
The NLRP3 Inflammasome Is Dispensable in Methicillin-Resistant Staphylococcus aureus Urinary Tract Infection

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria, Mice, and the Reagents
2.2. Mouse Models of Ascending UTI and Catheter-Associated UTI (CAUTI)
2.3. MCC950 Treatment of the Mouse Model of Ascending UTI
2.4. Cytokine Profiling by ELISA
2.5. Immune Cell Infiltration in the Bladder and the Kidney Tissues
2.6. Histopathological Examination of the Bladder and Kidney
2.7. Statistical Analysis
3. Results
3.1. The Pathophysiology of MRSA UTI in WT and Nlrp3−/− Mice
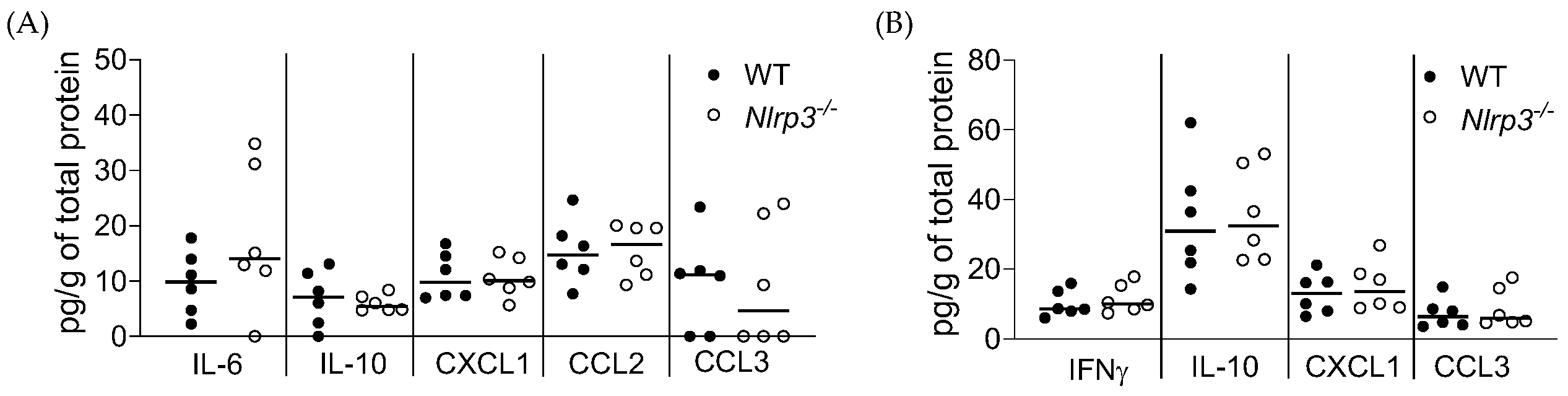
3.2. Acute MRSA 1369 UTI Induces Similar Cytokine Production and Immune Cell Recruitment to the UT of WT and Nlrp3−/− Mice
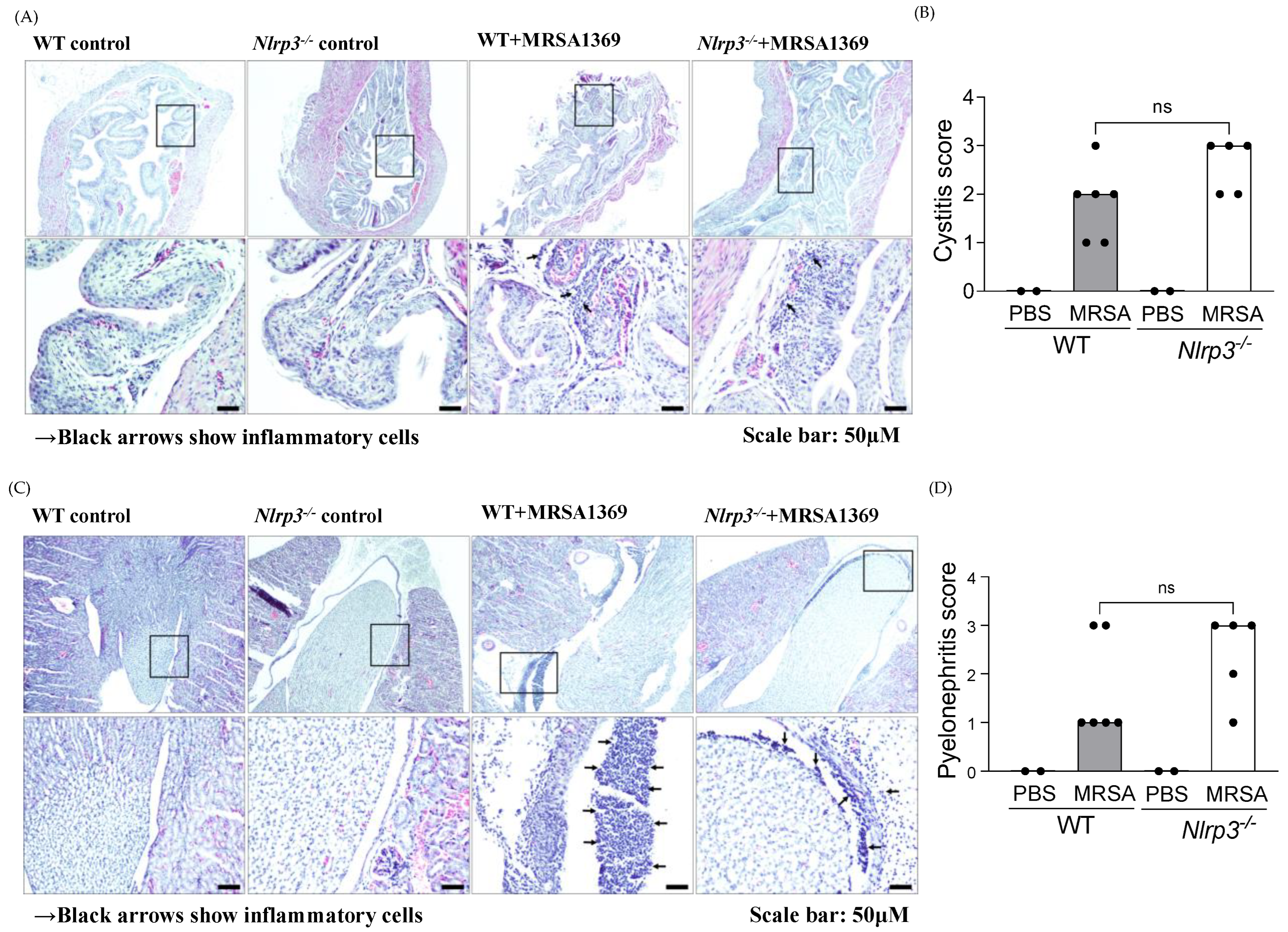
3.3. Histopathological Examination of WT and Nlrp3−/− Bladder and Kidney Sections
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wagenlehner, F.M.; Cek, M.; Naber, K.G.; Kiyota, H.; Bjerklund-Johansen, T.E. Epidemiology, treatment and prevention of healthcare-associated urinary tract infections. World J. Urol. 2012, 30, 59–67. [Google Scholar] [CrossRef]
- Gaston, J.R.; Johnson, A.O.; Bair, K.L.; White, A.N.; Armbruster, C.E. Polymicrobial interactions in the urinary tract: Is the enemy of my enemy my friend? Infect. Immun. 2021, 89, e00652-20. [Google Scholar] [CrossRef]
- Gajdacs, M.; Abrok, M.; Lazar, A.; Burian, K. Increasing relevance of Gram-positive cocci in urinary tract infections: A 10-year analysis of their prevalence and resistance trends. Sci. Rep. 2020, 10, 17658. [Google Scholar] [CrossRef]
- Ackermann, R.J.; Monroe, P.W. Bacteremic urinary tract infection in older people. J. Am. Geriatr. Soc. 1996, 44, 927–933. [Google Scholar] [CrossRef]
- Shrestha, L.B.; Baral, R.; Khanal, B. Comparative study of antimicrobial resistance and biofilm formation among Gram-positive uropathogens isolated from community-acquired urinary tract infections and catheter-associated urinary tract infections. Infect. Drug Resist. 2019, 12, 957–963. [Google Scholar] [CrossRef]
- Routh, J.C.; Alt, A.L.; Ashley, R.A.; Kramer, S.A.; Boyce, T.G. Increasing prevalence and associated risk factors for methicillin resistant Staphylococcus aureus bacteriuria. J. Urol. 2009, 181, 1694–1698. [Google Scholar] [CrossRef] [PubMed]
- Looney, A.T.; Redmond, E.J.; Davey, N.M.; Daly, P.J.; Troy, C.; Carey, B.F.; Cullen, I.M. Methicillin-resistant Staphylococcus aureus as a uropathogen in an Irish setting. Medicine (Baltimore) 2017, 96, e4635. [Google Scholar] [CrossRef] [PubMed]
- Shigemura, K.; Tanaka, K.; Osawa, K.; Arakawa, S.; Miyake, H.; Fujisawa, M. Clinical factors associated with shock in bacteremic UTI. Int. Urol. Nephrol. 2013, 45, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Muder, R.R.; Brennen, C.; Rihs, J.D.; Wagener, M.M.; Obman, A.; Stout, J.E.; Yu, V.L. Isolation of Staphylococcus aureus from the urinary tract: Association of isolation with symptomatic urinary tract infection and subsequent staphylococcal bacteremia. Clin. Infect. Dis. 2006, 42, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Grillo, S.; Cuervo, G.; Carratala, J.; Grau, I.; Llaberia, M.; Aguado, J.M.; Lopez-Cortes, L.E.; Lalueza, A.; Sanjuan, R.; Sanchez-Batanero, A.; et al. Characteristics and Outcomes of Staphylococcus aureus Bloodstream Infection Originating from the Urinary Tract: A Multicenter Cohort Study. Open Forum Infect. Dis. 2020, 7, ofaa216. [Google Scholar] [CrossRef] [PubMed]
- Lafon, T.; Hernandez Padilla, A.C.; Baisse, A.; Lavaud, L.; Goudelin, M.; Barraud, O.; Daix, T.; Francois, B.; Vignon, P. Community-acquired Staphylococcus aureus bacteriuria: A warning microbiological marker for infective endocarditis? BMC Infect. Dis. 2019, 19, 504. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.N.; Flores-Mireles, A.L.; Pinkner, C.L.; Schreiber, H.L.; Joens, M.S.; Park, A.M.; Potretzke, A.M.; Bauman, T.M.; Pinkner, J.S.; Fitzpatrick, J.A.J.; et al. Catheterization alters bladder ecology to potentiate Staphylococcus aureus infection of the urinary tract. Proc. Natl. Acad. Sci. USA 2017, 114, E8721–E8730. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.; Bagale, K.; Patel, S.; Kooyers, N.J.; Kulkarni, R. Human Urine Alters Methicillin-Resistant Staphylococcus aureus Virulence and Transcriptome. Appl. Environ. Microbiol. 2021, 87, e0074421. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.; Guedry, S.; Obernuefemann, C.L.P.; Hultgren, S.J.; Walker, J.N.; Kulkarni, R. Defining the Roles of Pyruvate Oxidation, TCA Cycle, and Mannitol Metabolism in Methicillin-Resistant Staphylococcus aureus Catheter-Associated Urinary Tract Infection. Microbiol. Spectr. 2023, 11, e0536522. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, P.; Faure, E.; Kipnis, E. Inflammasomes in tissue damages and immune disorders after trauma. Front. Immunol. 2018, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Schaale, K.; Peters, K.M.; Murthy, A.M.; Fritzsche, A.K.; Phan, M.D.; Totsika, M.; Robertson, A.A.; Nichols, K.B.; Cooper, M.A.; Stacey, K.J.; et al. Strain- and host species-specific inflammasome activation, IL-1beta release, and cell death in macrophages infected with uropathogenic Escherichia coli. Mucosal Immunol. 2016, 9, 124–136. [Google Scholar] [CrossRef]
- Nagamatsu, K.; Hannan, T.J.; Guest, R.L.; Kostakioti, M.; Hadjifrangiskou, M.; Binkley, J.; Dodson, K.; Raivio, T.L.; Hultgren, S.J. Dysregulation of Escherichia coli alpha-hemolysin expression alters the course of acute and persistent urinary tract infection. Proc. Natl. Acad. Sci. USA 2015, 112, E871–E880. [Google Scholar] [CrossRef]
- Demirel, I.; Persson, A.; Brauner, A.; Sarndahl, E.; Kruse, R.; Persson, K. Activation of the NLRP3 Inflammasome Pathway by Uropathogenic Escherichia coli Is Virulence Factor-Dependent and Influences Colonization of Bladder Epithelial Cells. Front. Cell Infect. Microbiol. 2018, 8, 81. [Google Scholar] [CrossRef]
- Waldhuber, A.; Puthia, M.; Wieser, A.; Cirl, C.; Durr, S.; Neumann-Pfeifer, S.; Albrecht, S.; Rommler, F.; Muller, T.; Zheng, Y.; et al. Uropathogenic Escherichia coli strain CFT073 disrupts NLRP3 inflammasome activation. J. Clin. Investig. 2016, 126, 2425–2436. [Google Scholar] [CrossRef]
- Symington, J.W.; Wang, C.; Twentyman, J.; Owusu-Boaitey, N.; Schwendener, R.; Nunez, G.; Schilling, J.D.; Mysorekar, I.U. ATG16L1 deficiency in macrophages drives clearance of uropathogenic E. coli in an IL-1beta-dependent manner. Mucosal Immunol. 2015, 8, 1388–1399. [Google Scholar] [CrossRef]
- Demirel, I.; Persson, A.; Brauner, A.; Sarndahl, E.; Kruse, R.; Persson, K. Activation of NLRP3 by uropathogenic Escherichia coli is associated with IL-1beta release and regulation of antimicrobial properties in human neutrophils. Sci. Rep. 2020, 10, 21837. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, A.K.; Paramel, G.V.; Demirel, K.J.; Demirel, I. Human Renal Fibroblasts, but Not Renal Epithelial Cells, Induce IL-1beta Release during a Uropathogenic Escherichia coli Infection In Vitro. Cells 2021, 10, 3522. [Google Scholar] [CrossRef] [PubMed]
- Ambite, I.; Puthia, M.; Nagy, K.; Cafaro, C.; Nadeem, A.; Butler, D.S.; Rydstrom, G.; Filenko, N.A.; Wullt, B.; Miethke, T.; et al. Molecular Basis of Acute Cystitis Reveals Susceptibility Genes and Immunotherapeutic Targets. PLoS Pathog. 2016, 12, e1005848. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.J.; Conover, M.S.; Hannan, T.J.; Hultgren, S.J. Uropathogenic Escherichia coli superinfection enhances the severity of mouse bladder infection. PLoS Pathog. 2015, 11, e1004599. [Google Scholar] [CrossRef] [PubMed]
- Cohen, T.S.; Boland, M.L.; Boland, B.B.; Takahashi, V.; Tovchigrechko, A.; Lee, Y.; Wilde, A.D.; Mazaitis, M.J.; Jones-Nelson, O.; Tkaczyk, C.; et al. S. aureus Evades Macrophage Killing through NLRP3-Dependent Effects on Mitochondrial Trafficking. Cell Rep. 2018, 22, 2431–2441. [Google Scholar] [CrossRef] [PubMed]
- Craven, R.R.; Gao, X.; Allen, I.C.; Gris, D.; Wardenburg, J.B.; McElvania-TeKippe, E.; Ting, J.P.; Duncan, J.A. Staphylococcus aureus α-Hemolysin Activates the NLRP3-Inflammasome in Human and Mouse Monocytic Cells. PLoS ONE 2009, 4, e7446. [Google Scholar] [CrossRef] [PubMed]
- Holzinger, D.; Gieldon, L.; Mysore, V.; Nippe, N.; Taxman, D.J.; Duncan, J.A.; Broglie, P.M.; Marketon, K.; Austermann, J.; Vogl, T.; et al. Staphylococcus aureus Panton-Valentine leukocidin induces an inflammatory response in human phagocytes via the NLRP3 inflammasome. J. Leukoc. Biol. 2012, 92, 1069–1081. [Google Scholar] [CrossRef] [PubMed]
- Melehani, J.H.; James, D.B.A.; DuMont, A.L.; Torres, V.J.; Duncan, J.A. Staphylococcus aureus Leukocidin A/B (LukAB) Kills Human Monocytes via Host NLRP3 and ASC when Extracellular, but Not Intracellular. PLoS Pathog. 2015, 11, e1004970. [Google Scholar] [CrossRef]
- Maher, B.M.; Mulcahy, M.E.; Murphy, A.G.; Wilk, M.; O’Keeffe, K.M.; Geoghegan, J.A.; Lavelle, E.C.; McLoughlin, R.M. Nlrp-3-Driven Interleukin 17 Production by γδT Cells Controls Infection Outcomes during Staphylococcus aureus Surgical Site Infection. Infect. Immun. 2013, 81, 4478. [Google Scholar] [CrossRef]
- Hanamsagar, R.; Aldrich, A.; Kielian, T. Critical role for the AIM2 inflammasome during acute CNS bacterial infection. J. Neurochem. 2014, 129, 704–711. [Google Scholar] [CrossRef]
- Tukenmez, H.; Nye, T.M.; Bonde, M.; Caparon, M.G.; Almqvist, F.; Hultgren, S.J.; Johansson, J. Complete Genome Sequence of the Uropathogenic Methicillin-Resistant Staphylococcus aureus Strain MRSA-1369. Microbiol. Resour. Announc. 2022, 11, e0098122. [Google Scholar] [CrossRef]
- Kovarova, M.; Hesker, P.R.; Jania, L.; Nguyen, M.; Snouwaert, J.N.; Xiang, Z.; Lommatzsch, S.E.; Huang, M.T.; Ting, J.P.; Koller, B.H. NLRP1-dependent pyroptosis leads to acute lung injury and morbidity in mice. J. Immunol. 2012, 189, 2006–2016. [Google Scholar] [CrossRef]
- Miyoshi, J.; Leone, V.; Nobutani, K.; Musch, M.W.; Martinez-Guryn, K.; Wang, Y.; Miyoshi, S.; Bobe, A.M.; Eren, A.M.; Chang, E.B. Minimizing confounders and increasing data quality in murine models for studies of the gut microbiome. PeerJ 2018, 6, e5166. [Google Scholar] [CrossRef]
- Hung, C.S.; Dodson, K.W.; Hultgren, S.J. A murine model of urinary tract infection. Nat. Protoc. 2009, 4, 1230–1243. [Google Scholar] [CrossRef] [PubMed]
- Ingersoll, M.A.; Kline, K.A.; Nielsen, H.V.; Hultgren, S.J. G-CSF induction early in uropathogenic Escherichia coli infection of the urinary tract modulates host immunity. Cell. Microbiol. 2008, 10, 2568–2578. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, C.E.; Smith, S.N.; Johnson, A.O.; DeOrnellas, V.; Eaton, K.A.; Yep, A.; Mody, L.; Wu, W.; Mobley, H.L.T. The pathogenic potential of Proteus mirabilis is enhanced by other uropathogens during polymicrobial urinary tract infection. Infect. Immun. 2017, 85, e00808-16. [Google Scholar] [CrossRef] [PubMed]
- Saenkham-Huntsinger, P.; Hyre, A.N.; Hanson, B.S.; Donati, G.L.; Adams, L.G.; Ryan, C.; Londono, A.; Moustafa, A.M.; Planet, P.J.; Subashchandrabose, S. Copper Resistance Promotes Fitness of Methicillin-Resistant Staphylococcus aureus during Urinary Tract Infection. mBio 2021, 12, e0203821. [Google Scholar] [CrossRef] [PubMed]
- Coll, R.C.; Hill, J.R.; Day, C.J.; Zamoshnikova, A.; Boucher, D.; Massey, N.L.; Chitty, J.L.; Fraser, J.A.; Jennings, M.P.; Robertson, A.A.B.; et al. MCC950 directly targets the NLRP3 ATP-hydrolysis motif for inflammasome inhibition. Nat. Chem. Biol. 2019, 15, 556–559. [Google Scholar] [CrossRef]
- Coll, R.C.; Robertson, A.A.B.; Chae, J.J.; Higgins, S.C.; Muñoz-Planillo, R.; Inserra, M.C.; Vetter, I.; Dungan, L.S.; Monks, B.G.; Stutz, A.; et al. A small molecule inhibitior of the NLRP3 inflammasome is a potential therapeutic for inflammatory diseases. Nat. Med. 2015, 21, 248. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Conjugate | Clone |
|---|---|---|
| CD64 | BV 786 | X54-5/7.1 |
| SiglecF | BV711 | M290 |
| CD3 | BV650 | 145-2C11 |
| CD45 | BV605 | 30-F11 |
| Live/Dead | Alexa fluor 430 | |
| Fc epsilon RI | Pac Blue | MAR1 |
| Ly6C | PE-Cy7 | HK1.4 |
| CD11c | PerCp-Cy5.5 | N418 |
| CD103 | PE-CF594 | M290 |
| c-kit (CD117) | PE | 2B8 |
| CD11b | FITC | M1/70 |
| MHC-II (I-A/I-E) | APC-Fire750 | M5/114.15.2 |
| F4/80 | Alexa Fluor 700 | CI:A3-1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paudel, S.; Kumar, R.; Rogers, K.A.; Saini, Y.; Patial, S.; Kulkarni, R. The NLRP3 Inflammasome Is Dispensable in Methicillin-Resistant Staphylococcus aureus Urinary Tract Infection. Pathogens 2024, 13, 106. https://doi.org/10.3390/pathogens13020106
Paudel S, Kumar R, Rogers KA, Saini Y, Patial S, Kulkarni R. The NLRP3 Inflammasome Is Dispensable in Methicillin-Resistant Staphylococcus aureus Urinary Tract Infection. Pathogens. 2024; 13(2):106. https://doi.org/10.3390/pathogens13020106
Chicago/Turabian StylePaudel, Santosh, Rahul Kumar, Kenneth A. Rogers, Yogesh Saini, Sonika Patial, and Ritwij Kulkarni. 2024. "The NLRP3 Inflammasome Is Dispensable in Methicillin-Resistant Staphylococcus aureus Urinary Tract Infection" Pathogens 13, no. 2: 106. https://doi.org/10.3390/pathogens13020106