
More than Three Decades of Bm86: What We Know and Where to Go


Abstract
:1. Introduction
2. The Tick Vaccine Movement (Pre-1980s): Laying the Groundwork for Proof-of-Concept
3. The Dawn of Cattle Tick Subunit Vaccines (1980s to 1990s): The Discovery of Bm86

{kind=link}
{kind=link}
| Vaccine and Major Antigen(s) | Breed | Adjuvant | Vaccination Regime | Efficacy in Bovine Vaccination Trials | Immune Markers | Reference(s) |
|---|---|---|---|---|---|---|
| Bm86-Based Recombinant Antigen Vaccines | ||||||
| Bm86 TickGARD® and TickGARD® Plus Discontinued Commercial Vaccines (Intervet Pty. Ltd., Bendigo, Australia) | -Hereford (Bos taurus taurus) [19,54,74] -Holstein Friesian (Bos taurus taurus) [60] | -Freund’s complete adjuvant [19,45] -MontanideTM (TickGARD®) [54] -Vaximax i.e., Marcol25: Montanide 888 (9:1) (TickGARD Plus®) [57,60] | Dosage: -2.3 μg (trial 1); 17 μg (trial 2) [18,45] -50 μg [51,61] Regime: -Three vaccinations at weeks 0, 4, and 8 [18,45] -Three vaccinations at weeks 0, 7, and 17; or two vaccinations at weeks 0 and 7 [51] -Three vaccinations at weeks 0, 4, and 9 [61] | REF: 20–56% Total efficacy: Ranges from 0 of >90% | Serum antibody levels: Anti-Bm86 mean IgG titres (dilution 1:4000): [60] -Day of 1st injection: 187. -2 weeks after 1st injection: 646. -2 weeks after 2nd injection: 2633. -8 weeks after 3rd injection: 582. | Patents: [75,76] Literature: [19,45,54,57,60] |
| Bm86 Gavac® Commercially available vaccine (Heber Biotec S.A., Cuba) | -Holstein Friesian (Bos taurus taurus) [58,70,73] -Crossbred (Bos taurus × Bos indicus) [55,77] -Bos taurus, Bos indicus and crossbred dairy and beef cattle (Bos taurus × Bos indicus) [56,68] | -Freund’s complete (1st vaccination) and incomplete (2nd and 3rd vaccination) adjuvant [58] -MontanideTM 888 VG (Gavac®) [55,56,59,68,70,73,77] -Saponin (saponin white pure, Merck [73] | Dosage: -100 μg [52,53,59,68,70,71] -400 μg [59] -50 μg [66] Regime: -Three vaccinations at weeks 0, 4, and 7 [52,53,59,68,70] -Three vaccinations at weeks 0, 4, and 7 followed in field trials by a maintenance dose approximately every 6 months [66] -Three vaccinations at weeks 1, 3, and 7 [71] | REF: 9–100% Total efficacy: Ranges from 0 to ≤100% | Serum antibody levels: Anti-Bm86 mean IgG titre: -2 weeks after 3rd injection (dilution n.i.): 100 × 104 [58] -2 weeks after 3rd injection (dilution n.i.): 48,000 ± 3500 [55] -3 weeks after 2nd injection (dilution n.i.): 16,000 [56] -18 weeks after 3rd injection (dilution n.i.): 6000 [56] -Average of 9 pen trials 2 weeks after 3rd injection (dilution n.i.): 6351 ± 578 [59] -2 weeks after 2nd injection (dilution 1:320): ±8000 [68] -5 weeks after 3rd injection (dilution 1:640): 7000 ± 1000 [70] Mean titres 2 weeks after 3rd injection (dilution n.i.): [73] Bm86 and MontanideTM 888 VG (Gavac®) -Total IgG: 23,744. -IgG1: 11,763 -IgG2: 1114 Bm86 and Saponin -Total IgG: 24,639 -IgG1: 8611 -IgG2: 2560 Anti-Bm86 mean OD values 2 weeks after 3rd injection (dilution 1: 1000): ± 0.3. [77] | Patent: [78] Literature: [55,56,58,59,68,70,73,77] |
| Bm95 (Bm86 strain variant) Gavac® Plus Commercially available vaccine (Heber Biotec S.A.,Cuba) | -Holstein Friesian (Bos taurus taurus) [70] -Crossbreed (n.i.) [79] -Crossbreed (Bos taurus × Bos indicus) [80] | -MontanideTM 888 [70,80] -Algel (Indian Immunological Ltd., Hyderabad) [79] | Dosage: -100 μg [68,73] -200 μg [72] Regime: -Three vaccinations at weeks 0, 4, and 7 [68] -Four vaccinations at weeks 4, 8, 12 and 16 [72] -Three vaccinations at weeks 0, 4, and 7; or two vaccinations at weeks 0 and 4; or at weeks 0 and 7 [73] | Total efficacy: -58–89% [70] -81.27% [79] Tick biological parameters: Significant reduction in weight of engorging females, weight of eggs and egg viability between vaccinated and unvaccinated groups. No significant differences observed between different vaccination schemes [80] | Serum antibody levels: Mean IgG titre 2 weeks after 2nd injection (dilution 1: 320): [70] -Anti-Bm86 ± 8000. -Anti-Bm95 ± 9000. Anti-Bm95 mean IgG titre 3 weeks after 3rd injection (dilution n.i.): [79] -7979.9 ± 312.5. Anti-Bm86 mean IgG titres (dilution n.i.): [80] Bovines vaccinated at weeks 0 and 4: -8 weeks after 2nd injection: 6000 ± 1000. -3 weeks after re-vaccination at week 24: 4000 ± 200. Bovines vaccinated at weeks 0 and 7: -5 weeks after 2nd injection: 5000 ± 100. -3 weeks after re-vaccination at week 24: 6500 ± 1000. Bovines vaccinated at weeks 0, 4, and 7: -5 weeks after 3rd injection: 7800 ± 1000. -3 weeks after re-vaccination at week 24: 6000 ± 1000. | Patent: [81,82] Literature: [70,79,80] |
| Bm86 Mozambique strain | Crossbreed (n.i.) | Montanide ISA 50V (Seppic, France) | Dosage: -100 μg Regime: -Three vaccinations at weeks 1, 3, and 7 | Total efficacy: 70.4 | Serum antibody levels: Anti-Bm86 mean OD value for IgG 2 weeks after 3rd injection (dilution 1: 1000): ±1.18. | Literature: [77] |
| Bm86-Based Epitope Vaccines | ||||||
| SBm4912® Bm86 SYNTHETIC EPITOPE CONSTRUCT | Jersey-breed (Bos Taurus) | Saponin | Dosage: -2 mg Regime: -Three vaccinations at weeks 0, 4, and 8 | Total efficacy: 72.4% | Serum antibody levels: Anti-Bm86 mean OD value for IgG 4 weeks after 3rd injection (dilution n.i.): 1.2 ± 0.4. | Patent: [83] Literature: [84] |
| SBm7462® Bm86 synthetic epitope construct | Holstein Friesian (Bos taurus taurus) | Saponin | Dosage: -2 mg Regime: -Three vaccinations at weeks 0, 4, and 8 | Total efficacy: 81.05% | Serum antibody levels: Anti-Bm86 mean OD value for 2 weeks after 3rd injection (dilution 1: 400): IgG: 1.2 ± 0.2. IgG1: 1.0 ± 0.2. Histology of LN tissue: -Increased GCs in B-cell follicles. -Hyperplasia of medullary cord. -Presence of antigens in APCs. -Apoptosis. Phenotype of circulating PBMCs: -Increased B-cells (CD21+). -Variation in WC1+ γδ T cells (CC101+). | Patent: [83] Literature: [72,84] |
| rSBm7462® Recombinant peptide derivative of SBm7462® | Holstein Friesian (Bos taurus taurus) | Saponin | Dosage: -2 mg Regime: -Three vaccinations at weeks 0, 4, and 8 | Total efficacy: 72.4% | Serum antibody levels: -IgG mean OD value 1 week after 3rd injection (dilution 1: 100): 2.3 ± 0.5). Cytokine gene expression: -Upregulated: IL-4, INF-ƴ, IL-10, and IL-12. -Down-regulated: TNF-α. Histology of LN tissue: -Increased GCs in B-cell follicles. -Hyperplasia of medullary cords. | Patent: [85,86] Literature: [87] |
| Combination Vaccines with Bm86 | ||||||
| Bm95-MSP1a Recombinant Bm95 + recombinant Anaplasma marginale MSP1a | Beefmaster (Bos taurus indicus) × Charolais (Bos taurus taurus) | Montanide ISA 50V2 (Seppic, France) | Dosage: -120 μg Regime: -Three vaccinations at weeks 0, 4, and 8 | Total efficacy: 64% | Serum antibody levels: IgG mean OD value 2 weeks after 3rd injection (dilution 1: 100): 0.5 ± 0.1. | Literature: [88] |
| Bm86 + SUB Recombinant Bm86 and SUB (salivary gland) proteins | Hereford/Holstein (Bos taurus taurus) mixed breed | Water-in-oil emulsion: Saponin QuilA (water) in Montanide ISA 50 V2 (Seppic, France) (oil) | Dosage: -100 μg Regime: -Three vaccinations at weeks 0, 3, and 6 | REF: 97% | Serum antibody levels: Mean IgG titre in 2Log 3 weeks after 3rd injection (dilution n.i.): Bm86 + SUB injected separately: -Bm86: 10.2. -SUB: <7. Bm86 + SUB dual vaccine: -Bm86: 10.6. -SUB: 10.0. | Patent: [89] |
| pP0-Bm86 Synthetic peptide derivative of p0 ribosomal protein conjugated to recombinant Bm86 | Cuban Siboney: 5/8 Holstein (Bos taurus taurus) 3/8 Zebu (Bos taurus indicus) | Montanide ISA 50V (Seppic, France) | Dosage: -500 μg Regime: -Three vaccinations at weeks 0, 3, and 5 | Total efficacy: 84% | Serum antibody levels: IgG mean OD value 2 weeks after 3rd injection (dilution n.i.): -anti-pP0: ± 10,000. -anti-Bm86: ± 8000. | Literature: [90] |
| pSUB + Bm86 SUB synthetic peptide derivative and recombinant Bm86 | Bos taurus × Bos indicus and 5/8 Holstein | Montanide ISA 50 V (Seppic, France) | Dosage: -100 μg Regime: -Three vaccinations at weeks 0, 4, and 9 | Total efficacy: Peptide SUB alone: 67% Bm86 alone: 56% Peptide SUB + Bm86: 49% | Serum antibody levels: IgG mean OD value 2 weeks after 3rd injection (dilution 1: 100): -anti-pSUB alone: 1.9 ± 0.1. -anti-Bm86 alone: 1.1 ± 0.1. -anti-pSUB dual: 1.7 ± 0.1. -anti-Bm86 dual: 1.4 ± 0.1. | Literature: [91] |
| Bm86 + H. anatolicum SUB and tropomyosin (TPM) | Bos taurus × Bos indicus | Montanide ISA 50 V2 (Seppic, France) | Dosage: -100 μg Regime: -Three vaccinations at weeks 0, 4, and 8 | Total efficacy: 87.6% | Serum antibody levels: Anti-rBm86 OD values across 6 bovines 2 weeks after 3rd injection (dilution 1:16,000): -IgG: 0.68–0.75 (IgG titre 90 DPI: 1:102,400). -IgG1: 0.68–0.75. -IgG2: 0.25–0.3. Anti-rSUB OD values across 6 bovines 2 weeks after 3rd injection (dilution 1:16,000): -IgG: 0.65–0.85 (IgG titre 75 DPI: 1:64,000). -IgG1: 0.68–0.80. -IgG2: 0.36–0.68. Anti-rTPM OD values across 6 bovines 2 weeks after 3rd injection (dilution 1:8000) -IgG: 0.6–0.72 (IgG titre 90 DPI: 1:51,200) -IgG1: 0.62–0.8. -IgG2: 0.28–0.75. | Literature: [92] |
4. First Encounter with the Antigen Diversity-Driven Bottleneck (2000–2020): Geographical Variation, Cross-Species Protection, and Attempts to Improve Bm86-Based Vaccines
| Vaccine and Major Antigen(s) | Breed | Adjuvant | Vaccination Regime | Tick Species | Efficacy in Bovine Vaccination Trials | Immune Markers | Reference(s) |
|---|---|---|---|---|---|---|---|
| Cross-Species Testing of Bm86, Bm86 Homolog and Bm86 Ortholog Vaccines | |||||||
| Bm86 Tick-GARD® and TickGARD® Plus Discontinued commercial vaccines (Intervet Pty. Ltd., Bendigo, Australia.) | 1. Holstein Friesian (Bos taurus taurus) [97] 2. N.i. [98] | Montanide ISA 50V (Seppic, France) | Dosage: -25 μg [97,98] Regime: -Two vaccinations at weeks 0 and 4 [97,98] | 1. R. annulatus [97] 2A. R. microplus [98] 2B. R. decoloratus [98] 2C. R. appendiculatus [98] 2D. H. a. anatolicum [98] 2E. H. dromedarii [98] 2F. A. variegatum [98] | Total efficacy: 1. 100% [97] 2A. 74% [98] 2B. 70% [98] 2C. 0% [98] 2F. 0% [98] Reduction in tick parameters: 2D. 50% reduction in the total weight of engorging nymphs. [98] 2E. 95% reduction in number of nymphs; 55% reduction in weight of surviving ticks. [98] | Serum antibody levels: 1. Mean IgG titre after 2nd injection: [97] -30 days 1:1600–1:24,000. -90 days: 1:6400–1:24,000. -138 days: 1:6400. -160 days: 1:400 (after repeated infestation). -270 days: 1:400 (after repeated infestation). -390 days: 1:400–1600 (after repeated infestation). 2. Mean IgG titre after 2nd injection (dilution 1:500): n.i. [98] | Patents: [75,76] Literature: [97,98] |
| Bm86 Gavac® Commercially available vaccine (Heber Biotec S.A., Cuba) | 1. Aberdeen Angus and cross-breed (n.i.) [94] 2. Crossbreed (n.i.) [77] 3. Bos taurus X B. indicus [99] 4. Holstein Friesian (Bos taurus taurus) [100] | Montanide ISA 50V (Seppic, France) | Dosage: -100 μg [77,94,99,100] Regime: -Three vaccinations at weeks 1, 3, and 7 [71] -Three vaccinations at weeks 0, 4, and 7 [88,96,98] | 1. R. annulatus [94] 2A. R. annulatus [77] 2B. R. microplus [77] 3A. H. a. anatolicum larvae [99] 3B. H. a. anatolicum unfed adults [99] 3C. R. microplus larvae [99] 4A. H. dromedarii [100] 4B. A. cajennense [100] | Total efficacy: 1. >99.9% [94] 2A. 99.6% [77] 2B. 85.2% [77] 3A. 26.8% [99] 3B. 25.1% [99] 3C. 44.5% [99] 4B. 0% [100] Reduction in tick parameters: 4A. 89% reduction in number of engorging nymphs; 32% reduction in weight of surviving ticks. [100] | Serum antibody levels: 1. N.i. [100] 2. Bm86 mean IgG OD values 2 weeks after 3rd injection (dilution 1: 1000): ±0.2–0.3. [77] 3. Bm86 mean IgG OD values two weeks after 3rd injection (dilution 1: 500): n.i. [99] 4. Bm86 mean IgG OD values two weeks after 3rd injection (dilution n.i.): n.i. [100] | Patent: [78] Literature: [77,94,99,100] |
| Ba86 Bm86 orthologue of R. annulatus | Crossbreed (n.i.) | Montanide ISA 50V (Seppic, France) | Dosage: -100 μg Regime: -Three vaccinations at weeks 1, 3, and 7 | A. R. annulatus B. R. microplus | Total efficacy: A. 83% B. 71.5% | Serum antibody levels: Bm86 mean IgG OD values 2 weeks after 3rd injection (dilution 1: 1000): ± 1.22. | Literature: [77] |
| Bm86 Mozambique strain | 1. Crossbreed (n.i.) [77] 2. Holstein Friesian (Bos taurus taurus) [101] | 1. Montanide ISA 50V [77] (Seppic, France) 2. Montanide 888 (50%) [101] | Dosage: -100 μg [71,102] Regime: -Three vaccinations at weeks 1, 3, and 7 [71] -Three vaccinations at weeks 0, 4, and 7 [102] | 1A. R. annulatus [77] 1B. R. microplus [77] 2A. H. scupense [101] 2B. H. excavatum [101] | Total efficacy: 1A. 99.6% [77] 1B. 70.4% [77] 2A. 0% [101] 2B. 0% [101] | Serum antibody levels: 1. Anti-Bm86 mean IgG OD values 2 weeks after 3rd injection (dilution 1: 1000): ±1.18 [77] 2. Anti-Bm86 mean IgG OD values in Bm86-vaccinated cattle (dilution 1:1000): [101] -2 weeks after 2nd injection: 1.2 ± 0.1. -2 weeks after 3rd injection: 1.3 ± 0.1. 2. Anti-Hd86 mean IgG OD values in Bm86-vaccinated cattle (1:1000): [101] -2 weeks after 2nd injection: 1.4 ± 0.2. -2 weeks after 3rd injection: 1.6 ± 0.1. | Literature: [77,101] |
| rHaa86 Bm86 orthologue of H. a. anatolicum | Bos taurus X B. indicus | 1. Saponin in mineral oil [102] 2. 10% Montanide 888 in mineral oil [99] | Dosage: -100 μg [96] -125 μg [103] Regime: -Three vaccinations at weeks 0, 4, and 7 [96,103] | 1. H. a. anatolicum unfed adults (1st) and larvae (2nd) [102] 2A. H. a. anatolicum larvae [99] 2B. H. a. anatolicum unfed adults [99] 2C. R. microplus larvae [99] | Total efficacy: 1. 61.6% [102] 2A. 68.7% [99] 2B. 45.8% [99] 2C. 36.5% [99] | Serum antibody levels: 1. Anti-rHaa86 mean IgG OD values two weeks after 3rd injection (dilution 1:50): [103] 1.4 ± 0.3. 2. Anti-rHaa86 mean IgG OD values two weeks after 3rd injection (dilution 1: 500): [100] -IgG ± 0.7. -IgG1 ± 0.65. -IgG2 ± 0.3. | Literature: [99,102] |
| Ra86 Bm86 ortholog of Rhipicephalus appendiculatus | Holstein Friesian (Bos taurus taurus) | Montanide ISA 50V (Seppic, France) | Dosage: -50 μg Regime: -Three vaccinations at weeks 0, 5, and 10 | R. appendiculatus | Reduction in tick parameters: No significant effect on adult engorgement weight, adult mortality, or nymphal mortality. Significant reduction in nymphal moulting and egg viability. | Serum antibody levels: Anti-Ra86 mean IgG OD values 2 weeks after 3rd injection (dilution 1:1000): 0.7 ± 0.1. | Literature: [103] |
| Hd86 Bm86 orthologue of Hyalomma scupense | Holstein Friesian (Bos taurus taurus) | Montanide 888 (50%) | Dosage: -100 μg Regime: -Three vaccinations at weeks 0, 4, and 7 | A. H. scupense B. H. excavatum | Total efficacy: B. 0% Reduction in tick parameters: A. 59.19% reduction in number of engorging nymphs; 0% efficacy against adult tick infestations. | Serum antibody levels: Anti-Hd86 mean IgG OD values in Hd86 vaccinated cattle (dilution 1:1000): -2 weeks after 2nd injection: 1.25 ± 0.5. -2 weeks after 3rd injection: 1.75 ± 0.1. Anti-Bm86 mean IgG OD values in Hd86 vaccinated cattle (dilution 1:1000): -2 weeks after 2nd injection: 1.1 ± 0.1. -2 weeks after 3rd injection: 1.3 ± 0.1. | Literature: [101] |
| ATAQ peptide- KLH conjugate Bm86 homolog | Holstein Friesian (Bos taurus taurus) | Montanide ISA 61 VG (Seppic®, Paris) | Dosage: -200 μg Regime: -Three vaccinations at weeks 0, 2, and 4 | R. microplus | Total efficacy: 35% | Serum antibody levels: ATAQ peptide IgG mean OD values 3 weeks after 3rd injection (dilution 1: 300): 0.7 ± 0.3. | Literature: [104] |
| Rhipicephalus microplus Bm86 + H. anatolicum Subolesin (SUB) and tropomyosin (TPM) | Bos taurus × Bos indicus | Montanide ISA 50 V2 (Seppic, France) | Dosage: -100 μg Regime: -Three vaccinations at weeks 0, 4, and 8 | A. R. microplus B. H. anatolicum | Total efficacy: A. 87.6% B. 87.2% against larvae B. 86.2% against adults | Serum antibody levels: OD values across 6 bovines 2 weeks after 3rd injection for rBm86 (dilution 1:16,000) -IgG: 0.68–0.75 (IgG titre 90 DPI: 1:102,400). -IgG1: 0.68–0.75. -IgG2: 0.25–0.3. OD values across 6 bovines 2 weeks after 3rd injection for rSUB (dilution 1:16,000) -IgG: 0.65–0.85 (IgG titre 75 DPI: 1:64,000). -IgG1: 0.68–0.80. -IgG2: 0.36–0.68. OD values across 6 bovines 2 weeks after 3rd injection for rTPM (dilution 1:8000) -IgG: 0.6–0.72 (IgG titre 90 DPI: 1:51,200). -IgG1: 0.62–0.8. -IgG2: 0.28–0.75. | Literature: [92] |
5. The Next Generation of Biotechnology and Tick Vaccines (2000–Present): New Advances in Tick Vaccine Development and Design
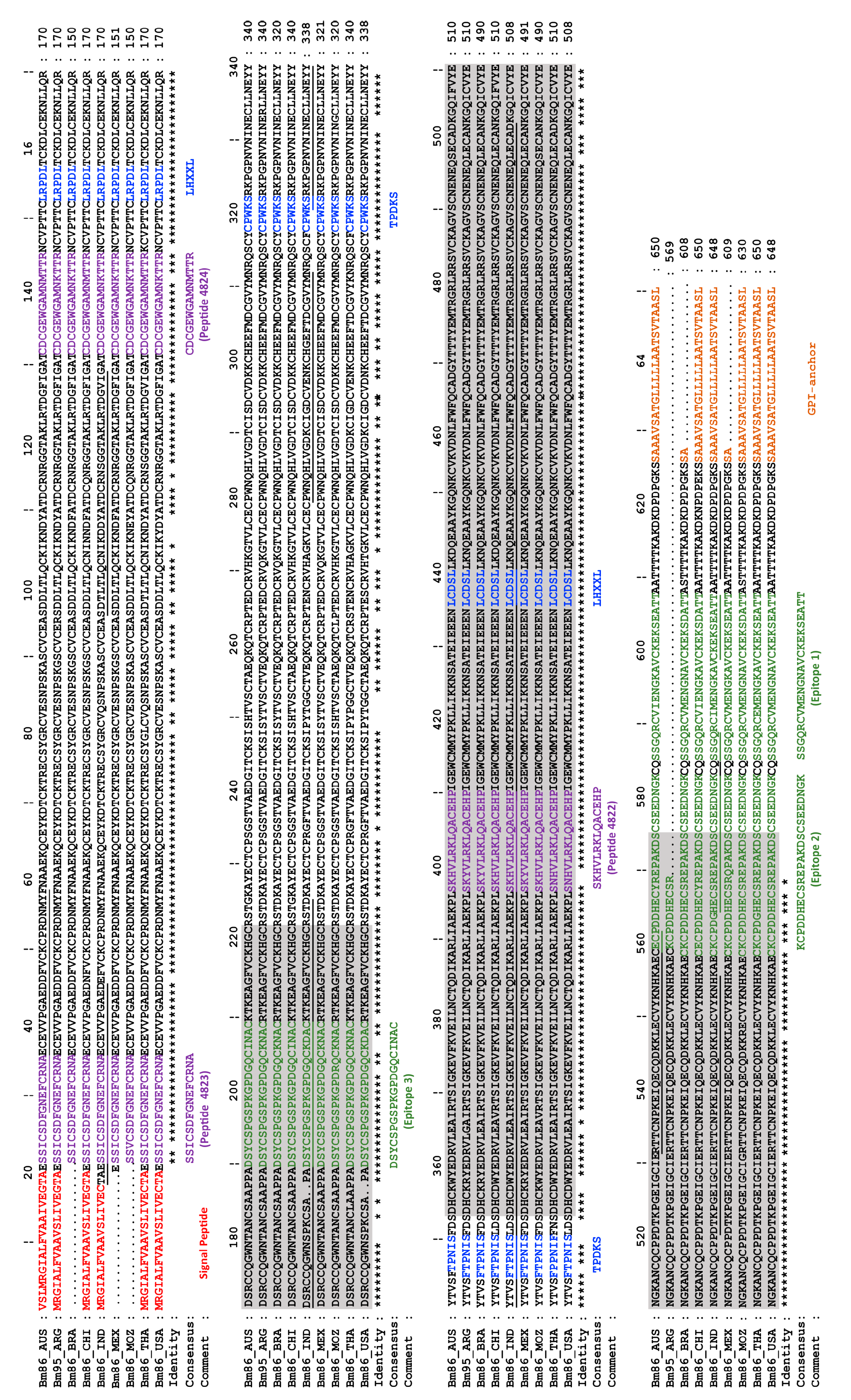
6. Advances in Bm86 Epitope Mapping—Entering the Era of Peptide Vaccines
7. Current Bm86-Based Commercial Vaccines and the Next Generation of Bm86-Based Cocktail Vaccines
8. Bm86-Based Vaccines: What Are the Current Gaps and Future Research Needed?
8.1. Antigen Stability and Adjuvants
8.2. Understanding the Immune Responses That Confer Protection across Genetically Diverse Bovine Breeds: The Need to Identity Shared Elements for Targeted Intervention
8.3. Impact of Field Conditions: Co-Infestation and Co-Infection
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Arthur, D.R. Ticks in Egypt in 1500 B.C.? Nature 1965, 206, 1060–1061. [Google Scholar] [CrossRef] [PubMed]
- Huchet, J.B.; Callou, C.; Lichtenberg, R.; Dunand, F. The dog mummy, the ticks and the louse fly: Archaeological report of severe ectoparasitosis in Ancient Egypt. Int. J. Paleopathol. 2013, 3, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Sonenshine, D.E.; Roe, R.M. Biology of Ticks Volume 1. Oxford University Press: Cary, NC, USA, 2013. [Google Scholar]
- Hurtado, O.J.B.; Giraldo-Ríos, C. Economic and health impact of the ticks in production animals. Ticks Tick-Borne Pathog. 2018, 9, 1–19. [Google Scholar]
- Lew-Tabor, A.E.; Rodriguez Valle, M. A review of reverse vaccinology approaches for the development of vaccines against ticks and tick borne diseases. Ticks Tick. Borne Dis. 2016, 7, 573–585. [Google Scholar] [CrossRef]
- Obaid, M.K.; Islam, N.; Alouffi, A.; Khan, A.Z.; da Silva Vaz, I., Jr.; Tanaka, T.; Ali, A. Acaricides Resistance in Ticks: Selection, Diagnosis, Mechanisms, and Mitigation. Front. Cell Infect. Microbiol. 2022, 12, 941831. [Google Scholar] [CrossRef]
- Borges, L.M.F.; Sousa, L.A.D.d.; Barbosa, C.d.S. Perspectives for the use of plant extracts to control the cattle tick Rhipicephalus (Boophilus) microplus. Rev. Bras. Parasitol. Veterinária 2011, 20, 89–96. [Google Scholar] [CrossRef]
- Mwangi, E.N.; Dipeolu, O.O.; Newson, R.M.; Kaaya, G.P.; Hassan, S.M. Predators, parasitoids and pathogens of ticks: A review. Biocontrol Sci. Technol. 1991, 1, 147–156. [Google Scholar] [CrossRef]
- Hajek, A.E.; Delalibera, I. Fungal pathogens as classical biological control agents against arthropods. BioControl 2010, 55, 147–158. [Google Scholar] [CrossRef]
- Brossard, M. The use of vaccines and genetically resistant animals in tick control. Rev. Sci. Tech. 1998, 17, 188–199. [Google Scholar] [CrossRef]
- Frisch, J. Identification of a major gene for resistance to cattle ticks. In Proceedings of the 5th World Congress on Genetics Applied to Livestock Production, Guelph, ON, Canada, 7–12 August 1994; Volume 20, pp. 293–295. [Google Scholar]
- Cardoso, F.F.; Matika, O.; Djikeng, A.; Mapholi, N.; Burrow, H.M.; Yokoo, M.J.I.; Campos, G.S.; Gulias-Gomes, C.C.; Riggio, V.; Pong-Wong, R.; et al. Multiple Country and Breed Genomic Prediction of Tick Resistance in Beef Cattle. Front. Immunol. 2021, 12, 620847. [Google Scholar] [CrossRef]
- Pereira, D.F.S.; Ribeiro, H.S.; Goncalves, A.A.M.; da Silva, A.V.; Lair, D.F.; de Oliveira, D.S.; Boas, D.F.V.; Conrado, I.; Leite, J.C.; Barata, L.M.; et al. Rhipicephalus microplus: An overview of vaccine antigens against the cattle tick. Ticks Tick. Borne Dis. 2022, 13, 101828. [Google Scholar] [CrossRef]
- Antunes, S.; Domingos, A. Tick Vaccines and Concealed versus Exposed Antigens. Pathogens 2023, 12, 374. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.N.; Jmel, M.A.; Mekki, I.; Dijkgraaf, I.; Kotsyfakis, M. Recent Advances in Tick Antigen Discovery and Anti-Tick Vaccine Development. Int. J. Mol. Sci. 2023, 24, 4969. [Google Scholar] [CrossRef] [PubMed]
- Pasteur, M. An address on vaccination in relation to chicken cholera and splenic fever. Br. Med. J. 1881, 2, 283. [Google Scholar] [CrossRef] [PubMed]
- Hilleman, M.R. Yeast recombinant hepatitis B vaccine. Infection 1987, 15, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Tabor, A.E. A Review of Australian Tick Vaccine Research. Vaccines 2021, 9, 1030. [Google Scholar] [CrossRef]
- Rand, K.N.; Moore, T.; Sriskantha, A.; Spring, K.; Tellam, R.; Willadsen, P.; Cobon, G.S. Cloning and expression of a protective antigen from the cattle tick Boophilus microplus. Proc. Natl. Acad. Sci. USA 1989, 86, 9657–9661. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Venzal, J.M.; Nava, S.; Mangold, A.; Guglielmone, A.A.; Labruna, M.B.; De La Fuente, J. Reinstatement of Rhipicephalus (Boophilus) australis (Acari: Ixodidae) with redescription of the adult and larval stages. J. Med. Entomol. 2012, 49, 794–802. [Google Scholar] [CrossRef]
- Johnston, T.H.; Bancroft, M.J. Tick-resistance of Cattle. Agric. Gaz. New South. Wales 1918, 29, 319–320. [Google Scholar]
- Johnston, T.H.; Bancroft, M.J. A tick resistant condition in cattle. Proc. R. Soc. Qld. 1918, 30, 219–317. [Google Scholar] [CrossRef]
- Strother, G.R.; Burns, E.C.; Smart, L.I. Resistance of purebred Brahman, Hereford, and Brahman times Hereford crossbred cattle to the lone star tick, Amblyomma americanum (Acarina: Ixodidae). J. Med. Entomol. 1974, 11, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Doube, B.M. Cattle and the paralysis tick Ixodes holocyclus. Aust. Vet. J. 1975, 51, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Wagland, B.M.; Roberts, J.A.; Sutherst, R.W. Growth of Haemaphysalis (Kaiseriana) longicornis on cattle. Int. J. Parasitol. 1979, 9, 177–182. [Google Scholar] [CrossRef]
- Kotal, J.; Langhansova, H.; Lieskovska, J.; Andersen, J.F.; Francischetti, I.M.; Chavakis, T.; Kopecky, J.; Pedra, J.H.; Kotsyfakis, M.; Chmelar, J. Modulation of host immunity by tick saliva. J. Proteom. 2015, 128, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Aounallah, H.; Bensaoud, C.; M’Ghirbi, Y.; Faria, F.; Chmelar, J.I.; Kotsyfakis, M. Tick Salivary Compounds for Targeted Immunomodulatory Therapy. Front. Immunol. 2020, 11, 583845. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, S.; Miyake, K.; Kamiya, A.; Karasuyama, H. The role of basophils in acquired protective immunity to tick infestation. Parasite Immunol. 2021, 43, e12804. [Google Scholar] [CrossRef]
- Roberts, J.A.; Kerr, J.D. Boophilus microplus: Passive transfer of resistance in cattle. J. Parasitol. 1976, 62, 485–488. [Google Scholar] [CrossRef]
- Mapholi, N.O.; Marufu, M.C.; Maiwashe, A.; Banga, C.B.; Muchenje, V.; MacNeil, M.D.; Chimonyo, M.; Dzama, K. Towards a genomics approach to tick (Acari: Ixodidae) control in cattle: A review. Ticks Tick. Borne Dis. 2014, 5, 475–483. [Google Scholar] [CrossRef]
- Robbertse, L.; Richards, S.A.; Maritz-Olivier, C. Bovine Immune Factors Underlying Tick Resistance: Integration and Future Directions. Front. Cell Infect. Microbiol. 2017, 7, 522. [Google Scholar] [CrossRef]
- Tabor, A.E.; Ali, A.; Rehman, G.; Rocha Garcia, G.; Zangirolamo, A.F.; Malardo, T.; Jonsson, N.N. Cattle tick Rhipicephalus microplus-host interface: A review of resistant and susceptible host responses. Front. Cell. Infect. Microbiol. 2017, 7, 506. [Google Scholar] [CrossRef]
- O’Neill, C.J.; Swain, D.L.; Kadarmideen, H.N. Evolutionary process of Bos taurus cattle in favourable versus unfavourable environments and its implications for genetic selection. Evol. Appl. 2010, 3, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Stutzer, C.; Richards, S.A.; Ferreira, M.; Baron, S.; Maritz-Olivier, C. Metazoan Parasite Vaccines: Present Status and Future Prospects. Front. Cell Infect. Microbiol. 2018, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Stafford III, K.C.; Williams, S.C.; Molaei, G. Integrated pest management in controlling ticks and tick-associated diseases. J. Integr. Pest. Manag. 2017, 8, 28. [Google Scholar] [CrossRef]
- Trager, W. Acquired Immunity to Ticks. J. Parasitol. 1939, 25, 57–81. [Google Scholar] [CrossRef]
- Trager, W. Further observations on acquired immunity to the tick Dermacentor variabilis Say. J. Parasitol. 1939, 25, 137–139. [Google Scholar] [CrossRef]
- Roberts, J. Acquisition by the host of resistance to the cattle tick, Boophilus microplus (Canestrini). J. Parasitol. 1968, 54, 657–662. [Google Scholar] [CrossRef]
- Galun, R. Research into Alternative Arthropod Control Measures against livestock Pests. I. Proceedings of the Workshop on the Ecology and Control of External Parasites of Economic Importance on Bovines in Latin America, 25–30 August 1975; Kenneth, C.T., Ed.; Centro International de Agricultura Tropical: Cali, Colombia, 1978. [Google Scholar]
- Allen, J.R.; Humphreys, S.J. Immunisation of guinea pigs and cattle against ticks. Nature 1979, 280, 491–493. [Google Scholar] [CrossRef]
- Almazán, C. Impact of the Paper by Allen and Humphreys (1979) on Anti-Tick Vaccine Research. Pathogens 2022, 11, 1253. [Google Scholar] [CrossRef]
- Agbede, R.; Kemp, D. Immunization of cattle against Boophilus microplus using extracts derived from adult female ticks: Histopathology of ticks feeding on vaccinated cattle. Int. J. Parasitol. 1986, 16, 35–41. [Google Scholar] [CrossRef]
- Johnston, L.; Kemp, D.; Pearson, R. Immunization of cattle against Boophilus microplus using extracts derived from adult female ticks: Effects of induced immunity on tick populations. Int. J. Parasitol. 1986, 16, 27–34. [Google Scholar] [CrossRef]
- Kemp, D.; Pearson, R.; Gough, J.; Willadsen, P. Vaccination against Boophilus microplus: Localization of antigens on tick gut cells and their interaction with the host immune system. Exp. Appl. Acarol. 1989, 7, 43–58. [Google Scholar] [PubMed]
- Willadsen, P.; Riding, G.A.; McKenna, R.V.; Kemp, D.H.; Tellam, R.L.; Nielsen, J.N.; Lahnstein, J.; Cobon, G.S.; Gough, J.M. Immunologic control of a parasitic arthropod. Identification of a protective antigen from Boophilus microplus. J. Immunol. 1989, 143, 1346–1351. [Google Scholar] [CrossRef] [PubMed]
- Willadsen, P.; McKenna, R.V.; Riding, G.A. Isolation from the cattle tick, Boophilus microplus, of antigenic material capable of eliciting a protective immunological response in the bovine host. Int. J. Parasitol. 1988, 18, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Willadsen, P. The Development of a New or Improved Vaccine against Boophilus microplus: Opportunities for R&D Investment; Meat & Livestock Australia Limited: Sydney, Australia, 2008. [Google Scholar]
- Willadsen, P.; Kemp, D. Vaccination with ‘concealed’antigens for tick control. Parasitol. Today 1988, 4, 196–198. [Google Scholar] [PubMed]
- Tellam, R.L.; Smith, D.; Kemp, D.H.; Willadsen, P. Vaccination against ticks. In Animal Parasite Control Utilizing Biotechnology; Wong, W.K., Ed.; CRC Press, Inc.: Boca Raton, FL, USA, 1992; Volume 1, pp. 303–331. [Google Scholar]
- Gough, J.M.; Kemp, D.H. Localization of a low abundance membrane protein (Bm86) on the gut cells of the cattle tick Boophilus microplus by immunogold labeling. J. Parasitol. 1993, 79, 900–907. [Google Scholar] [CrossRef]
- Willadsen, P.; McKenna, R. Vaccination with ‘concealed’antigens: Myth or reality? Parasite Immunol. 1991, 13, 605–616. [Google Scholar]
- Turnbull, I.; Smith, D.; Sharp, P.; Cobon, G.; Hynes, M. Expression and secretion in Aspergillus nidulans and Aspergillus niger of a cell surface glycoprotein from the cattle tick, Boophilus microplus, by using the fungal amdS promoter system. Appl. Environ. Microbiol. 1990, 56, 2847–2852. [Google Scholar]
- Richardson, M.; Smith, D.; Kemp, D.; Tellam, R. Native and baculovirus-expressed forms of the immunoprotective protein BM86 from Boophilus microplus are anchored to the cell membrane by a glycosylphosphatidyl inositol linkage. Insect Mol. Biol. 1993, 1, 139–147. [Google Scholar] [CrossRef]
- Willadsen, P.; Bird, P.; Cobon, G.S.; Hungerford, J. Commercialisation of a recombinant vaccine against Boophilus microplus. Parasitology 1995, 110, S43–S50. [Google Scholar] [CrossRef]
- Rodriguez, M.; Penichet, M.L.; Mouris, A.E.; Labarta, V.; Luaces, L.L.; Rubiera, R.; Cordoves, C.; Sanchez, P.A.; Ramos, E.; Soto, A.; et al. Control of Boophilus microplus populations in grazing cattle vaccinated with a recombinant Bm86 antigen preparation. Vet. Parasitol. 1995, 57, 339–349. [Google Scholar] [CrossRef]
- Rodriguez, M.; Massard, C.L.; da Fonseca, A.H.; Ramos, N.F.; Machado, H.; Labarta, V.; de la Fuente, J. Effect of vaccination with a recombinant Bm86 antigen preparation on natural infestations of Boophilus microplus in grazing dairy and beef pure and cross-bred cattle in Brazil. Vaccine 1995, 13, 1804–1808. [Google Scholar] [CrossRef] [PubMed]
- Willadsen, P. Novel vaccines for ectoparasites. Vet. Parasitol. 1997, 71, 209–222. [Google Scholar] [CrossRef]
- Rodriguez, M.; Rubiera, R.; Penichet, M.; Montesinos, R.; Cremata, J.; Falcon, V.; Sanchez, G.; Bringas, R.; Cordoves, C.; Valdes, M.; et al. High level expression of the B. microplus Bm86 antigen in the yeast Pichia pastoris forming highly immunogenic particles for cattle. J. Biotechnol. 1994, 33, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Canales, M.; Enriquez, A.; Ramos, E.; Cabrera, D.; Dandie, H.; Soto, A.; Falcon, V.; Rodriguez, M.; de la Fuente, J. Large-scale production in Pichia pastoris of the recombinant vaccine Gavac against cattle tick. Vaccine 1997, 15, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, N.N.; Matschoss, A.L.; Pepper, P.; Green, P.E.; Albrecht, M.S.; Hungerford, J.; Ansell, J. Evaluation of tickGARD(PLUS), a novel vaccine against Boophilus microplus, in lactating Holstein-Friesian cows. Vet. Parasitol. 2000, 88, 275–285. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente, J.; Almazan, C.; Canales, M.; Perez de la Lastra, J.M.; Kocan, K.M.; Willadsen, P. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim. Health Res. Rev. 2007, 8, 23–28. [Google Scholar] [CrossRef]
- Willadsen, P.; Smith, D.; Cobon, G.; McKenna, R.V. Comparative vaccination of cattle against Boophilus microplus with recombinant antigen Bm86 alone or in combination with recombinant Bm91. Parasite Immunol. 1996, 18, 241–246. [Google Scholar] [CrossRef]
- Jarmey, J.; Riding, G.; Pearson, R.; McKenna, R.; Willadsen, P. Carboxydipeptidase from Boophilus microplus: A “concealed” antigen with similarity to angiotensin-converting enzyme. Insect Biochem. Mol. Biol. 1995, 25, 969–974. [Google Scholar]
- Riding, G.A.; Jarmey, J.; McKenna, R.V.; Pearson, R.; Cobon, G.S.; Willadsen, P. A protective" concealed" antigen from Boophilus microplus. Purification, localization, and possible function. J. Immunol. 1994, 153, 5158–5166. [Google Scholar] [CrossRef]
- Lambertz, C.; Chongkasikit, N.; Jittapalapong, S.; Gauly, M. Immune response of Bos indicus cattle against the anti-tick antigen Bm91 derived from local Rhipicephalus (Boophilus) microplus ticks and its effect on tick reproduction under natural infestation. J. Parasitol. Res. 2012, 2012, 907607. [Google Scholar] [CrossRef]
- De La Fuente, J.; Rodríguez, M.; Redondo, M.; Montero, C.; García-García, J.; Méndez, L.; Serrano, E.; Valdés, M.; Enriquez, A.; Canales, M. Field studies and cost-effectiveness analysis of vaccination with Gavac™ against the cattle tick Boophilus microplus. Vaccine 1998, 16, 366–373. [Google Scholar]
- Vanegas, L.; Parra, S.; Vanegas, C.; de la Fuente, J. Recombinant Vaccines for the Control of Cattle Tick. Elfos Sci. 1995, 196–199. [Google Scholar]
- de la Fuente, J.; Rodriguez, M.; Montero, C.; Redondo, M.; Garcia-Garcia, J.C.; Mendez, L.; Serrano, E.; Valdes, M.; Enriquez, A.; Canales, M.; et al. Vaccination against ticks (Boophilus spp.): The experience with the Bm86-based vaccine Gavac. Genet. Anal. 1999, 15, 143–148. [Google Scholar] [CrossRef]
- Valle, M.R.; Mèndez, L.; Valdez, M.; Redondo, M.; Espinosa, C.M.; Vargas, M.; Cruz, R.L.; Barrios, H.P.; Seoane, G.; Ramirez, E.S. Integrated control of Boophilus microplus ticks in Cuba based on vaccination with the anti-tick vaccine Gavac TM. Exp. Appl. Acarol. 2004, 34, 375–382. [Google Scholar] [PubMed]
- Garcia-Garcia, J.C.; Montero, C.; Redondo, M.; Vargas, M.; Canales, M.; Boue, O.; Rodriguez, M.; Joglar, M.; Machado, H.; Gonzalez, I.L.; et al. Control of ticks resistant to immunization with Bm86 in cattle vaccinated with the recombinant antigen Bm95 isolated from the cattle tick, Boophilus microplus. Vaccine 2000, 18, 2275–2287. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.A.; Opdebeeck, J.P. Humoral immune responses of Hereford cattle vaccinated with midgut antigens of the cattle tick, Boophilus microplus. Parasite Immunol. 1990, 12, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Patarroyo, S.J.H.; Vargas, M.I.; Gonzalez, C.Z.; Guzman, F.; Martins-Filho, O.A.; Afonso, L.C.; Valente, F.L.; Peconick, A.P.; Marciano, A.P.; Patarroyo, A.M.; et al. Immune response of bovines stimulated by synthetic vaccine SBm7462 against Rhipicephalus (Boophilus) microplus. Vet. Parasitol. 2009, 166, 333–339. [Google Scholar] [CrossRef]
- Rodriguez Valle, M.; Montero, C.; Machado, H.; Joglar, M.; de la Fuente, J.; Garcia-Garcia, J.C. The evaluation of yeast derivatives as adjuvants for the immune response to the Bm86 antigen in cattle. BMC Biotechnol. 2001, 1, 2. [Google Scholar] [CrossRef]
- Willadsen, P.; Kemp, D.H. Novel vaccination for control of the Babesia vector, Boophilus microplus. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 107. [Google Scholar] [CrossRef]
- Cobon, G.S.; Moore, J.T.; Johnston, L.A.Y.; Willadsen, P.; Kemp, D.H.; Sriskantha, A.; Riding, G.A.; Rand, K.N. DNA Encoding a Cell Membrane Glycoproten of a Tick Gut. United States US5587311A, 4 October 1996. [Google Scholar]
- Cobon, G.S.; Moore, J.T.; Johnston, L.A.Y.; Willadsen, P.; Kemp, D.H.; Sriskantha, A.; Riding, G.A.; Rand, K.N.; Moore, J.T.; Johnston, L.A.Y.; et al. Vaccine Containing Tick Antigens. Australia WO1988003929A1, 2 June 1988. [Google Scholar]
- Canales, M.; Almazan, C.; Naranjo, V.; Jongejan, F.; de la Fuente, J. Vaccination with recombinant Boophilus annulatus Bm86 ortholog protein, Ba86, protects cattle against B. annulatus and B. microplus infestations. BMC Biotechnol. 2009, 9, 29. [Google Scholar] [CrossRef]
- Chaplen, R.R.; Rodriguez-Valle, M.; Segui, R.M.; Rodriguez, M.V.; Cespedes, C.O.C.; Fuentes, A.R.C.; Garcia, M.P.E.; Cama, V.F.; Padro, M.L.P.; Ricardo, B.P.; et al. Process for the Production of a Particulate Tick Antigen in Yeast Pichia Pastoris, Transformed Microorganism, Antigen and Vaccine Preparation. Brazil BR9300625A, 4 October 1993. [Google Scholar]
- Kumar, A.; Garg, R.; Yadav, C.L.; Vatsya, S.; Kumar, R.R.; Sugumar, P.; Chandran, D.; Mangamoorib, L.N.; Bedarkar, S.N. Immune responses against recombinant tick antigen, Bm95, for the control of Rhipicephalus (Boophilus) microplus ticks in cattle. Vet. Parasitol. 2009, 165, 119–124. [Google Scholar] [CrossRef]
- Vargas, M.; Montero, C.; Sanchez, D.; Perez, D.; Valdes, M.; Alfonso, A.; Joglar, M.; Machado, H.; Rodriguez, E.; Mendez, L.; et al. Two initial vaccinations with the Bm86-based Gavacplus vaccine against Rhipicephalus (Boophilus) microplus induce similar reproductive suppression to three initial vaccinations under production conditions. BMC Vet. Res. 2010, 6, 43. [Google Scholar] [CrossRef]
- Garcia-Garcia, J.C.; Garcia, J.D.L.; Rodriguez-Valle, M.; Espinosa, C.A.M. BM95 Antigen of the Cattle Tick Boophilus Microplus, Isolation, Sequence, Recombinant Molecules, Transformed Microorganisms and Vaccine Preparations. South Africa ZA991320B, 19 August 1999. [Google Scholar]
- Garcia-Garcia, J.C.; Espinosa, C.A.M.; Rodriguez-Valle, M.; Fuente, J.D.L. Vaccine Preparation Based on the Obtaining and Characterization of Boophilus Microplus Antigen Bm95. Cuba CU22694A1, 31 September 2001. [Google Scholar]
- Patarroyo, S.J.H.; Viloria, M.I.V.; Prates, A.A.; Mendes, M.A.D.; Guzman, F.; Portela, R.W.D.; Oliveira, R.D.C.; Murillo, M.E.P. Synthetic Vaccine for Tick Control. Brazil EP1289545B1, 26 January 2001. [Google Scholar]
- Patarroyo, S.J.H.; Portela, R.W.; De Castro, R.O.; Pimentel, J.C.; Guzman, F.; Patarroyo, M.E.; Vargas, M.I.; Prates, A.A.; Mendes, M.A. Immunization of cattle with synthetic peptides derived from the Boophilus microplus gut protein (Bm86). Vet. Immunol. Immunopathol. 2002, 88, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Patarroyo, S.J.H.; Viloria, M.I.V.; Sossal, S.; Araujo, L.S.d.; Gomez, G.A.T.; Mendes, M.A.D. Recombinant Peptide Vaccines against Ticks, and Nucleotide Sequences Coding for the Recombinant Peptides. Worldwide WO2015054764A1, 23 April 2015. [Google Scholar]
- Patarroyo, S.J.H.; Viloria, M.I.V.; Sossal, S.; Araujo, L.S.d.; Gomez, G.A.T.; Mendes, M.A.D. Recombinant Peptide Vaccines against Ticks, and Nucleotide Sequences Coding for the Recombinant Peptides. United States US10584151B2, 10 March 2020. [Google Scholar]
- Patarroyo, S.J.H.; de Sousa Neves, E.; Fidelis, C.F.; Tafur-Gomez, G.A.; de Araujo, L.; Vargas, M.I.; Sossai, S.; Prates-Patarroyo, P.A. Bovine immunisation with a recombinant peptide derived from synthetic SBm7462(R) (Bm86 epitope construct) immunogen for Rhipicephalus microplus control. Ticks Tick. Borne Dis. 2020, 11, 101461. [Google Scholar] [CrossRef]
- Almazan, C.; Moreno-Cantu, O.; Moreno-Cid, J.A.; Galindo, R.C.; Canales, M.; Villar, M.; de la Fuente, J. Control of tick infestations in cattle vaccinated with bacterial membranes containing surface-exposed tick protective antigens. Vaccine 2012, 30, 265–272. [Google Scholar] [CrossRef]
- Schetters, T.P.M.; Jansen, T. Vaccine against Rhipicephalus Ticks. Worldwide WO2014154847A1, 2 October 2014. [Google Scholar]
- Rodriguez Mallon, A.; Javier Gonzalez, L.; Encinosa Guzman, P.E.; Bechara, G.H.; Sanches, G.S.; Pousa, S.; Cabrera, G.; Cabrales, A.; Garay, H.; Mejias, R.; et al. Functional and Mass Spectrometric Evaluation of an Anti-Tick Antigen Based on the P0 Peptide Conjugated to Bm86 Protein. Pathogens 2020, 9, 513. [Google Scholar] [CrossRef]
- Mendoza-Martinez, N.; Alonso-Diaz, M.A.; Merino, O.; Fernandez-Salas, A.; Lagunes-Quintanilla, R. Protective efficacy of the peptide Subolesin antigen against the cattle tick Rhipicephalus microplus under natural infestation. Vet. Parasitol. 2021, 299, 109577. [Google Scholar] [CrossRef]
- Parthasarathi, B.C.; Kumar, B.; Bhure, S.K.; Sharma, A.K.; Manisha; Nagar, G.; Kumar, S.; Nandi, A.; Manjunathachar, H.V.; Chigure, G.M.; et al. Co-Immunization Efficacy of Recombinant Antigens against Rhipicephalus microplus and Hyalomma anatolicumTick Infestations. Pathogens 2023, 12, 433. [Google Scholar]
- Willadsen, P.; Jongejan, F. Immunology of the tick-host interaction and the control of ticks and tick-borne diseases. Parasitol. Today 1999, 15, 258–262. [Google Scholar] [CrossRef]
- Fragoso, H.; Rad, P.H.; Ortiz, M.; Rodriguez, M.; Redondo, M.; Herrera, L.; de la Fuente, J. Protection against Boophilus annulatus infestations in cattle vaccinated with the B. microplus Bm86-containing vaccine Gavac. off. Vaccine 1998, 16, 1990–1992. [Google Scholar] [CrossRef]
- Garcia-Garcia, J.C.; Gonzalez, I.L.; Gonzalez, D.M.; Valdes, M.; Mendez, L.; Lamberti, J.; D’Agostino, B.; Citroni, D.; Fragoso, H.; Ortiz, M.; et al. Sequence variations in the Boophilus microplus Bm86 locus and implications for immunoprotection in cattle vaccinated with this antigen. Exp. Appl. Acarol. 1999, 23, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Hüe, T.; Petermann, J.; Bonnefond, R.; Mermoud, I.; Rantoen, D.; Vuocolo, T. Experimental efficacy of a vaccine against Rhipicephalus australis. Exp. Appl. Acarol. 2017, 73, 245–256. [Google Scholar] [PubMed]
- Pipano, E.; Alekceev, E.; Galker, F.; Fish, L.; Samish, M.; Shkap, V. Immunity against Boophilus annulatus induced by the Bm86 (Tick-GARD) vaccine. Exp. Appl. Acarol. 2003, 29, 141–149. [Google Scholar] [CrossRef] [PubMed]
- De Vos, S.; Zeinstra, L.; Taoufik, O.; Willadsen, P.; Jongejan, F. Evidence for the utility of the Bm86 antigen from Boophilus microplus in vaccination against other tick species. Exp. Appl. Acarol. 2001, 25, 245–261. [Google Scholar] [CrossRef]
- Kumar, B.; Azhahianambi, P.; Ray, D.D.; Chaudhuri, P.; De La Fuente, J.; Kumar, R.; Ghosh, S. Comparative efficacy of rHaa86 and rBm86 against Hyalomma anatolicum anatolicum and Rhipicephalus (Boophilus) microplus. Parasite Immunol. 2012, 34, 297–301. [Google Scholar] [CrossRef]
- Rodriguez-Valle, M.; Taoufik, A.; Valdes, M.; Montero, C.; Ibrahin, H.; Hassan, S.M.; Jongejan, F.; de la Fuente, J. Efficacy of Rhipicephalus (Boophilus) microplus Bm86 against Hyalomma dromedarii and Amblyomma cajennense tick infestations in camels and cattle. Vaccine 2012, 30, 3453–3458. [Google Scholar] [CrossRef]
- Galai, Y.; Canales, M.; Ben Said, M.; Gharbi, M.; Mhadhbi, M.; Jedidi, M.; de La Fuente, J.; Darghouth, M.A. Efficacy of Hyalomma scupense (Hd86) antigen against Hyalomma excavatum and H. scupense tick infestations in cattle. Vaccine 2012, 30, 7084–7089. [Google Scholar] [CrossRef]
- Azhahianambi, P.; Ray, D.D.; Chaudhuri, P.; Gupta, R.; Ghosh, S. Vaccine Efficacy of Bm86 Ortholog of H. a. anatolicum, rHaa86 Expressed in Prokaryotic Expression System. J. Parasitol. Res. 2009, 2009, 165812. [Google Scholar] [CrossRef]
- Olds, C.; Mwaura, S.; Crowder, D.; Odongo, D.; van Oers, M.; Owen, J.; Bishop, R.; Daubenberger, C. Immunization of cattle with Ra86 impedes Rhipicephalus appendiculatus nymphal-to-adult molting. Ticks Tick. Borne Dis. 2012, 3, 170–178. [Google Scholar] [CrossRef]
- Aguirre Ade, A.; Lobo, F.P.; Cunha, R.C.; Garcia, M.V.; Andreotti, R. Design of the ATAQ peptide and its evaluation as an immunogen to develop a Rhipicephalus vaccine. Vet. Parasitol. 2016, 221, 30–38. [Google Scholar] [CrossRef]
- Odongo, D.; Kamau, L.; Skilton, R.; Mwaura, S.; Nitsch, C.; Musoke, A.; Taracha, E.; Daubenberger, C.; Bishop, R. Vaccination of cattle with TickGARD induces cross-reactive antibodies binding to conserved linear peptides of Bm86 homologues in Boophilus decoloratus. Vaccine 2007, 25, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Blecha, I.M.Z.; Csordas, B.G.; Aguirre, A.A.R.; Cunha, R.C.; Garcia, M.V.; Andreotti, R. Analysis of Bm86 conserved epitopes: Is a global vaccine against Cattle Tick Rhipicephalus microplus possible? Rev. Bras. Parasitol. Vet. 2018, 27, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathi, B.C.; Kumar, B.; Nagar, G.; Manjunathachar, H.V.; de la Fuente, J.; Ghosh, S. Analysis of Genetic Diversity in Indian Isolates of Rhipicephalus microplus Based on Bm86 Gene Sequence. Vaccines 2021, 9, 194. [Google Scholar] [CrossRef] [PubMed]
- Prudencio, C.R.; Rodrigues, A.A.R.; Cardoso, R.; Szabó, M.P.J.; Goulart, L.R. Antigen Fingerprinting of Polyclonal Antibodies raised in Immunized chickens with tick total Proteins: A reservoir for the discovery of novel Antigens. J. Biomol. Screen. 2011, 16, 1027–1036. [Google Scholar] [PubMed]
- Nijhof, A.M.; Balk, J.A.; Postigo, M.; Jongejan, F. Selection of reference genes for quantitative RT-PCR studies in Rhipicephalus (Boophilus) microplus and Rhipicephalus appendiculatus ticks and determination of the expression profile of Bm86. BMC Mol. Biol. 2009, 10, 112. [Google Scholar] [CrossRef]
- Nijhof, A.M.; Balk, J.A.; Postigo, M.; Rhebergen, A.M.; Taoufik, A.; Jongejan, F. Bm86 homologues and novel ATAQ proteins with multiple epidermal growth factor (EGF)-like domains from hard and soft ticks. Int. J. Parasitol. 2010, 40, 1587–1597. [Google Scholar] [CrossRef]
- Ben Said, M.; Galai, Y.; Mhadhbi, M.; Jedidi, M.; de la Fuente, J.; Darghouth, M.A. Molecular characterization of Bm86 gene orthologs from Hyalomma excavatum, Hyalomma dromedarii and Hyalomma marginatum marginatum and comparison with a vaccine candidate from Hyalomma scupense. Vet. Parasitol. 2012, 190, 230–240. [Google Scholar] [CrossRef]
- Coumou, J.; Wagemakers, A.; Trentelman, J.J.; Nijhof, A.M.; Hovius, J.W. Vaccination against Bm86 homologues in rabbits does not impair Ixodes ricinus feeding or oviposition. PLoS One 2015, 10, e0123495. [Google Scholar]
- Bastos, R.G.; Ueti, M.W.; Knowles, D.P.; Scoles, G.A. The Rhipicephalus (Boophilus) microplus Bm86 gene plays a critical role in the fitness of ticks fed on cattle during acute Babesia bovis infection. Parasites Vectors 2010, 3, 1–11. [Google Scholar] [CrossRef]
- Adjou Moumouni, P.F.; Naomasa, S.; Tuvshintulga, B.; Sato, N.; Okado, K.; Zheng, W.; Lee, S.-H.; Mosqueda, J.; Suzuki, H.; Xuan, X. Identification and Characterization of Rhipicephalus microplus ATAQ Homolog from Haemaphysalis longicornis Ticks and Its Immunogenic Potential as an Anti-Tick Vaccine Candidate Molecule. Microorganisms 2023, 11, 822. [Google Scholar]
- Lugo-Caro del Castillo, S.M.; Hernández-Ortiz, R.; Gómez-Romero, N.; Martínez-Velázquez, M.; Castro-Saines, E.; Lagunes-Quintanilla, R. Genetic diversity of the ATAQ gene in Rhipicephalus microplus collected in Mexico and implications as anti-tick vaccine. Parasitol. Res. 2020, 119, 3523–3529. [Google Scholar] [CrossRef] [PubMed]
- Barry, M.A.; Lai, W.C.; Johnston, S.A. Protection against mycoplasma infection using expression-library immunization. Nature 1995, 377, 632–635. [Google Scholar] [CrossRef] [PubMed]
- DeRisi, J.; Penland, L.; Brown, P.O.; Bittner, M.L.; Meltzer, P.S.; Ray, M.; Chen, Y.; Su, Y.A.; Trent, J.M. Use of a cDNA microarray to analyse gene expression patterns in human cancer. Nat. Genet. 1996, 14, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S.; Johnson, M.; Bridgham, J.; Golda, G.; Lloyd, D.H.; Johnson, D.; Luo, S.; McCurdy, S.; Foy, M.; Ewan, M.; et al. Gene expression analysis by massively parallel signature sequencing (MPSS) on microbead arrays. Nat. Biotechnol. 2000, 18, 630–634. [Google Scholar] [CrossRef]
- Hill, C.A.; Wikel, S.K. The Ixodes scapularis Genome Project: An opportunity for advancing tick research. Trends Parasitol. 2005, 21, 151–153. [Google Scholar] [CrossRef]
- Guerrero, F.D.; Nene, V.M.; George, J.E.; Barker, S.C.; Willadsen, P. Sequencing a new target genome: The Boophilus microplus (Acari: Ixodidae) genome project. J. Med. Entomol. 2006, 43, 9–16. [Google Scholar] [CrossRef]
- Barrero, R.A.; Guerrero, F.D.; Black, M.; McCooke, J.; Chapman, B.; Schilkey, F.; de León, A.A.P.; Miller, R.J.; Bruns, S.; Dobry, J. Gene-enriched draft genome of the cattle tick Rhipicephalus microplus: Assembly by the hybrid Pacific Biosciences/Illumina approach enabled analysis of the highly repetitive genome. Int. J. Parasitol. 2017, 47, 569–583. [Google Scholar]
- Guerrero, F.D.; Ghaffari, N.; Bendele, K.G.; Metz, R.P.; Dickens, C.M.; Blood, P.D.; Tidwell, J.; Miller, R.J.; de León, A.A.P.; Teel, P.D. Raw pacific biosciences and illumina sequencing reads and assembled genome data for the cattle ticks Rhipicephalus microplus and Rhipicephalus annulatus. Data Brief. 2021, 35, 106852. [Google Scholar] [CrossRef]
- De, S.; Kingan, S.B.; Kitsou, C.; Portik, D.M.; Foor, S.D.; Frederick, J.C.; Rana, V.S.; Paulat, N.S.; Ray, D.A.; Wang, Y.; et al. A high-quality Ixodes scapularis genome advances tick science. Nat. Genet. 2023, 55, 301–311. [Google Scholar] [CrossRef]
- Cheung, F.; Haas, B.J.; Goldberg, S.M.; May, G.D.; Xiao, Y.; Town, C.D. Sequencing Medicago truncatula expressed sequenced tags using 454 Life Sciences technology. BMC Genom. 2006, 7, 272. [Google Scholar] [CrossRef]
- Bainbridge, M.N.; Warren, R.L.; Hirst, M.; Romanuik, T.; Zeng, T.; Go, A.; Delaney, A.; Griffith, M.; Hickenbotham, M.; Magrini, V.; et al. Analysis of the prostate cancer cell line LNCaP transcriptome using a sequencing-by-synthesis approach. BMC Genom. 2006, 7, 246. [Google Scholar] [CrossRef]
- Thompson, D.M.; Khalil, S.M.; Jeffers, L.A.; Sonenshine, D.E.; Mitchell, R.D.; Osgood, C.J.; Michael Roe, R. Sequence and the developmental and tissue-specific regulation of the first complete vitellogenin messenger RNA from ticks responsible for heme sequestration. Insect Biochem. Mol. Biol. 2007, 37, 363–374. [Google Scholar] [CrossRef]
- Saldivar, L.; Guerrero, F.; Miller, R.; Bendele, K.; Gondro, C.; Brayton, K. Microarray analysis of acaricide-inducible gene expression in the southern cattle tick, Rhipicephalus (Boophilus) microplus. Insect Mol. Biol. 2008, 17, 597–606. [Google Scholar] [PubMed]
- Valle, M.R.; Guerrero, F.D. Anti-tick vaccines in the omics era. Front. Biosci. 2018, 10, 122–136. [Google Scholar] [CrossRef]
- Aljamali, M.N.; Sauer, J.R.; Essenberg, R.C. RNA interference: Applicability in tick research. Exp. Appl. Acarol. 2002, 28, 89–96. [Google Scholar] [CrossRef]
- Almazan, C.; Kocan, K.M.; Bergman, D.K.; Garcia-Garcia, J.C.; Blouin, E.F.; de la Fuente, J. Identification of protective antigens for the control of Ixodes scapularis infestations using cDNA expression library immunization. Vaccine 2003, 21, 1492–1501. [Google Scholar] [CrossRef]
- de la Fuente, J.; Almazán, C.; Blouin, E.F.; Naranjo, V.; Kocan, K.M. RNA interference screening in ticks for identification of protective antigens. Parasitol. Res. 2005, 96, 137–141. [Google Scholar]
- Nijhof, A.M.; Taoufik, A.; de la Fuente, J.; Kocan, K.M.; de Vries, E.; Jongejan, F. Gene silencing of the tick protective antigens, Bm86, Bm91 and subolesin, in the one-host tick Boophilus microplus by RNA interference. Int. J. Parasitol. 2007, 37, 653–662. [Google Scholar] [CrossRef]
- De la Fuente, J.; Kocan, K. Strategies for development of vaccines for control of ixodid tick species. Parasite Immunol. 2006, 28, 275–283. [Google Scholar]
- Peconick, A.; Sossai, S.; Girao, F.; Rodrigues, M.; e Silva, C.S.; Vargas, M.; Patarroyo, J. Synthetic vaccine (SBm7462) against the cattle tick Rhipicephalus (Boophilus) microplus: Preservation of immunogenic determinants in different strains from South America. Exp. Parasitol. 2008, 119, 37–43. [Google Scholar] [CrossRef]
- Leal, B.F.; Ferreira, C.A.S. Ticks and antibodies: May parasite density and tick evasion influence the outcomes following immunization protocols? Vet. Parasitol. 2021, 300, 109610. [Google Scholar]
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef]
- Fisch, A.; Reynisson, B.; Benedictus, L.; Nicastri, A.; Vasoya, D.; Morrison, I.; Buus, S.; Ferreira, B.R.; Kinney Ferreira de Miranda Santos, I.; Ternette, N. Integral Use of Immunopeptidomics and Immunoinformatics for the Characterization of Antigen Presentation and Rational Identification of BoLA-DR–Presented Peptides and Epitopes. J. Immunol. 2021, 206, 2489–2497. [Google Scholar] [PubMed]
- Vasoya, D.; Oliveira, P.S.; Muriel, L.A.; Tzelos, T.; Vrettou, C.; Morrison, W.I.; de Miranda Santos, I.K.F.; Connelley, T. High throughput analysis of MHC-I and MHC-DR diversity of Brazilian cattle populations. HLA 2021, 98, 93–113. [Google Scholar] [CrossRef] [PubMed]
- Ndawula, C., Jr.; Tabor, A.E. Cocktail Anti-Tick Vaccines: The Unforeseen Constraints and Approaches toward Enhanced Efficacies. Vaccines 2020, 8, 457. [Google Scholar] [CrossRef] [PubMed]
- Suarez, M.; Rubi, J.; Pérez, D.; Cordova, V.; Salazar, Y.; Vielma, A.; Barrios, F.; Gil, C.A.; Segura, N.; Carrillo, Y. High impact and effectiveness of Gavac™ vaccine in the national program for control of bovine ticks Rhipicephalus microplus in Venezuela. Livest. Sci. 2016, 187, 48–52. [Google Scholar]
- Schetters, T.; Bishop, R.; Crampton, M.; Kopacek, P.; Lew-Tabor, A.; Maritz-Olivier, C.; Miller, R.; Mosqueda, J.; Patarroyo, J.; Rodriguez-Valle, M.; et al. Cattle tick vaccine researchers join forces in CATVAC. Parasit. Vectors 2016, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- Van Zee, J.P.; Geraci, N.; Guerrero, F.; Wikel, S.; Stuart, J.; Nene, V.; Hill, C. Tick genomics: The Ixodes genome project and beyond. Int. J. Parasitol. 2007, 37, 1297–1305. [Google Scholar] [CrossRef]
- Parizi, L.F.; Reck Jr, J.; Oldiges, D.P.; Guizzo, M.G.; Seixas, A.; Logullo, C.; de Oliveira, P.L.; Termignoni, C.; Martins, J.R.; da Silva Vaz Jr, I. Multi-antigenic vaccine against the cattle tick Rhipicephalus (Boophilus) microplus: A field evaluation. Vaccine 2012, 30, 6912–6917. [Google Scholar]
- Scheiblhofer, S.; Laimer, J.; Machado, Y.; Weiss, R.; Thalhamer, J. Influence of protein fold stability on immunogenicity and its implications for vaccine design. Exp. Rev. Vaccines 2017, 16, 479–489. [Google Scholar]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [PubMed]
- Kumru, O.S.; Joshi, S.B.; Smith, D.E.; Middaugh, C.R.; Prusik, T.; Volkin, D.B. Vaccine instability in the cold chain: Mechanisms, analysis and formulation strategies. Biologicals 2014, 42, 237–259. [Google Scholar] [PubMed]
- Taylor, K.; Gillam, F.; Lohneis, T.; Zhang, C. Designs of antigen structure and composition for improved protein-based vaccine efficacy. Front. Immunol. 2020, 11, 283. [Google Scholar]
- Vargas-Hernández, M.; Santana-Rodríguez, E.; Montero-Espinosa, C.; Sordo-Puga, Y.; Acosta-Hernández, A.; Fuentes-Rodríguez, Y.; Pérez-Pérez, D.; Oliva-Cárdenas, A.; González-Ramos, E.; Duarte, C.A. Stability, safety and protective immunity of Gavac® vaccine subjected to heat stress. Biotecnol. Apl. 2018, 35, 1501–1503. [Google Scholar]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Anasir, M.I.; Poh, C.L. Structural vaccinology for viral vaccine design. Front. Microbiol. 2019, 10, 738. [Google Scholar]
- Dormitzer, P.R.; Grandi, G.; Rappuoli, R. Structural vaccinology starts to deliver. Nat. Rev. Microbiol. 2012, 10, 807–813. [Google Scholar]
- Cozzi, R.; Scarselli, M.; Ilaria Ferlenghi, I.F. Structural vaccinology: A three-dimensional view for vaccine development. Curr. Top. Med. Chem. 2013, 13, 2629–2637. [Google Scholar]
- Kumar, R.; Kumar, P. Yeast-based vaccines: New perspective in vaccine development and application. FEMS Yeast Res. 2019, 19, foz007. [Google Scholar] [CrossRef]
- Lestari, C.; Novientri, G. Advantages of yeast-based recombinant protein technology as vaccine products against infectious diseases. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 913, p. 012099. [Google Scholar]
- Carter, D.; Duthie, M.S.; Reed, S.G. Adjuvants. In Vaccination Strategies against Highly Variable Pathogens; NIH: Bethesda, MD, USA, 2020; pp. 103–127. [Google Scholar]
- Reed, S.G.; Tomai, M.; Gale Jr, M.J. New horizons in adjuvants for vaccine development. Curr. Opin. Immunol. 2020, 65, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Aucouturier, J.; Dupuis, L.; Deville, S.; Ascarateil, S.; Ganne, V. Montanide ISA 720 and 51: A new generation of water in oil emulsions as adjuvants for human vaccines. Exp. Rev. Vaccines 2002, 1, 111–118. [Google Scholar]
- Aucouturier, J.; Dupuis, L.; Ganne, V. Adjuvants designed for veterinary and human vaccines. Vaccine 2001, 19, 2666–2672. [Google Scholar] [CrossRef] [PubMed]
- Heegaard, P.M.; Dedieu, L.; Johnson, N.; Le Potier, M.F.; Mockey, M.; Mutinelli, F.; Vahlenkamp, T.; Vascellari, M.; Sorensen, N.S. Adjuvants and delivery systems in veterinary vaccinology: Current state and future developments. Arch. Virol. 2011, 156, 183–202. [Google Scholar] [CrossRef]
- Heegaard, P.M.; Fang, Y.; Jungersen, G. Novel adjuvants and immunomodulators for veterinary vaccines. Vaccine Technol. Vet. Viral Dis. Methods Protoc. 2016, 1349, 63–82. [Google Scholar]
- Aguilar, J.C.; Rodriguez, E.G. Vaccine adjuvants revisited. Vaccine 2007, 25, 3752–3762. [Google Scholar] [CrossRef]
- Reed, S.G.; Bertholet, S.; Coler, R.N.; Friede, M. New horizons in adjuvants for vaccine development. Trends Immunol. 2009, 30, 23–32. [Google Scholar] [CrossRef]
- Chauhan, N.; Tiwari, S.; Iype, T.; Jain, U. An overview of adjuvants utilized in prophylactic vaccine formulation as immunomodulators. Exp. Rev. Vaccines 2017, 16, 491–502. [Google Scholar]
- Spickler, A.R.; Roth, J.A. Adjuvants in veterinary vaccines: Modes of action and adverse effects. J. Vet. Intern. Med. 2003, 17, 273–281. [Google Scholar]
- Van Donkersgoed, J.; Dubeski, P.L.; Aalhus, J.L.; VanderKop, M.; Dixon, S.; Starr, W.N. The effect of vaccines and antimicrobials on the formation of injection site lesions in subprimals of experimentally injected beef calves. Can. Vet. J. 1999, 40, 245. [Google Scholar]
- Meng, X.-J. Porcine Circovirus Type 2 (PCV2): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2013, 1, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Hou, L.; Liu, J. Host immune response to infection with porcine circoviruses. Anim. Dis. 2021, 1, 1–10. [Google Scholar]
- Rakibuzzaman, A.; Ramamoorthy, S. Comparative immunopathogenesis and biology of recently discovered porcine circoviruses. Transbound. Emerg. Dis. 2021, 68, 2957–2968. [Google Scholar] [CrossRef] [PubMed]
- Gershwin, L.J. Immunology of bovine respiratory syncytial virus infection of cattle. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.C. Human and bovine respiratory syncytial virus: Immunopathologic mechanisms. Vet. Q. 1991, 13, 47–59. [Google Scholar] [CrossRef]
- Mc Keever, D. Cellular immunity against Theileria parva and its influence on parasite diversity. Res. Vet. Sci. 2001, 70, 77–81. [Google Scholar] [CrossRef]
- Dobbelaere, D.A.; Fernandez, P.C.; Heussler, V.T. Theileria parva: Taking control of host cell proliferation and survival mechanisms: Microreview. Cell. Microbiol. 2000, 2, 91–99. [Google Scholar] [CrossRef]
- McKeever, D.J.; Taracha, E.; Morrison, W.I.; Musoke, A.; Morzaria, S. Protective immune mechanisms against Theileria parva: Evolution of vaccine development strategies. Parasitol. Today 1999, 15, 263–267. [Google Scholar] [CrossRef]
- Mattioli, R.C.; Pandey, V.S.; Murray, M.; Fitzpatrick, J.L. Immunogenetic influences on tick resistance in African cattle with particular reference to trypanotolerant N’Dama (Bos taurus) and trypanosusceptible Gobra zebu (Bos indicus) cattle. Acta Trop. 2000, 75, 263–277. [Google Scholar] [CrossRef]
- Nascimento, C.; Machado, M.; Guimarães, S.; Martins, M.; Peixoto, J.; Furlong, J.; Prata, M.; Verneque, R.; Teodoro, R.; Lopes, P. Expressed sequenced tags profiling of resistant and susceptible Gyr x Holstein cattle infested with the tick Rhipicephalus (Boophilus) microplus. Genet. Mol. Res. 2011, 10, 3803–3816. [Google Scholar] [CrossRef]
- Robbertse, L.; Richards, S.A.; Clift, S.J.; Barnard, A.C.; Leisewitz, A.; Crafford, J.E.; Maritz-Olivier, C. Comparison of the differential regulation of T and B-lymphocyte subsets in the skin and lymph nodes amongst three cattle breeds as potential mediators of immune-resistance to Rhipicephalus microplus. Ticks Tick. Borne Dis. 2018, 9, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Piper, E.K.; Jonsson, N.N.; Gondro, C.; Lew-Tabor, A.E.; Moolhuijzen, P.; Vance, M.E.; Jackson, L.A. Immunological profiles of Bos taurus and Bos indicus cattle infested with the cattle tick, Rhipicephalus (Boophilus) microplus. Clin. Vaccine Immunol. 2009, 16, 1074–1086. [Google Scholar] [CrossRef] [PubMed]
- Kazimírová, M.; Štibrániová, I. Tick salivary compounds: Their role in modulation of host defences and pathogen transmission. Front. Cell. Infect. Microbiol. 2013, 3, 43. [Google Scholar] [CrossRef]
- Untalan, P.M.; Pruett, J.H.; Steelman, C.D. Association of the bovine leukocyte antigen major histocompatibility complex class II DRB3* 4401 allele with host resistance to the Lone Star tick, Amblyomma americanum. Vet. Parasitol. 2007, 145, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Holmes, E.C.; Roberts, A.F.; Staines, K.A.; Ellis, S.A. Evolution of major histocompatibility complex class I genes in Cetartiodactyls. Immunogenetics 2003, 55, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.A.; Marsh, S.G.; Robinson, J.; Davies, C.J.; Stear, M.J.; Ellis, S.A. Cattle MHC nomenclature: Is it possible to assign sequences to discrete class I genes? Immunogenetics 2012, 64, 475–480. [Google Scholar] [CrossRef]
- Gelhaus, A.; Förster, B.; Horstmann, R.D. Evidence for an additional cattle DQB locus. Immunogenetics 1999, 49, 879–885. [Google Scholar] [CrossRef]
- Gelhaus, A.; Förster, B.; Wippern, C.; Horstmann, R.D. Evidence for an additional cattle DQA locus, BoLA-DQA5. Immunogenetics 1999, 49, 321–327. [Google Scholar] [CrossRef]
- Andersson, L.; Rask, L. Characterization of the MHC class II region in cattle. The number of DQ genes varies between haplotypes. Immunogenetics 1988, 27, 110–120. [Google Scholar] [CrossRef]
- Zhou, H.; Hickford, J.; Fang, Q.; Byun, S. Identification of allelic variation at the bovine DRA locus by polymerase chain reaction-single strand conformational polymorphism. J. Dairy. Sci. 2007, 90, 1943–1946. [Google Scholar] [CrossRef]
- Burke, M.; Stone, R.; Muggli-Cockett, N. Nucleotide sequence and Northern analysis of a bovine major histocompatibility class II DRß-like cDNA. Anim. Genet. 1991, 22, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Takeshima, S.-N.; Corbi-Botto, C.; Giovambattista, G.; Aida, Y. Genetic diversity of BoLA-DRB3 in South American Zebu cattle populations. BMC Genet. 2018, 19, 33. [Google Scholar] [CrossRef] [PubMed]
- Baltian, L.R.; Ripoli, M.V.; Sanfilippo, S.; Takeshima, S.; Aida, Y.; Giovambattista, G. Association between BoLA-DRB3 and somatic cell count in Holstein cattle from Argentina. Mol. Biol. Rep. 2012, 39, 7215–7220. [Google Scholar] [CrossRef] [PubMed]
- Miyasaka, T.; Takeshima, S.n.; Jimba, M.; Matsumoto, Y.; Kobayashi, N.; Matsuhashi, T.; Sentsui, H.; Aida, Y. Identification of bovine leukocyte antigen class II haplotypes associated with variations in bovine leukemia virus proviral load in Japanese Black cattle. Tissue Antigens 2013, 81, 72–82. [Google Scholar] [CrossRef]
- Tiele, D.; Sebro, E.; H/Meskel, D.; Mathewos, M. Epidemiology of Gastrointestinal Parasites of Cattle in and Around Hosanna Town, Southern Ethiopia. Vet. Med. Res. Rep. 2023, 14, 1–9. [Google Scholar] [CrossRef]
- Marskole, P.; Verma, Y.; Dixit, A.K.; Swamy, M. Prevalence and burden of gastrointestinal parasites in cattle and buffaloes in Jabalpur, India. Vet. World 2016, 9, 1214. [Google Scholar] [CrossRef]
- Thapa, B.; Parajuli, R.P.; Dhakal, P. Prevalence and burden of gastrointestinal parasites in stray cattle of the Kathmandu Valley. J. Parasit. Dis. 2022, 46, 845–853. [Google Scholar] [CrossRef]
- Urban, J.F., Jr.; Steenhard, N.R.; Solano-Aguilar, G.I.; Dawson, H.D.; Iweala, O.I.; Nagler, C.R.; Noland, G.S.; Kumar, N.; Anthony, R.M.; Shea-Donohue, T. Infection with parasitic nematodes confounds vaccination efficacy. Vet. Parasitol. 2007, 148, 14–20. [Google Scholar] [CrossRef]
- Hananeh, W.M.; Radhi, A.; Mukbel, R.M.; Ismail, Z.B. Effects of parasites coinfection with other pathogens on animal host: A literature review. World 2022, 15, 2414–2424. [Google Scholar] [CrossRef]
- Mabbott, N.A. The influence of parasite infections on host immunity to co-infection with other pathogens. Front. Immunol. 2018, 9, 2579. [Google Scholar] [CrossRef]
- Zhu, F.; Liu, W.; Liu, T.; Shi, L.; Zheng, W.; Guan, F.; Lei, J. A New Role for Old Friends: Effects of Helminth Infections on Vaccine Efficacy. Pathogens 2022, 11, 1163. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Su, X.-Z.; Lu, F. The Roles of Type I Interferon in Co-infections With Parasites and Viruses, Bacteria, or Other Parasites. Front. Immunol. 2020, 11, 1805. [Google Scholar] [CrossRef] [PubMed]
- Torina, A.; Blanda, V.; Villari, S.; Piazza, A.; La Russa, F.; Grippi, F.; La Manna, M.P.; Di Liberto, D.; de la Fuente, J.; Sireci, G. Immune response to Tick-Borne hemoparasites: Host adaptive immune response mechanisms as potential targets for therapies and vaccines. Int. J. Mol. Sci. 2020, 21, 8813. [Google Scholar] [CrossRef] [PubMed]
- Thumbi, S.M.; Bronsvoort, B.M.; Poole, E.J.; Kiara, H.; Toye, P.G.; Mbole-Kariuki, M.N.; Conradie, I.; Jennings, A.; Handel, I.G.; Coetzer, J.A.; et al. Parasite co-infections and their impact on survival of indigenous cattle. PLoS ONE 2014, 9, e76324. [Google Scholar] [CrossRef]
- Vaumourin, E.; Vourc’h, G.; Gasqui, P.; Vayssier-Taussat, M. The importance of multiparasitism: Examining the consequences of co-infections for human and animal health. Parasit. Vectors 2015, 8, 545. [Google Scholar] [CrossRef]
- Ahmed, N.; French, T.; Rausch, S.; Kuhl, A.; Hemminger, K.; Dunay, I.R.; Steinfelder, S.; Hartmann, S. Toxoplasma Co-infection Prevents Th2 Differentiation and Leads to a Helminth-Specific Th1 Response. Front. Cell Infect. Microbiol. 2017, 7, 341. [Google Scholar] [CrossRef]
- Newton, L.G. Acaricide resistance and cattle tick control. Aust. Vet. J. 1967, 43, 389–394. [Google Scholar] [CrossRef]
- Cobbett, N.G. Preliminary tests in Mexico with DDT, cube, hexachlorocyclohexane (benzene hexachloride) and combinations thereof, for the control of the cattle fever tick, Boophilus annulatus. Am. J. Vet. Res. 1947, 8, 280–283. [Google Scholar]
- Cook, J. Action of rumen fluid on pesticides, in vitro destruction of some organophosphate pesticides by bovine rumen fluid. J. Agricult. Food Chem. 1957, 5, 859–863. [Google Scholar] [CrossRef]
- O’Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 1975, 250, 4007–4021. [Google Scholar] [CrossRef]
- Grunstein, M.; Hogness, D.S. Colony hybridization: A method for the isolation of cloned DNAs that contain a specific gene. Proc. Natl. Acad. Sci. USA 1975, 72, 3961–3965. [Google Scholar] [CrossRef]
- Hollingworth, R. Chemistry, biological activity, and uses of formamidine pesticides. Environ. Health Perspect. 1976, 14, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed]
- Itakura, K.; Hirose, T.; Crea, R.; Riggs, A.D.; Heyneker, H.L.; Bolivar, F.; Boyer, H.W. Expression in Escherichia coli of a chemically synthesized gene for the hormone somatostatin. Science 1977, 198, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Nolan, J.; Roulston, W.; Schnitzerling, H. The potential of some synthetic pyrethroids for control of the cattle tick (Boophilus microplus). Austr. Vet. J. 1979, 55, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.A. The eradication of smallpox. Sci. Am. 1976, 235, 25–33. [Google Scholar] [CrossRef]
- Taylor, M.G. Irradiation-attenuated anti-parasite vaccines in ruminants. Present status and future prospects. In Isotopes and Radiation in Parasitology IV; IAEA: Vienna, Austria, 1981. [Google Scholar]
- Putter, I.; Connell, J.M.; Preiser, F.; Haidri, A.; Ristich, S.; Dybas, R. Avermectins: Novel insecticides, acaricides and nematicides from a soil microorganism. Experientia 1981, 37, 963–964. [Google Scholar] [CrossRef]
- Johnson, I.S. Human insulin from recombinant DNA technology. Science 1983, 219, 632–637. [Google Scholar] [CrossRef]
- Mullis, K.B. Target amplification for DNA analysis by the polymerase chain reaction. Ann. Biol. Clin. 1990, 48, 579–582. [Google Scholar]
- Davey, R.B.; Ahrens, E.H.; George, J.E.; Hunter, J.S., III; Jeannin, P. Therapeutic and persistent efficacy of fipronil against Boophilus microplus (Acari: Ixodidae) on cattle. Vet. Parasitol. 1998, 74, 261–276. [Google Scholar] [CrossRef]
- Moffat, A.S. New chemicals seek to outwit insect pests. Science 1993, 261, 550–551. [Google Scholar] [CrossRef]
- Luguru, S.; Chizyuka, H.; Musisi, F. A survey for resistance to acaricides in cattle ticks (Acari: Ixodidae) in three major traditional cattle areas in Zambia. Bull. Entomol. Res. 1987, 77, 569–574. [Google Scholar] [CrossRef]
- Bull, M.; Swindale, S.; Overend, D.; Hess, E. Suppression of Boophilus microplus populations with fluazuron–an acarine growth regulator. Austr. Vet. J. 1996, 74, 468–470. [Google Scholar] [CrossRef]
- Martins Joao, R.; Corrêa, B.; Ceresér, V.; Arteche, C.C. A Situation Report on Resistance to Acaricides by the Cattle Tick Boophilus microplus in the State of Rio Grande do Sul, Southern Brazil; Publicacion Especial; NHI: Bethesda, MD, USA, 1995. [Google Scholar]
- Martins, J.; Furlong, J. Avermectin resistance of the cattle tick Boophilus microplus in Brazil. Vet. Record 2001, 149, 64. [Google Scholar]
- Mikheyev, A.S.; Tin, M.M. A first look at the Oxford Nanopore MinION sequencer. Mol. Ecol. Resour. 2014, 14, 1097–1102. [Google Scholar] [CrossRef]
- Cuore, U.; Trelles, A.; Sanchís, J.; Gayo, V.; Solari, M. First diagnosis of resistance to fipronil in the common cattle tick Boophilus microplus. Veterinaria 2007, 42, 35–41. [Google Scholar]
- Nagalakshmi, U.; Wang, Z.; Waern, K.; Shou, C.; Raha, D.; Gerstein, M.; Snyder, M. The transcriptional landscape of the yeast genome defined by RNA sequencing. Science 2008, 320, 1344–1349. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Schwarz, A.; von Reumont, B.M.; Erhart, J.; Chagas, A.C.; Ribeiro, J.M.; Kotsyfakis, M. De novo Ixodes ricinus salivary gland transcriptome analysis using two next-generation sequencing methodologies. FASEB J. 2013, 27, 4745–4756. [Google Scholar] [CrossRef]
- Heekin, A.M.; Guerrero, F.D.; Bendele, K.G.; Saldivar, L.; Scoles, G.A.; Dowd, S.E.; Gondro, C.; Nene, V.; Djikeng, A.; Brayton, K.A. The ovarian transcriptome of the cattle tick, Rhipicephalus (Boophilus) microplus, feeding upon a bovine host infected with Babesia bovis. Parasit. Vectors 2013, 6, 276. [Google Scholar] [CrossRef] [PubMed]
- Heekin, A.M.; Guerrero, F.D.; Bendele, K.G.; Saldivar, L.; Scoles, G.A.; Dowd, S.E.; Gondro, C.; Nene, V.; Djikeng, A.; Brayton, K.A. Gut transcriptome of replete adult female cattle ticks, Rhipicephalus (Boophilus) microplus, feeding upon a Babesia bovis-infected bovine host. Parasitol. Res. 2013, 112, 3075–3090. [Google Scholar] [CrossRef]
- Reck, J.; Klafke, G.M.; Webster, A.; Dall’Agnol, B.; Scheffer, R.; Souza, U.A.; Corassini, V.B.; Vargas, R.; dos Santos, J.S.; de Souza Martins, J.R. First report of fluazuron resistance in Rhipicephalus microplus: A field tick population resistant to six classes of acaricides. Vet. Parasitol. 2014, 201, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Gulia-Nuss, M.; Nuss, A.B.; Meyer, J.M.; Sonenshine, D.E.; Roe, R.M.; Waterhouse, R.M.; Sattelle, D.B.; de la Fuente, J.; Ribeiro, J.M.; Megy, K.; et al. Genomic insights into the Ixodes scapularis tick vector of Lyme disease. Nat. Commun. 2016, 7, 10507. [Google Scholar] [CrossRef] [PubMed]
- Datoo, M.S.; Natama, M.H.; Some, A.; Traore, O.; Rouamba, T.; Bellamy, D.; Yameogo, P.; Valia, D.; Tegneri, M.; Ouedraogo, F.; et al. Efficacy of a low-dose candidate malaria vaccine, R21 in adjuvant Matrix-M, with seasonal administration to children in Burkina Faso: A randomised controlled trial. Lancet 2021, 397, 1809–1818. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bishop, L.J.; Stutzer, C.; Maritz-Olivier, C. More than Three Decades of Bm86: What We Know and Where to Go. Pathogens 2023, 12, 1071. https://doi.org/10.3390/pathogens12091071
Bishop LJ, Stutzer C, Maritz-Olivier C. More than Three Decades of Bm86: What We Know and Where to Go. Pathogens. 2023; 12(9):1071. https://doi.org/10.3390/pathogens12091071
Chicago/Turabian StyleBishop, Laura Jane, Christian Stutzer, and Christine Maritz-Olivier. 2023. "More than Three Decades of Bm86: What We Know and Where to Go" Pathogens 12, no. 9: 1071. https://doi.org/10.3390/pathogens12091071