
Association of SFG Rickettsia massiliae and Candidatus Rickettsia shennongii with Different Hard Ticks Infesting Livestock Hosts

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Tick Sampling and Identification
2.3. DNA Extraction and PCR
2.4. DNA Sequencing and Phylogenetic Analysis
3. Results
3.1. Occurrence of Morphologically Identified Ticks
3.2. Detection of Rickettsia spp. in Ticks
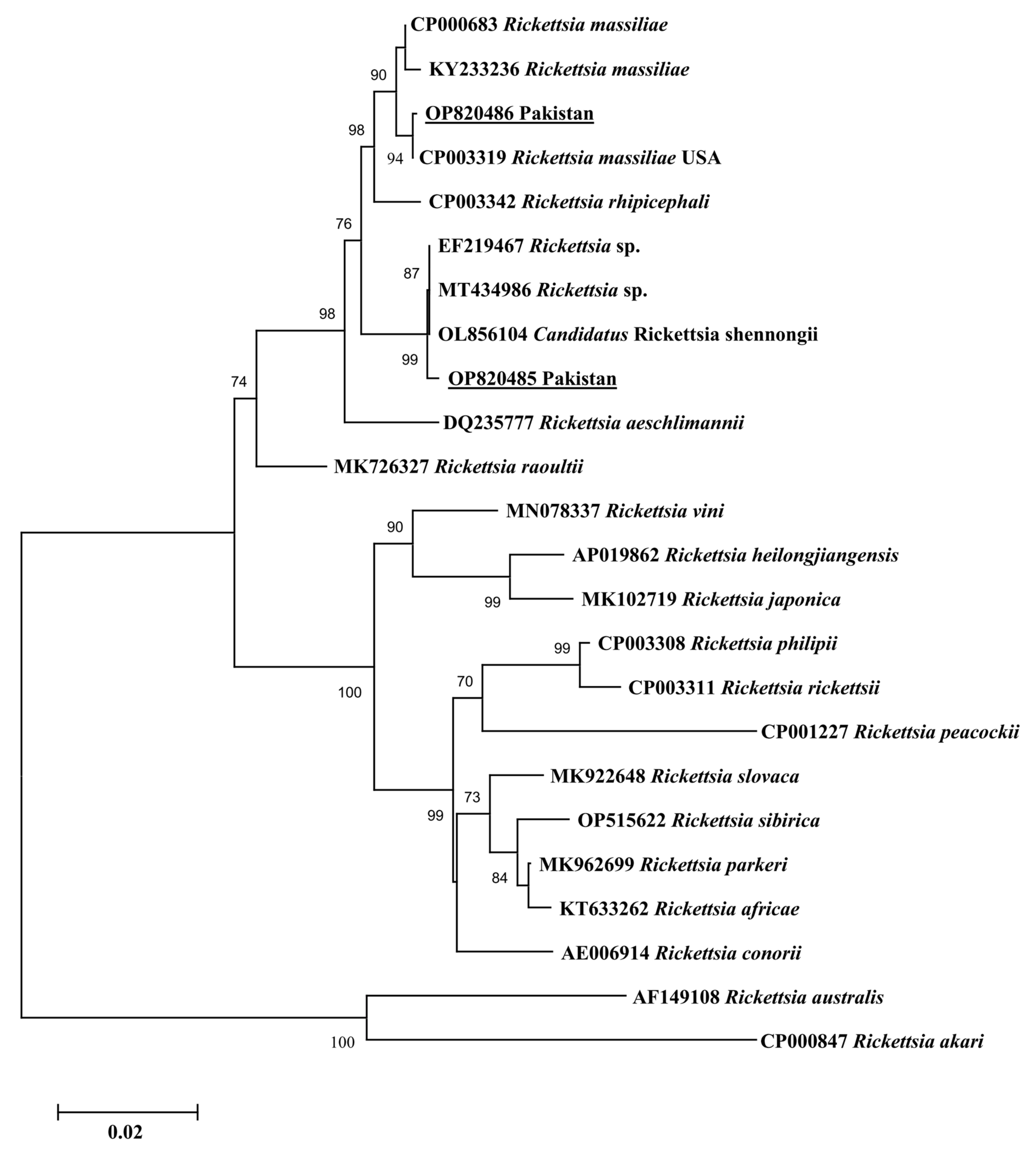
3.3. Sequencing and Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, J.; Farris, C.M.; Yeh, K.B.; Richards, A.L. International Rickettsia Disease surveillance: An example of cooperative research to increase laboratory capability and capacity for risk assessment of rickettsial outbreaks worldwide. Front. Med. 2021, 8, 622015. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Zahid, H.; Zeb, I.; Tufail, M.; Khan, S.; Haroon, M.; Bilal, M.; Hussain, M.; Alouffi, A.S.; Muñoz-Leal, S.; et al. Risk factors associated with tick infestations on equids in Khyber Pakhtunkhwa, Pakistan, with notes on Rickettsia massiliae detection. Parasites Vectors 2021, 14, 363. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Shehla, S.; Zahid, H.; Ullah, F.; Zeb, I.; Ahmed, H.; Da Silva Vaz, I., Jr.; Tanaka, T. Molecular survey and spatial distribution of Rickettsia spp. in ticks infesting free-ranging wild animals in Pakistan (2017–2021). Pathogens 2022, 11, 162. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.; Budachetri, K.; Mukherjee, N.; Williams, J.; Kausar, A.; Hassan, M.J.; Adamson, S.; Dowd, S.E.; Apanskevich, D.; Arijo, A.; et al. A study of ticks and tick-borne livestock pathogens in Pakistan. PLoS Negl. Trop. Dis. 2017, 11, e0005681. [Google Scholar] [CrossRef] [PubMed]
- García-García, J.C.; Portillo, A.; Núñez, M.J.; Santibáñez, S.; Castro, B.; Oteo, J.A. Case report: A patient from Argentina infected with Rickettsia massiliae. Am. J. Trop. Med. Hyg. 2010, 82, 691. [Google Scholar] [CrossRef] [PubMed]
- Dumic, I.; Jevtic, D.; Veselinovic, M.; Nordstrom, C.W.; Jovanovic, M.; Mogulla, V.; Veselinovic, E.M.; Hudson, A.; Simeunovic, G.; Petcu, E.; et al. Human granulocytic anaplasmosis—A systematic review of published cases. Microorganisms 2022, 10, 1433. [Google Scholar] [CrossRef]
- Khan, Z.; Shehla, S.; Alouffi, A.; Kashif Obaid, M.; Zeb Khan, A.; Almutairi, M.M.; Numan, M.; Aiman, O.; Alam, S.; Ullah, S.; et al. Molecular survey and genetic characterization of Anaplasma marginale in ticks collected from livestock hosts in Pakistan. Animals 2022, 12, 1708. [Google Scholar] [CrossRef]
- Binetruy, F.; Buysse, M.; Barosi, R.; Duron, O. Novel Rickettsia genotypes in ticks in French Guiana, South America. Sci. Rep. 2020, 10, 2537. [Google Scholar] [CrossRef]
- Weinert, L.A.; Morand, S.; Krasnov, B.R.; Littlewood, D.T.J. The diversity and phylogeny of Rickettsia. In Parasite Diversity and Diversification: Evolutionary Ecology Meets Phylogenetics; Cambridge University Press: Cambridge, UK, 2015; pp. 150–181. [Google Scholar]
- El Karkouri, K.; Ghigo, E.; Raoult, D.; Fournier, P.E. Genomic evolution and adaptation of arthropod-associated Rickettsia. Sci. Rep. 2022, 12, 3807. [Google Scholar] [CrossRef]
- Lu, M.; Tian, J.; Wang, W.; Zhao, H.; Jiang, H.; Han, J.; Guo, W.; Li, K. High diversity of Rickettsia spp., Anaplasma spp., and Ehrlichia spp. in ticks from Yunnan Province, Southwest China. Front. Microbiol. 2022, 13, 1008110. [Google Scholar] [CrossRef] [PubMed]
- Beati, L.; Finidori, J.P.; Gilot, B.; Raoult, D. Comparison of serologic typing, sodium dodecyl sulfate-polyacrylamide gel electrophoresis protein analysis, and genetic restriction fragment length polymorphism analysis for identification of Rickettsiae: Characterization of two new rickettsial strains. J. Clin. Microbiol. 1992, 30, 1922–1930. [Google Scholar] [CrossRef] [PubMed]
- Blanda, V.; Torina, A.; La Russa, F.; D’Agostino, R.; Randazzo, K.; Scimeca, S.; Giudice, E.; Caracappa, S.; Cascio, A.; de la Fuente, J. A retrospective study of the characterization of Rickettsia species in ticks collected from humans. Ticks Tick-Borne Dis. 2017, 8, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Chisu, V.; Leulmi, H.; Masala, G.; Piredda, M.; Foxi, C.; Parola, P. Detection of Rickettsia hoogstraalii, Rickettsia helvetica, Rickettsia massiliae, Rickettsia slovaca and Rickettsia aeschlimannii in ticks from Sardinia, Italy. Ticks Tick-Borne Dis. 2017, 8, 347–352. [Google Scholar] [CrossRef]
- Beeler, E.; Abramowicz, K.F.; Zambrano, M.L.; Sturgeon, M.M.; Khalaf, N.; Hu, R.; Dasch, G.A.; Eremeeva, M.E. A focus of dogs and Rickettsia massiliae–infected Rhipicephalus sanguineus in California. Am. J. Trop. Med. Hyg. 2011, 84, 244. [Google Scholar] [CrossRef]
- Cicuttin, G.L.; Brambati, D.F.; Eugui, J.I.R.; Lebrero, C.G.; De Salvo, M.N.; Beltrán, F.J.; Dohmen, F.E.G.; Jado, I.; Anda, P. Molecular characterization of Rickettsia massiliae and Anaplasma platys infecting Rhipicephalus sanguineus ticks and domestic dogs, Buenos Aires (Argentina). Ticks Tick-Borne Dis. 2014, 5, 484–488. [Google Scholar] [CrossRef]
- López-Pérez, A.M.; Sánchez-Montes, S.; Foley, J.; Guzmán-Cornejo, C.; Colunga-Salas, P.; Pascoe, E.; Becker, I.; Delgado-de la Mora, J.; Licona-Enriquez, J.D.; Suzan, G. Molecular evidence of Borrelia burgdorferi sensu stricto and Rickettsia massiliae in ticks collected from a domestic-wild carnivore interface in Chihuahua, Mexico. Ticks Tick-Borne Dis. 2019, 10, 1118–1123. [Google Scholar] [CrossRef]
- Wei, Q.Q.; Guo, L.P.; Wang, A.D.; Mu, L.M.; Zhang, K.; Chen, C.F.; Zhang, W.J.; Wang, Y.Z. The first detection of Rickettsia aeschlimannii and Rickettsia massiliae in Rhipicephalus turanicus ticks, in northwest China. Parasites Vectors 2015, 8, 631. [Google Scholar] [CrossRef]
- Krishnamoorthy, P.; Sudhagar, S.; Goudar, A.L.; Jacob, S.S.; Suresh, K.P. Molecular survey and phylogenetic analysis of tick-borne pathogens in ticks infesting cattle from two South Indian states. Vet. Parasitol. Reg. Stud. 2021, 25, 100595. [Google Scholar] [CrossRef]
- Mostafavi, S.M.; Khalili, M.; Akhtardanesh, B.; Nourollahifard, S.R.; Esmaeili, S. Rickettsia spp. in Rhipicephalus sanguineus sensu lato ticks collected from stray dogs in Kerman city, Iran. Ticks Tick-Borne Dis. 2022, 13, 101985. [Google Scholar] [CrossRef]
- Numan, M.; Islam, N.; Adnan, M.; Safi, S.Z.; Chitimia-Dobler, L.; Labruna, M.B.; Ali, A. First genetic report of Ixodes kashmiricus and associated Rickettsia sp. Parasites Vectors 2022, 15, 378. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Islam, N.; Khan, A.; Islam, Z.U.; Muñoz-Leal, S.; Labruna, M.B.; Ali, A. New records of Amblyomma gervaisi from Pakistan, with detection of a reptile-associated Borrelia sp. Ticks Tick-Borne Dis. 2022, 13, 102047. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Ullah, S.; Alouffi, A.; Almutairi, M.M.; Khan, M.; Numan, M.; Safi, S.Z.; Chitimia-Dobler, L.; Tanaka, T.; Ali, A. Description of Male, Redescription of Female, Host Record, and Phylogenetic Position of Haemaphysalis danieli. Pathogens 2022, 11, 1495. [Google Scholar] [CrossRef] [PubMed]
- Aiman, O.; Ullah, S.; Chitimia-Dobler, L.; Nijhof, A.M.; Ali, A. First report of Nosomma monstrosum ticks infesting Asian water buffaloes (Bubalus bubalis) in Pakistan. Ticks Tick-Borne Dis. 2022, 13, 101899. [Google Scholar] [CrossRef]
- Alam, S.; Khan, M.; Alouffi, A.; Almutairi, M.M.; Ullah, S.; Numan, M.; Islam, N.; Khan, Z.; Aiman, O.; Zaman Safi, S.; et al. Spatio-Temporal Patterns of Ticks and Molecular Survey of Anaplasma marginale, with Notes on Their Phylogeny. Microorganisms 2022, 10, 1663. [Google Scholar] [CrossRef]
- Ullah, S.; Alouffi, A.; Almutairi, M.M.; Islam, N.; Rehman, G.; Ul Islam, Z.; Ahmed, H.; Vaz, I., Jr.; Labruna, M.B.; Tanaka, T.; et al. First Report of Rickettsia conorii in Hyalomma kumari Ticks. Animals 2023, 13, 1488. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Obaid, M.K.; Almutairi, M.; Alouffi, A.; Numan, M.; Ullah, S.; Rehman, G.; Islam, Z.U.; Khan, S.B.; Tanaka, T. Molecular Detection of Coxiella spp. in Ticks (Ixodidae and Argasidae) Infesting Domestic and Wild Animals: With Notes on the Epidemiology of Tick-borne Coxiella burnetii in Asia. Front. Microbiol. 2023, 14, 1229950. [Google Scholar] [CrossRef]
- Robertson, R.G.; Wisseman, C.L., Jr. Tick-Borne Rickettsiae of the Spotted Fever Group in west Pakistan: II. Serological classification of isolates from west Pakistan and Thailand: Evidence for two new species. Am. J. Epidemiol. 1973, 97, 55–64. [Google Scholar] [CrossRef]
- Ali, A.; Numan, M.; Khan, M.; Aiman, O.; Muñoz-Leal, S.; Chitimia-Dobler, L.; Labruna, M.B.; Nijhof, A.M. Ornithodoros (Pavlovskyella) ticks associated with a Rickettsia sp. in Pakistan. Parasites Vectors 2022, 15, 138. [Google Scholar] [CrossRef]
- Walker, J.B.; Keirans, J.E.; Horak, I.G. The Genus Rhipicephalus (Acari, Ixodidae): A Guide to the Brown Ticks of the World; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Apanaskevich, D.A. Differentiation of closely related species Hyalomma anatolicum and H. excavatum (Acari: Ixodidae) based on a study of all life cycle stages, throughout entire geographical range. Parazitologiia 2003, 37, 259–280. [Google Scholar]
- Apanaskevich, D.A.; Schuster, A.L.; Horak, I.G. The genus Hyalomma: VII. Redescription of all parasitic stages of H. (Euhyalomma) dromedarii and H. (E.) schulzei (Acari: Ixodidae). J. Med. Entomol. 2008, 45, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Geevarghese, G.; Mishra, A.C. Haemaphysalis Ticks of India; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 2001; pp. 23–44. [Google Scholar]
- Labruna, M.B.; Whitworth, T.; Horta, M.C.; Bouyer, D.H.; McBride, J.W.; Pinter, A.; Popov, V.; Gennari, S.M.; Walker, D.H. Rickettsia species infecting Amblyomma cooperi ticks from an area in the state of São Paulo, Brazil, where Brazilian spotted fever is endemic. J. Clin. Microbiol. 2004, 42, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Roux, V.; Fournier, P.E.; Raoult, D. Differentiation of spotted fever group Rickettsiae by sequencing and analysis of restriction fragment length polymorphism of PCR-amplified DNA of the gene encoding the protein rompA. J. Clin. Microbiol. 1996, 34, 2058. [Google Scholar] [CrossRef] [PubMed]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Matsumoto, K.; Ogawa, M.; Brouqui, P.; Raoult, D.; Parola, P. Transmission of Rickettsia massiliae in the tick, Rhipicephalus turanicus. Med. Vet. Entomol. 2005, 19, 263–270. [Google Scholar] [CrossRef]
- Eldin, C.; Virgili, G.; Attard, L.; Edouard, S.; Viale, P.; Raoult, D.; Parola, P. Rickettsia massiliae infection after a tick bite on the eyelid. Travel. Med. Infect. Dis. 2018, 26, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Kamran, K.; Ali, A.; Villagra, C.; Siddiqui, S.; Alouffi, A.S.; Iqbal, A. A cross-sectional study of hard ticks (acari: Ixodidae) on horse farms to assess the risk factors associated with tick-borne diseases. Zoonoses Public Health 2021, 68, 247–262. [Google Scholar] [CrossRef] [PubMed]
- Vatansever, Z. Hyalomma anatolicum Koch, 1844 (Figs. 158–160). In Ticks of Europe and North Africa: A Guide to Species Identification; Springer: Cham, Switzerland, 2017; pp. 391–395. [Google Scholar]
- Nourollahi Fard, S.R.; Fathi, S.; Norouzi Asl, E.; Asgary Nazhad, H.; Salehzadeh Kazeroni, S. Hard ticks on one-humped camel (Camelus dromedarius) and their seasonal population dynamics in southeast, Iran. Trop. Anim. Health Prod. 2012, 44, 197–200. [Google Scholar] [CrossRef]
- Marié, J.L.; Davoust, B.; Socolovschi, C.; Raoult, D.; Parola, P. Molecular detection of rickettsial agents in ticks and fleas collected from a European hedgehog (Erinaceus europaeus) in Marseilles, France. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Liu, Z.; Niu, Q.; Yang, J.; Abdallah, M.O.; Chen, Z.; Liu, G.; Luo, J.; Yin, H. Molecular evidence of tick-borne pathogens in Hyalomma anatolicum ticks infesting cattle in Xinjiang Uygur Autonomous Region, Northwestern China. Exp. Appl. Acarol. 2017, 73, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Ortuño, A.; Sanfeliu, I.; Nogueras, M.M.; Pons, I.; López-Claessens, S.; Castellà, J.; Antón, E.; Segura, F. Detection of Rickettsia massiliae/Bar29 and Rickettsia conorii in red foxes (Vulpes vulpes) and their Rhipicephalus sanguineus complex ticks. Ticks Tick-Borne Dis. 2018, 9, 629–631. [Google Scholar] [CrossRef]
- Belkahia, H.; Selmi, R.; Zamiti, S.; Daaloul-Jedidi, M.; Messadi, L.; Ben Said, M. Zoonotic Rickettsia Species in Small Ruminant Ticks from Tunisia. Front. Vet. Sci. 2021, 8, 676896. [Google Scholar] [CrossRef]
- Elelu, N.; Ola-Fadunsin, S.D.; Bankole, A.A.; Raji, M.A.; Ogo, N.I.; Cutler, S.J. Prevalence of tick infestation and molecular characterization of spotted fever Rickettsia massiliae in Rhipicephalus species parasitizing domestic small ruminants in north-central Nigeria. PLoS ONE 2022, 17, 0263843. [Google Scholar] [CrossRef]
- Mesquita, J.R.; Santos-Silva, S.; de Sousa Moreira, A.; Baptista, M.B.; Cruz, R.; Esteves, F.; Vala, H.; Barradas, P.F. Rickettsia massiliae circulation in sheep and attached Rhipicephalus sanguineus in Central Portugal. Trop. Anim. Health Prod. 2022, 54, 199. [Google Scholar] [CrossRef]
- Monje, L.D.; Linares, M.C.; Beldomenico, P.M. Prevalence and infection intensity of Rickettsia massiliae in Rhipicephalus sanguineus sensu lato ticks from Mendoza, Argentina. Microbes Infect. 2016, 18, 701–705. [Google Scholar] [CrossRef]
- Nguyen, V.L.; Colella, V.; Greco, G.; Fang, F.; Nurcahyo, W.; Hadi, U.K.; Venturina, V.; Tong, K.B.Y.; Tsai, Y.L.; Taweethavonsawat, P.; et al. Molecular detection of pathogens in ticks and fleas collected from companion dogs and cats in East and Southeast Asia. Parasites Vectors 2020, 13, 420. [Google Scholar] [CrossRef] [PubMed]
- Klein, D.; Beth-Din, A.; Cohen, R.; Lazar, S.; Glinert, I.; Zayyad, H.; Atiya-Nasagi, Y. New spotted fever group Rickettsia isolate, identified by sequence analysis of conserved genomic regions. Pathogens 2019, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.P.; Jiang, S.H.; Liu, D.; Wang, S.W.; Chen, C.F.; Wang, Y.Z. Emerging spotted fever group Rickettsiae in ticks, northwestern China. Ticks Tick-Borne Dis. 2016, 7, 1146–1150. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) | Amplicon Size | Reference |
|---|---|---|---|---|
| gltA | CS-78 | GCAAGTATCGGTGAGGATGTAAT | 401 bp | [36] |
| CS-323 | GCTTCCTTAAAATTCAATAAATCAGGAT | |||
| ompA | Rr190.70p | ATGGCGAATATTTCTCCAAAA | 631 bp | [37] |
| Rr190.701n | GTTCCGTTAATGGCAGCATCT | |||
| ompB | 120-M59 | CCGCAGGGTTGGTAACTGC | 862 bp | [38] |
| 120-807 | CCTTTTAGATTACCGCCTAA |
| District | Hosts | No. of Infested/No. of Examined Livestock Hosts | Identified Ticks | No. of Ticks According to Life Stage | No. of Tested Ticks | Molecular Detection of Rickettsia spp. In Morphologically Identified Ticks (N, F, M) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of Ticks with R. massiliae | No. of Ticks with Ca. R. shennongii | ||||||||||
| gltA | ompA | ompB | gltA | ompA | ompB | ||||||
| Charsadda | Equids | 4/9 | Rh. turanicus | 1N, 2F | 1N, 1F | 1N | 1N | 1N | |||
| Rh. haemaphysaloides | 3N, 1M, 1F | 1N, 1M, 1F | |||||||||
| Cattle | 14/20 | Rh. turanicus | 1N, 1M, 2F | 1M, 1F | |||||||
| Hy. anatolicum | 2N, 2F | 1N, 1F | |||||||||
| Rh. haemaphysaloides | 4N, 1M, 3F | 1N, 1M, 1F | 1F | ||||||||
| Rh. microplus | 7N, 2M, 11F | 1N, 1M, 1F | |||||||||
| Buffaloes | 8/15 | Hy. anatolicum | 1N, 1M, 3F | 1N, 1M, 1F | |||||||
| Rh. turanicus | 1N, 1M, 1F | 1M, 1F | 1F | 1F | |||||||
| Sheep | 8/20 | Hy. anatolicum | 1N, 1M, 2F | 1N, 1F | |||||||
| Rh. turanicus | 2N, 1M, 2F | 1M | |||||||||
| Goats | 9/15 | Hy. anatolicum | 1N, 1M, 1F | 1N, 1F | |||||||
| Rh. turanicus | 2N, 1M, 3F | 1N, 1M, 1F | |||||||||
| Rh. haemaphysaloides | 3N, 2M, 6F | 1N, 1M, 1F | |||||||||
| Camels | 6/10 | Hy. dromedarii | 3N, 2M, 5F | 1N, 1M, 1F | |||||||
| Total | 49/89 | 91 | 34 | 3 | |||||||
| Peshawar | Equids | 4/11 | Rh. turanicus | 2N, 1M, 2F | 1N, 1M | ||||||
| Cattle | 13/19 | Hy. anatolicum | 4N, 2M, 5F | 1N, 1M, 1F | |||||||
| Rh. haemaphysaloides | 2N, 2M, 4F | 1N, 1M, 1F | |||||||||
| Buffaloes | 5/9 | Hy. anatolicum | 2N, 1M, 3F | 1N, 1M, 1F | |||||||
| Rh. turanicus | 3N, 1M, 3F | 1N, 1M, 1F | |||||||||
| Sheep | 6/8 | Rh. turanicus | 3N, 1M, 3F | 1N, 1M, 1F | |||||||
| Goats | 5/7 | Rh. haemaphysaloides | 4N, 1M, 4F | 1N, 1M, 1F | 1N | 1N | 1N | ||||
| Rh. microplus | 3N, 2M, 6F | 1N, 1M, 1F | 1N | 1N | 1N | ||||||
| Camels | 2/4 | Hy. dromedarii | 3N, 1M, 5F | 1N, 1M, 5F | |||||||
| Total | 35/58 | 73 | 30 | 2 | |||||||
| Mardan | Equids | 3/5 | Rh. turanicus | 1N, 2M, 5F | 1N, 1M, 1F | 1M | 1M | ||||
| Cattle | 11/15 | Hy. anatolicum | 1N, 2M, 2F | 1N, 1M, 1F | |||||||
| Rh. turanicus | 1N, 2M, 3F | 1N, 1M, 1F | 1N | 1N | 1N | ||||||
| Rh. microplus | 7N, 4M, 13F | 1N, 1M, 1F | |||||||||
| Buffaloes | 5/10 | Rh. haemaphysaloides | 2N, 2M, 3F | 1N, 1M, 1F | |||||||
| Sheep | 4/10 | Rh. haemaphysaloides | 2N, 2M, 2F | 1N, 1M, 1F | 1N | 1N | |||||
| Goats | 4/8 | Rh. haemaphysaloides | 1N, 2M | 1N, 1M | |||||||
| Rh. turanicus | 3N, 2M, 1F | 1N, 1M, 1F | 1N | 1N | 1N | ||||||
| Camels | 2/4 | Hy. dromedarii | 1N, 2M, 2F | 1N, 1M, 1F | |||||||
| Total | 29/52 | 70 | 26 | 4 | |||||||
| Swabi | Equids | 3/5 | Rh. turanicus | 1N, 2M, 1F | 1N, 1M, 1F | ||||||
| Cattle | 9/9 | Rh. microplus | 5N, 3M, 7F | 3N, 1M, 1F | |||||||
| Buffaloes | 8/8 | Hy. anatolicum | 1N, 2M, 3F | 1N, 1M, 1F | |||||||
| Rh. turanicus | 1N, 1M, 1F | 1N, 1M, 1F | 1N | 1N | 1N | ||||||
| Sheep | 5/7 | Rh. haemaphysaloides | 1N, 2M, 3F | 1N, 1M, 1F | |||||||
| Rh. turanicus | 1N, 1M, 2F | 1N, 1M, 1F | |||||||||
| Goats | 5/6 | Rh. microplus | 2N, 3M, 5F | 1N, 2M, 2F | 1M | 1M | |||||
| Camels | 2/4 | Hy. dromedarii | 2N, 3M, 4F | 1N, 1M, 1F | |||||||
| Total | 32/39 | 57 | 28 | 2 | |||||||
| Lakki Marwat | Equids | 3/4 | Rh. turanicus | 1N, 1M, 2F | 1N, 1M, 1F | ||||||
| Cattle | 7/8 | Hy. tnatolicum | 1N, 1M, 1F | 1N, 1M, 1F | |||||||
| Rh. turanicus | 3N, 1M, 1F | 1N, 1F | |||||||||
| Rh. microplus | 3N, 2M, 6F | 1N, 2M | |||||||||
| Buffaloes | 5/7 | Hy. anatolicum | 1N, 2M, 3F | 1N, 1F | |||||||
| Sheep | 6/10 | Hy. anatolicum | 3N, 1M, 4F | 1N, 1F | |||||||
| Rh. turanicus | 1N, 1M, 2F | 1N, 1M, 1F | |||||||||
| Goats | 5/7 | Hy. anatolicum | 1N, 2M, 3F | 1N, 1M, 1F | |||||||
| Rh. haemaphysaloides | 2N, 2M, 4F | 1N, 1M, 1F | 1N | 1N | 1N | ||||||
| Camels | 2/3 | Hy. dromedarii | 2N, 2M, 3F | 1N, 1F | |||||||
| Total | 28/39 | 62 | 26 | 1 | |||||||
| Bannu | Equids | 0/2 | None | None | None | ||||||
| Cattle | 7/7 | Hy. anatolicum | 1N, 2M, 3F | 1N, 1M, 1F | |||||||
| Rh. haemaphysaloides | 3N, 1M, 5F | 1N, 1M, 1F | |||||||||
| Rh. microplus | 3N, 2M, 5F | 1N, 1M, 1F | |||||||||
| Buffaloes | 4/6 | Rh. turanicus | 1N, 1M, 2F | 1N, 1M, 1F | 1N | 1N | 1N | ||||
| Sheep | 3/9 | Hy. anatolicum | 1N, 2M | 1N, 1M | |||||||
| Goats | 5/7 | Hy. anatolicum | 1N, 2M, 2F | 1N, 1M, 1F | |||||||
| Camels | 2/4 | Hy. dromedarii | 2N, 1M, 2F | 1N, 1M, 1F | |||||||
| Total | 21/35 | 42 | 20 | 1 | |||||||
| Nowshera | Equids | 2/2 | Rh. turanicus | 1N, 1M, 1F | 1N, 1M, 1F | ||||||
| Cattle | 8/9 | Hy. anatolicum | 1N, 2M, 3F | 1N, 1M, 1F | |||||||
| Rh. microplus | 4N, 2M, 6F | 1N, 1M, 1F | |||||||||
| Buffaloes | 6/6 | Hy. anatolicum | 1N, 2M, 1F | 1N, 1M, 1F | |||||||
| Rh. turanicus | 1N, 1M, 1F | 1N, 1F | 1N | 1N | 1N | ||||||
| Sheep | 6/7 | Hy. anatolicum | 4N, 1M, 5F | 1N, 1M, 1F | |||||||
| Goats | 5/7 | Rh. haemaphysaloides | 4N, 1M, 6F | 1N, 1M, 1F | 1M | 1M | |||||
| Ha. sulcata | 1N, 1M, 2F | 1N, 1M, 1F | |||||||||
| Camels | 2/4 | Hy. dromedarii | 1N, 1M, 2F | 1N, 1M, 1F | |||||||
| Total | 29/35 | 57 | 26 | 2 | |||||||
| Bajaur | Equids | 2/2 | Rh. turanicus | 1N, 2M, 1F | 1N, 1M, 1F | ||||||
| Cattle | 6/7 | Rh. turanicus | 1N, 2M, 2F | 1N, 1M, 1F | 1F | 1F | |||||
| Rh. microplus | 5N, 1M, 5F | 1N, 1M, 1F | |||||||||
| Buffaloes | 4/5 | Hy. anatolicum | 1N, 2M, 3F | 1N, 1M, 1F | |||||||
| Rh. turanicus | 1N, 1M, 1F | 1N, 1M, 1F | |||||||||
| Sheep | 4/6 | Rh. turanicus | 1N, 1M, 2F | 1N, 1M, 2F | |||||||
| Rh. haemaphysaloides | 2N, 2M, 3F | 2N, 1M, 1F | 1F | 1F | |||||||
| Goats | 3/5 | Hy. anatolicum | 1N, 1M, 2F | 1N, 1M, 1F | |||||||
| Rh. haemaphysaloides | 1N, 2M, 2F | 1N, 1M, 1F | 1N | 1N | 1N | ||||||
| Camels | 1/2 | Hy. dromedarii | 1N, 1M, 2F | 1N, 1F | |||||||
| Total | 20/27 | 53 | 33 | 3 | |||||||
| Malakand | Equids | 2/5 | Rh. turanicus | 1N, 1M, 2F | 1N, 1F | ||||||
| Cattle | 4/11 | Rh. turanicus | 1N, 1M, 2F | 1N, 1M, 1F | |||||||
| Buffaloes | 3/8 | Hy. anatolicum | 1N, 1M, 1F | 1N, 1F | |||||||
| Sheep | 4/9 | Ha. cornupunctata | 1N, 2M, 3F | 1N, 2M,3F | 1N | 1N | 1N | ||||
| Goats | 3/6 | Hy. anatolicum | 1N, 2M | 1N, 1M | |||||||
| Camels | 3/5 | Hy. dromedarii | 4N, 1M, 4F | 1N, 1M | |||||||
| Total | 19/44 | 29 | 18 | 1 | |||||||
| Mohmand | Equid | 3/4 | Rh. turanicus | 1N, 1M, 3F | 1N, 1M, 1F | ||||||
| Cattle | 6/7 | Hy. anatolicum | 1N, 1M, 1F | 1N, 1F | |||||||
| Ha. cornupunctata | 4N, 1M, 4F | 1N, 1M, 1F | |||||||||
| Buffaloes | 2/8 | Hy. anatolicum | 1N, 1M | 1N, 1M | |||||||
| Sheep | 5/10 | Rh. turanicus | 1N, 1M, 1F | 1N, 1F | 1N | 1N | 1N | ||||
| Rh. haemaphysaloides | 1N, 2M | 1N, 1M | |||||||||
| Goats | 4/6 | Rh. haemaphysaloides | 1N, 1M, 1F | 1N, 1F | 1F | 1F | |||||
| Ha. sulcata | 1N, 1M | 1N, 1M | |||||||||
| Camels | 3/4 | Hy. dromedarii | 3N, 1M, 3F | 1N, 1M, 1F | |||||||
| Total | 23/39 | 37 | 20 | 2 | |||||||
| Lower Dir | Equids | 2/6 | Rh. turanicus | 1N, 1M, 1F | 1M, 1F | ||||||
| Cattle | 7/8 | Hy. anatolicum | 1N, 1F | 1N, 1F | |||||||
| Rh. microplus | 4N, 1M, 5F | 2N, 1M, 2F | |||||||||
| Buffaloes | 4/6 | Rh. turanicus | 1N, 1M, 2F | 1N, 1M, 1F | 1F | 1F | |||||
| Sheep | 7/10 | Ha. cornupunctata | 1N, 2M, 1F | 1N, 1M, 1F | |||||||
| Rh. turanicus | 1N, 1M, 2F | 1N, 1F | 1N | 1N | 1N | ||||||
| Goats | 6/9 | Ha. cornupunctata | 1N, 1M, 1F | 1N, 1M, 1F | |||||||
| Rh. haemaphysaloides | 1N, 1M, 2F | 1N, 2F | |||||||||
| Total | 26/39 | 34 | 23 | 2 | |||||||
| Buner | Equids | 3/4 | Rh. turanicus | 1N, 2M, 3F | 1M, 1F | ||||||
| Cattle | 6/7 | Rh. microplus | 3N, 2M, 4F | 1N, 2F | |||||||
| Buffaloes | 9/10 | Rh. turanicus | 1N, 1M, 1F | 1N, 1F | |||||||
| Hy. anatolicum | 1N, 1M, 1F | 1N, 1M, 1F | |||||||||
| Rh. microplus | 4N, 2M, 5F | 3N, 1F | |||||||||
| Sheep | 6/7 | Ha. sulcata | 1N, 1M, 1F | 1N, 1M, 1F | |||||||
| Rh. turanicus | 1N, 2M, 2F | 1N, 1M, 1F | |||||||||
| Goats | 5/5 | Rh. turanicus | 1N, 1M, 1F | 1N, 1M | |||||||
| Rh. haemaphysaloides | 2N, 1M, 2F | 1N, 1F | 1F | 1F | |||||||
| Camels | 4/7 | Hy. dromedarii | 1N, 1M, 3F | 1N, 1M, 1F | |||||||
| Total | 33/40 | 53 | 27 | 1 | |||||||
| Swat | Equids | 2/5 | Rh. turanicus | 1N, 1M, 1F | 1N, 1F | ||||||
| Cattle | 7/9 | Rh. microplus | 4N, 1M, 6F | 1N, 1M, 1F | |||||||
| Buffaloes | 8/10 | Rh. turanicus | 1N, 1M, 1F | 1N, 1M, 1F | |||||||
| Rh. microplus | 5N, 2M, 7F | 1N, 1M,1F | |||||||||
| Sheep | 6/8 | Hy. anatolicum | 1N, 1M, 1F | 1N, 1F | |||||||
| Rh. turanicus | 1N, 2M, 2F | 1N, 1M, 1F | 1N | 1N | 1N | ||||||
| Goats | 6/7 | Hy. anatolicum | 1N, 1M, 2F | 1N, 1F | |||||||
| Ha. cornupunctata | 1N, 2M, 3F | 1N, 1M, 1F | |||||||||
| Total | 29/39 | 49 | 19 | 1 | |||||||
| Total | 373/575 (64.9%) | Rh. turanicus (163, 23.1%), Rh. microplus (179, 25.3%), Hy. anatolicum (136, 19.2%), Rh. haemaphysaloides (118, 16.6%), Hy. dromedarii (74, 10.5%), Ha. sulcata (9, 1.3%), Ha. cornupunctata (28, 1.8%) | 707 (219N, 320F, 168M) | 330 (118N, 95M, 117F) | 4/330 (1.2%) | 25/330 (7.6%) | |||||
| 25/330 (7.6%) Rh. haemaphysaloides (9/50, 18.0%) Rh. turanicus (13/99, 13.1%), Rh. microplus (2/49, 4.1%) Ha. cornupunctata (1/18, 5.5%), | |||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shehla, S.; Ullah, F.; Alouffi, A.; Almutairi, M.M.; Khan, Z.; Tanaka, T.; Labruna, M.B.; Tsai, K.-H.; Ali, A. Association of SFG Rickettsia massiliae and Candidatus Rickettsia shennongii with Different Hard Ticks Infesting Livestock Hosts. Pathogens 2023, 12, 1080. https://doi.org/10.3390/pathogens12091080
Shehla S, Ullah F, Alouffi A, Almutairi MM, Khan Z, Tanaka T, Labruna MB, Tsai K-H, Ali A. Association of SFG Rickettsia massiliae and Candidatus Rickettsia shennongii with Different Hard Ticks Infesting Livestock Hosts. Pathogens. 2023; 12(9):1080. https://doi.org/10.3390/pathogens12091080
Chicago/Turabian StyleShehla, Shehla, Farman Ullah, Abdulaziz Alouffi, Mashal M. Almutairi, Zaibullah Khan, Tetsuya Tanaka, Marcelo B. Labruna, Kun-Hsien Tsai, and Abid Ali. 2023. "Association of SFG Rickettsia massiliae and Candidatus Rickettsia shennongii with Different Hard Ticks Infesting Livestock Hosts" Pathogens 12, no. 9: 1080. https://doi.org/10.3390/pathogens12091080