrison, T.G.; Saunders, N.A. Taxonomy and Typing of Legionellae. Med. Microbiol. 1994, 5, 79–90. [Google Scholar] [CrossRef]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) Moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Dupuy, M.; Mazoua, S.; Berne, F.; Bodet, C.; Garrec, N.; Herbelin, P.; Ménard-Szczebara, F.; Oberti, S.; Rodier, M.H.; Soreau, S.; et al. Efficiency of Water Disinfectants against Legionella pneumophila and Acanthamoeba. Water Res. 2011, 45, 1087–1094. [Google Scholar] [CrossRef] [PubMed]
- Amemura-Maekawa, J.; Hayakawa, Y.; Sugie, H.; Moribayashi, A.; Kura, F.; Chang, B.; Wada, A.; Watanabe, H. Legioliulin, a New Isocoumarin Compound Responsible for Blue-White Autofluorescence in Legionella (Fluoribacter) dumoffii under Long-Wavelength UV Light. Biochem. Biophys. Res. Commun. 2004, 323, 954–959. [Google Scholar] [CrossRef]
- Ko, K.S.; Lee, H.K.; Park, M.Y.; Lee, K.H.; Yun, Y.J.; Woo, S.Y.; Miyamoto, H.; Kook, Y.H. Application of RNA Polymerase β-Subunit Gene (RpoB) Sequences for the Molecular Differentiation of Legionella Species. J. Clin. Microbiol. 2002, 40, 2653–2658. [Google Scholar] [CrossRef]
- Ratcliff, R.M.; Lanser, J.A.; Manning, P.A.; Heuzenroeder, M.W. Sequence-Based Classification Scheme for the Genus Legionella Targeting the Mip Gene. J. Clin. Microbiol. 1998, 36, 1560–1567. [Google Scholar] [CrossRef]
- Birtles, R.J.; Rowbotham, T.J.; Raoult, D.; Harrison, T.G. Phylogenetic Diversity of Intra-Amoebal Legionellae as Revealed by 16S RRNA Gene Sequence Comparison. Microbiology 1996, 142, 3525–3530. [Google Scholar] [CrossRef]
- Diogo, A.; Verissimo, A.; Nobre, M.F.; Da Costa, M.S. Usefulness of Fatty Acid. Composition for Differentiation of Legionella Species. J. Clin. Microbiol. 1999, 37, 2248–2254. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Legionella (Legionnaires’ Disease and Pontiac Fever)—Causes, How It Spreads, and People at Increased Risk. Available online: https://www.cdc.gov/legionella/about/causes-transmission.html (accessed on 6 July 2023).
- Rota, M.C.; Caporali, M.G.; Bella, A.; Scaturro, M.; Giannitelli, S.; Ricci, M.L. I Risultati Del Sistema Di Sorveglianza Della Legionellosi Nel 2021. Boll. Epidemiol. Naz. 2022, 2, 9–16. [Google Scholar] [CrossRef]
- Herwaldt, L.A.; Marra, A.R. Legionella: A Reemerging Pathogen. Curr. Opin. Infect. Dis. 2018, 31, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Newton, H.J.; Ang, D.K.Y.; Van Driel, I.R.; Hartland, E.L. Molecular Pathogenesis of Infections Caused by Legionella pneumophila. Clin. Microbiol. Rev. 2010, 23, 274–298. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control (ECDC). Legionnaires’ Disease. In Annual Epidemiological Report for 2021; ECDC: Stockholm, Sweden, 2023. [Google Scholar]
- David, S.; Rusniok, C.; Mentasti, M.; Gomez-Valero, L.; Harris, S.R.; Lechat, P.; Lees, J.; Ginevra, C.; Glaser, P.; Ma, L.; et al. Multiple Major Disease-Associated Clones of Legionella pneumophila Have Emerged Recently and Independently. Genome Res. 2016, 26, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Chambers, S.T.; Slow, S.; Scott-thomas, A.; Murdoch, D.R. Legionellosis Caused by Non-Legionella pneumophila Species, with a Focus on Legionella longbeachae. Microorganisms 2021, 9, 291. [Google Scholar] [CrossRef] [PubMed]
- Muder, R.R.; Yu, V.L. Infection Due to Legionella Species Other than L. Pneumophila. Clin. Infect. Dis. 2002, 35, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Dedicoat, M.; Venkatesan, P. The Treatment of Legionnaires’ Disease. J. Antimicrob. Chemother. 1999, 43, 747–752. [Google Scholar] [CrossRef]
- Mazzotta, M.; Salaris, S.; Pascale, M.R.; Girolamini, L.; Cristino, S. Occurrence of Legionella Spp. In Man-Made Water Sources: Isolates Distribution and Phylogenetic Characterization in the Emilia-Romagna Region. Pathogens 2021, 10, 552. [Google Scholar] [CrossRef]
- Emilia-Romagna Region. Regional Guidelines for Surveillance and Control of Legionellosis. Delibera Della Giunta Regionale 12 Giugno 2017, N. 828. 2017. 2017, 2012, 1–136. Available online: https://www.salute.gov.it/imgs/C_17_pubblicazioni_2362_allegato.pdf (accessed on 6 July 2023).
- Girolamini, L.; Pascale, M.R.; Mazzotta, M.; Spiteri, S.; Marino, F.; Salaris, S.; Grottola, A.; Orsini, M.; Cristino, S. Combining Traditional and Molecular Techniques Supports the Discovery of a Novel Legionella Species during Environmental Surveillance in a Healthcare Facility. Front. Microbiol. 2022, 13, 900936. [Google Scholar] [CrossRef]
- Italian National Institute of Health. Guidelines for Prevention and Control of Legionellosis. Approvate in Conferenza Stato-Regioni Seduta Del 7 Maggio 2015. Italy. 2015. Available online: https://bur.regione.emilia-romagna.it/bur/area-bollettini/bollettini-in-lavorazione/n-167-del-19-06-2017-parte-seconda.2017-06-19.6161668613/approvazione-delle-linee-guida-regionali-per-la-sorveglianza-e-il-controllo-della-legionellosi/allegato-1-linee-guida-regiona.2017-06-19.1497864629 (accessed on 6 July 2023).
- UNI EN ISO 19458:2006; Water Quality—Sampling for Microbiological Analysis. ISO: Geneva, Switzerland, 2006.
- UNI EN ISO 11731:2017; Water Quality—Enumeration of Legionella. ISO: Geneva, Switzerland, 2017.
- Pascale, M.R.; Mazzotta, M.; Salaris, S.; Girolamini, L.; Grottola, A.; Simone, M.L.; Cordovana, M.; Bisognin, F.; Dal Monte, P.; Bucci Sabattini, M.A.; et al. Evaluation of MALDI–TOF Mass Spectrometry in Diagnostic and Environmental Surveillance of Legionella Species: A Comparison With Culture and Mip-Gene Sequencing Technique. Front. Microbiol. 2020, 11, 589369. [Google Scholar] [CrossRef]
- Crespi, S.; Drašar, V.; Salvà-Serra, F.; Jaén-Luchoro, D.; Piñeiro-Iglesias, B.; Lindemann, P.C.; Aliaga-Lozano, F.; Fernández-Juárez, V.; Coll-García, G.; Moore, E.R.B.; et al. Legionella maioricensis Sp. Nov., a New Species Isolated from the Hot Water Distribution Systems of a Hospital and a Shopping Center during Routine Sampling. Int. J. Syst. Evol. Microbiol. 2023, 73, 00568. [Google Scholar] [CrossRef]
- Lambert, M.A.; Moss, C.W. Cellular Fatty Acid Compositions and Isoprenoid Quinone Contents of 23 Legionella Species. J. Clin. Microbiol. 1989, 27, 465–473. [Google Scholar] [CrossRef]
- Thacker, W.L.; Benson, R.F.; Schifman, R.B.; Pugh, E.; Steigerwalt, A.G.; Mayberry, W.R.; Brenner, D.J.; Wilkinson, H.W. Legionella tucsonensis Sp. Nov. Isolated from a Renal Transplant Recipient. J. Clin. Microbiol. 1989, 27, 1831–1834. [Google Scholar] [CrossRef]
- Yong, S.F.Y.; Tan, S.H.; Wee, J.; Tee, J.J.; Sansom, F.M.; Newton, H.J.; Hartland, E.L. Molecular Detection of Legionella: Moving on from Mip. Front. Microbiol. 2010, 1, 123. [Google Scholar] [CrossRef]
- Cianciotto, N.P.; Kwaik, Y.; Edelstein, P.H.; Fields, B.S.; Geary, D.; Harrison, T.G.; Joseph, C.A.; Ratcliff, R.M.; Stout, J.E. Legionella—State of the Art 30 Years after Its Recognition; Swanson, M.S., Ed.; AMS Press: Washington, DC, USA, 2006. [Google Scholar]
- Fry, N.K.; Afshar, B.; Bellamy, W.; Underwood, A.P.; Ratcliff, R.M.; Harrison, T.G.; Bangsborg, J.; Blanco, S.; Etienne, J.; Fendukly, F.; et al. Identification of Legionella Spp. by 19 European Reference Laboratories: Results of the European Working Group for Legionella Infections External Quality Assessment Scheme Using DNA Sequencing of the Macrophage Infectivity Potentiator Gene and Dedicated Online Tools. Clin. Microbiol. Infect. 2007, 13, 1119–1124. [Google Scholar] [CrossRef]
- Pascale, M.R.; Salaris, S.; Mazzotta, M.; Girolamini, L.; Fregni Serpini, G.; Manni, L.; Grottola, A.; Cristino, S. New Insight Regarding Legionella Non-Pneumophila Species Identification: Comparison between the Traditional Mip Gene Classification Scheme and a Newly Proposed Scheme Targeting the RpoB Gene. Microbiol. Spectr. 2021, 9, e0116121. [Google Scholar] [CrossRef]
- Kim, B.J.; Lee, S.H.; Lyu, M.A.; Kim, S.J.; Bai, G.H.; Kim, S.J.; Chae, G.T.; Kim, E.C.; Cha, C.Y.; Kook, Y.H. Identification of Mycobacterial Species by Comparative Sequence Analysis of the RNA Polymerase Gene (RpoB). Int. J. Lepr. Other Mycobact. Dis. 1999, 67, 514. [Google Scholar] [CrossRef]
- Rafiee, M.; Jahangiri-Rad, M.; Hajjaran, H.; Mesdaghinia, A.; Hajaghazadeh, M. Detection and Identification of Legionella Species in Hospital Water Supplies through Polymerase Chain Reaction (16S RRNA). J. Environ. Health Sci. Eng. 2014, 12, 83. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16s RRNA Sequence Analysis in the Present. Species Definition in Bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Ebers, J. Taxonomic Parameters Revisited: Tarnished Gold Standards. Microbiol. Today 2006, 33, 152–155. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian Evolutionary Analysis by Sampling Trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Quijada, N.M.; Rodríguez-Lázaro, D.; Eiros, J.M.; Hernández, M. TORMES: An Automated Pipeline for Whole Bacterial Genome Analysis. Bioinformatics 2019, 35, 4207–4212. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Chen, K.T.; Liu, C.L.; Huang, S.H.; Shen, H.T.; Shieh, Y.K.; Chiu, H.T.; Lu, C.L. CSAR: A Contig Scaffolding Tool Using Algebraic Rearrangements. Bioinformatics 2018, 34, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness. Methods Mol. Biol. 2019, 1962, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; Dicuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI Prokaryotic Genome Annotation Pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.-M.; Lim, J.; Kwon, S.; Chun, J. A Large-Scale Evaluation of Algorithms to Calculate Average Nucleotide Identity. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High Throughput ANI Analysis of 90K Prokaryotic Genomes Reveals Clear Species Boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef]
- Tanizawa, Y.; Fujisawa, T.; Nakamura, Y. DFAST: A Flexible Prokaryotic Genome Annotation Pipeline for Faster Genome Publication. Bioinformatics 2018, 34, 1037–1039. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A Web Server for Prokaryotic Species Circumscription Based on Pairwise Genome Comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A Database Tandem for Fast and Reliable Genome-Based Classification and Nomenclature of Prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Olson, R.D.; Assaf, R.; Brettin, T.; Conrad, N.; Cucinell, C.; Davis, J.J.; Dempsey, D.M.; Dickerman, A.; Dietrich, E.M.; Kenyon, R.W.; et al. Introducing the Bacterial and Viral Bioinformatics Resource Center (BV-BRC): A Resource Combining PATRIC, IRD and ViPR. Nucleic Acids Res. 2023, 51, D678–D689. [Google Scholar] [CrossRef]
- Cock, P.J.A.; Antao, T.; Chang, J.T.; Chapman, B.A.; Cox, C.J.; Dalke, A.; Friedberg, I.; Hamelryck, T.; Kauff, F.; Wilczynski, B.; et al. Biopython: Freely Available Python Tools for Computational Molecular Biology and Bioinformatics. Bioinformatics 2009, 25, 1422–1423. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A Rapid Bootstrap Algorithm for the RAxML Web Servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Georghiou, R.; Doggett, A.M.; Kielhofner, M.A.; Stout, J.E.; Watson, D.A.; Lupski, J.R.; Hamill, R.J. Molecular Fingerprinting of Legionella Species by Repetitive Element PCR. J. Clin. Microbiol. 1994, 32, 2989–2994. [Google Scholar] [CrossRef] [PubMed]
- Michelini, S.; Modesto, M.; Pisi, A.M.; Filippini, G.; Sandri, C.; Spiezio, C.; Biavati, B.; Sgorbati, B.; Mattarelli, P. Bifidobacterium eulemuris Sp. Nov., Isolated from Faeces of Black Lemurs (Eulemur Macaco). Int. J. Syst. Evol. Microbiol. 2016, 66, 1567–1576. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple Prokaryote Genome Comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed]
- Sitto, F.; Battistuzzi, F.U. Estimating Pangenomes with Roary. Mol. Biol. Evol. 2020, 37, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid Large-Scale Prokaryote Pan Genome Analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Park, S.; Chun, J. Introducing EzAAI: A Pipeline for High Throughput Calculations of Prokaryotic Average Amino Acid Identity. J. Microbiol. 2021, 59, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Moreira, B.; Vinuesa, P. GET_HOMOLOGUES, a Versatile Software Package for Scalable and Robust Microbial Pangenome Analysis. Appl. Environ. Microbiol. 2013, 79, 7696–7701. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, D.M.; Kannan, L.; Coleman, M.K.; Wolf, Y.I.; Sorokin, A.; Koonin, E.V.; Mushegian, A. A Low-Polynomial Algorithm for Assembling Clusters of Orthologous Groups from Intergenomic Symmetric Best Matches. Bioinformatics 2010, 26, 1481–1487. [Google Scholar] [CrossRef]
- Li, L.; Stoeckert, C.J.; Roos, D.S. OrthoMCL: Identification of Ortholog Groups for Eukaryotic Genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef]
- Schmartz, G.P.; Hartung, A.; Hirsch, P.; Kern, F.; Fehlmann, T.; Müller, R.; Keller, A. PLSDB: Advancing a Comprehensive Database of Bacterial Plasmids. Nucleic Acids Res. 2022, 50, D273–D278. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations Using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A General Classification Scheme for Bacterial Virulence Factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-Annot, a New Bioinformatic Tool to Discover Antibiotic Resistance Genes in Bacterial Genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2019, 48, D517–D525. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Cosentino, S.; Voldby Larsen, M.; Møller Aarestrup, F.; Lund, O. PathogenFinder—Distinguishing Friend from Foe Using Bacterial Whole Genome Sequence Data. PLoS ONE 2013, 8, e77302. [Google Scholar] [CrossRef]
- EUCAST. Antimicrobial Susceptibility Testing of Legionella pneumophila; EUCAST: Vaxjo, Sweden, 2021; Volume 2021, Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Guidance_documents/Legionella_guidance_note_-_20210528.pdf (accessed on 6 July 2023).
- Portal, E.; Sands, K.; Portnojs, A.; Chalker, V.J.; Spiller, O.B. Legionella Antimicrobial Sensitivity Testing: Comparison of Microbroth Dilution with BCYE and LASARUS Solid Media. J. Antimicrob. Chemother. 2021, 76, 1197–1204. [Google Scholar] [CrossRef]
- Brenner, D.J.; Steigerwalt, A.G.; Gorman, G.W.; Wilkinson, H.W.; Bibb, W.F.; Hackel, M.; Tyndall, R.L.; Campbell, J.; Feeley, J.C.; Lanier Thacker, W.; et al. Ten New Species of Legionella. Int. J. Syst. Evol. Microbiol. 1985, 35, 50–59. [Google Scholar] [CrossRef]
- Edelstein, P.H.; Brenner, J.; Moss, C.W.; Steigerwalt, A.G.; Francis, E.M.; Lance George, W. Legionella wadsworthii Species Nova: A Cause of Human Pneumonia. Ann. Intern. Med. 1982, 97, 809–881. [Google Scholar] [CrossRef]
- Gorman, G.W.; Feeley, J.C.; Steigerwalt, A.; Edelstein, P.H.; Wayne Moss, C.; Brenner, D.J. Legionella anisa: A New Species of Legionella Isolated from Potable Waters and a Cooling Tower. Appl. Environ. Microbiol. 1985, 49, 305–309. [Google Scholar] [CrossRef]
- Journal, A.I.; Brenner, D.J.; Steigerwalt, A.G.; Gorman, G.W.; Weaver, R.E.; James, C.F.; Feeley, C.; Cot-Des, L.G.; Wilkinson, H.W.; Patton, C.; et al. Legionella bozemanii Sp. Nov. and Legionella dumoffii Sp. Nov.: Classification of Two Additional Species of Legionella Associated with Human Pneumonia. Curr. Microbiol. 1980, 4, 111–116. [Google Scholar]
- Appelt, S.; Heuner, K. The Flagellar Regulon of Legionella—A Review. Front. Cell Infect. Microbiol. 2017, 7, 454. [Google Scholar] [CrossRef]
- Fang, Z.; Li, L.; Zhou, J.; You, A.; Gao, L.; Li, T.; Chen, H.; Han, R.; Cui, Y.; Chen, L.; et al. Multiple Nucleotide Polymorphism DNA Markers for the Accurate Evaluation of Genetic Variations. bioRxiv 2021. [Google Scholar] [CrossRef]
- Mercuri, P.S.; García-Sáez, I.; De Vriendt, K.; Thamm, I.; Devreese, B.; Van Beeumen, J.; Dideberg, O.; Rossolini, G.M.; Frère, J.M.; Galleni, M. Probing the Specificity of the Subclass B3 FEZ-1 Metallo-β-Lactamase by Site-Directed Mutagenesis. J. Biol. Chem. 2004, 279, 33630–33638. [Google Scholar] [CrossRef] [PubMed]
- Svetlicic, E.; Jaén-Luchoro, D.; Klobucar, R.S.; Jers, C.; Kazazic, S.; Franjevic, D.; Klobucar, G.; Shelton, B.G.; Mijakovic, I. Genomic Characterization and Assessment of Pathogenic Potential of Legionella Spp. Isolates from Environmental Monitoring. Front. Microbiol. 2023, 13, 1091964. [Google Scholar] [CrossRef]
- Cocuzza, C.E.; Martinelli, M.; Perdoni, F.; Giubbi, C.; Vinetti, M.E.A.; Calaresu, E.; Frugoni, S.; Scaturro, M.; Ricci, M.L.; Musumeci, R. Antibiotic Susceptibility of Environmental Legionella pneumophila Strains Isolated in Northern Italy. Int. J. Environ. Res. Public Health 2021, 18, 9352. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | 8cVS16T and 9fVS26 | L. anisa | L. bozemanae | L. parisiensis | L. tucsonensis | L. wadsworthii | Lp 1 |
|---|---|---|---|---|---|---|---|
| Accession number | DSM 114356 | ATCC 35292 | ATCC33217 | ATCC 35299 | ATCC 40180 | ATCC 33877 | ATCC 33152 |
| Catalase | + | + | + | + | + | + | + |
| Urease | − | − | − | − | − | − | − |
| Hippurate hydrolysis | − | − | − | − | − | − | + |
| Oxidase | + | + | − | + | − | − | + |
| β-Lactamase production | + | + | +/− | + | + | + | + |
| Gelatin liquefaction (gelatinase) | + | + | + | + | + | + | + |
| Glucose fermentation | − | − | − | − | − | − | − |
| Autofluorescence | + | + | + | + | + | + | − |
| Substrates | 8cVS16T and 9fVS26 | L. anisa | Lp1 | Substrates | 8cVS16T and 9fVS26 | L. anisa | Lp1 |
|---|---|---|---|---|---|---|---|
| ARA (arabinose) | − | − | − | GLU (glucose) | − | − | − |
| MNS (mannose) | − | − | − | PRO (proline-β-naphthylamide) | − | + | − |
| SUC (sucrose) | − | − | − | PYR (pyrrolidine-b-naphthylamide) | − | − | − |
| MEL (melibiose) | − | − | − | GGT (g-Glutamyl b-naphthylamide) | − | + | − |
| RHA (rhamnose) | − | − | − | TRY (tryptophane b-naphthylamide) | − | − | + |
| SOR (sorbitol) | − | − | − | IND (tryptophane) | − | − | − |
| MNT (mannitol) | − | − | − | NO3 (sodium nitrate) | + | + | + |
| ADO (adonitol) | − | − | − | GLR (p-nitrophenly β-glucuronide) | − | − | − |
| ONPG (ρ-Nitrophenyl, b,D-galactoside) | − | − | − | NAG (ρ-Nitrophenyl-N-acetyl-β,D-glucosaminide) | − | − | − |
| PHO (p-nitrophenyl phosphate) | + | + | + | GGL (γ-L-glutamyl p-nitroanilide) | − | − | − |
| BGL (p-nitrophenyl α-β-glucoside) | − | − | − | ESC (esculin) | − | − | − |
| NPG (p-nitrophenyl β-galactoside) | − | − | − | PHE (p-nitro-DL-phenylalanine) | − | − | − |
| BPH (p-nitrophenyl bis-phosphate) | − | − | − | URE (urea) | − | − | + |
| BXY (p-nitrophenyl xyloside) | − | − | − | CIT (citrate) | − | − | − |
| AAR (p-nitrophenyl α-arabinoside) | − | − | − | MLO (malonate) | − | − | − |
| PHC (p-nitrophenyl phosphorylcholine) | − | − | − | TTC (tetrazolium) | − | − | − |
| ADH/ARG (arginine) | − | − | − | LYS (lysine) | − | − | − |
| TRD (aliphatic thiol) | + | + | + | GLY (glycine) | − | − | + |
| PHS (ρ-Nitrophenyl-phosphoester) | + | + | + | BANA (N-Benzyl-arginine-b-naphthylamide) | − | − | − |
| αGLU (ρ-Nitrophenyl-α,D-glucoside) | − | − | − | EST (triglyceride) | + | + | + |
| βGLU (ρ-Nitrophenyl-β,D-glucoside) | − | − | − | INO (inositol) | − | − | − |
| GAL (galactose) | − | − | − |
| 16S rRNA Gene (1537 bp) (Accession Number) | Identity Percentage | Coverage | Partial mip Gene (611 bp) (Accession Number) | Identity Percentage | Coverage | rpoB Gene (4107 bp) (Accession Number) | Identity Percentage | Coverage |
|---|---|---|---|---|---|---|---|---|
| L. anisa DSM 17627T (CP082852.1) | 99.29% | 100% | L. anisa DSM 17627T (CP082852.1) | 96.73% | 100% | L. anisa DSM 17627T (CP082852.1) | 95.13% | 100% |
| L. cherrii NCTC 11976T (LR134173.1) | 98.51% | 100% | L. tucsonensis ATCC 49180T (U92224.1) | 94.95% | 100% | L. cherrii NCTC 11976T (LR134173.1) | 87.60% | 100% |
| L. sainthelensi NCTC 11988T (LR134388.1) | 98.25% | 100% | L. bozemanae ATCC 33217T (U91609.1) | 94.30% | 100% | L. oakridgensis NCTC11531T (LR134286.1) | 83.94 | 100% |
| L. oakrigensis NCTC 11531T (LR134286.1) | 98.12% | 100% | L. parisiensis ATCC 35299T (GU083754.1) | 93.97% | 100% | L. longbeachae DSM 10572T (CP082850.1) | 83.94% | 100% |
| L. longbeachae DSM 10572T (CP082850.1) | 98.12% | 100% | L. steigerwaltii ATCC 35302T (U92223.1) | 91.53% | 100% | L. sainthelensi NCTC11988T (LR134388.1) | 83.63% | 100% |
| L. qingyii KCTC 15636T (NR_171519.1) | 98.80% | 97% | L. cherrii NCTC 11976T (LR134173.1) | 91.35% | 99% | L. lytica PCM 2298T (CP071527.1) | 83.02% | 100% |
| L. pneumophila ATCC 33152T (CP040987.1) | 97.92% | 100% | L. gormanii ATCC 33297T (U91638.1) | 90.88% | 100% | L. antarctica NCTC 14581T (AP022839.1) | 81.22% | 100% |
| L. fallonii ATCC 700992T (LN614827.1) | 97.53% | 100% | L. steelei IMVS 3376T (HQ398203.1) | 90.55% | 100% | L. fallonii DSM 19889T (LN614827.1) | 80.91% | 100% |
| L. parisiensis JCM 7561T (LC504039.1) | 99.05% | 95% | L. wadsworthii ATCC 33877T (U92225.1) | 88.93% | 100% | L. pneumophila subsp. Pascullei NCTC12273T (LR134380.1) | 80.39% | 100% |
| L. waltersii NTCT 13017T (LT906442.1) | 97.34% | 100% | L. qingyii KCTC 15636T (MH189580.1) | 91.16% | 90% | L. pneumophila ATCC33152T (CP040987.1) | 80.32% | 99% |
| Attribute | Data for Strain | |
|---|---|---|
| 8cVS16T | 9fVS26 | |
| No. of raw reads | 1,787,078 | 1,952,986 |
| Avg read length (bp) | 256 | 259 |
| Coverage (×) | 115 | 127 |
| Total Length (bp) | 3,906,083 | 3,906,100 |
| No. of contigs | 7 | 10 |
| GC Content (mol%) | 38.2 | 38.2 |
| N50 (bp) | 855,940 | 858,038 |
| No. of coding sequences | 3362 | 3360 |
| No. of rRNAs | 6 | 6 |
| No. of tRNAs | 42 | 42 |
| Type Strains | Taxon Name | GenBank Accession ID | No. of Contigs | Size (Mbp) | GC (mol%) | No. of CDS | No. of rRNA | No. of tRNA |
|---|---|---|---|---|---|---|---|---|
| 8cVS16T | 8cVS16T | GCA_021344005.1 | 7 | 3.9 | 38.2 | 3368 | 9 | 42 |
| 9fVS26 | 9fVS26 | GCA_021282285.1 | 10 | 3.91 | 38.2 | 3371 | 9 | 42 |
| WA-316-C3 | L. anisa | GCA_900639785.1 | 178 | 4.4 | 38.17 | 3.869 | 3 | 43 |
| WIGA | L. bozemanae | GCA_900640135.1 | 98 | 4.13 | 37.91 | 3.665 | 4 | 43 |
| NCTC11983 | L. parisiensis | GCA_900461585.1 | 2 | 4.2 | 38.0 | 3.663 | 9 | 44 |
| 1087-AZ-H | L. tusconensis | GCA_900640035.1 | 27 | 3.36 | 37.41 | 2.948 | 3 | 42 |
| NCTC11532 | L. wadswarthii | GCA_900452925.1 | 2 | 3.6 | 38.08 | 3.147 | 11 | 43 |
| C9_S | L. pneumophila | GCA_001753085.1 | 3 | 3.5 | 38.0 | 3084 | 9 | 43 |
| Antimicrobial | Concentration Range Tested (mg/L) | MIC for 8cVS16T | Interpretation | MIC for 9fVS26 | Interpretation | MIC for Lp1 ATCC 33152T | Interpretation | Lp EUCAST Cut-Off (Not Standardized) |
|---|---|---|---|---|---|---|---|---|
| Azithromycin | 0.016–256 | 1 | R | 1 | R | 0.125 | S | 0.25 |
| Ciprofloxacin | 0.002–32 | 0.25 | S | 0.25 | S | 0.75 | S | 0.5 |
| Doxicicline | 0.016–256 | 3 | S | 3 | S | N.D. | / | 8 |
| Erythromycin | 0.016–256 | 0.5 | S | 0.5 | S | 0.125 | S | 0.5 |
| Levofloxacine | 0.002–32 | 0.125 | S | 0.125 | S | N.D. | / | 0.25 |
| Rifampicin | 0.002–32 | 0.064 | S | 0.064 | S | 0.023 | S | 0.032 |
| Antimicrobial | Concentration Range Tested (mg/L) | MIC for 8cVS16T | Interpretation | MIC for 9fVS26 | Interpretation | MIC for Lp1 ATCC 33152T | Interpretation | Lp EUCAST Cut-Off (Not Standardized) |
|---|---|---|---|---|---|---|---|---|
| Azithromycin | 0.0075–16 | 1 | R | 1 | R | 0.5 | R | 0.125 |
| Erythromycin | 0.0075–16 | 0.5 | S | 0.5 | S | 1 | S | 1 |
| Ciprofloxacin | 0.00025–0.512 | 0.016 | S | 0.016 | S | 0.032 | S | 0.032 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cristino, S.; Pascale, M.R.; Marino, F.; Derelitto, C.; Salaris, S.; Orsini, M.; Squarzoni, S.; Grottola, A.; Girolamini, L. Characterization of a Novel Species of Legionella Isolated from a Healthcare Facility: Legionella resiliens sp. nov. Pathogens 2024, 13, 250. https://doi.org/10.3390/pathogens13030250
Cristino S, Pascale MR, Marino F, Derelitto C, Salaris S, Orsini M, Squarzoni S, Grottola A, Girolamini L. Characterization of a Novel Species of Legionella Isolated from a Healthcare Facility: Legionella resiliens sp. nov. Pathogens. 2024; 13(3):250. https://doi.org/10.3390/pathogens13030250
Chicago/Turabian StyleCristino, Sandra, Maria Rosaria Pascale, Federica Marino, Carlo Derelitto, Silvano Salaris, Massimiliano Orsini, Stefano Squarzoni, Antonella Grottola, and Luna Girolamini. 2024. "Characterization of a Novel Species of Legionella Isolated from a Healthcare Facility: Legionella resiliens sp. nov" Pathogens 13, no. 3: 250. https://doi.org/10.3390/pathogens13030250