
50 Years since Kaufman and Phillips’ Groundbreaking Trilogy Elucidating Ion and Water Homeostasis in Ixodid Ticks

{kind=link}
{kind=link}
Abstract
:1. Introduction
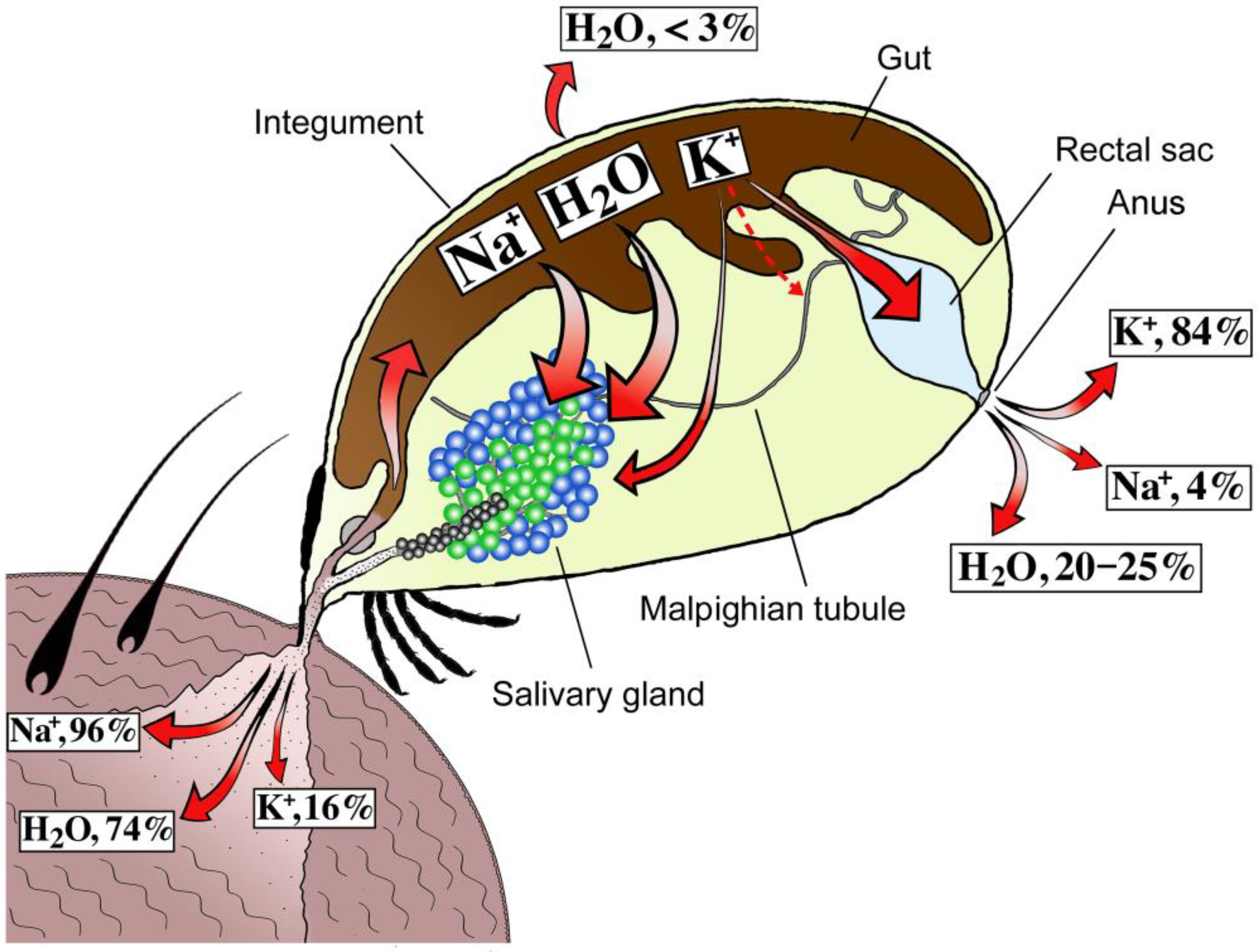
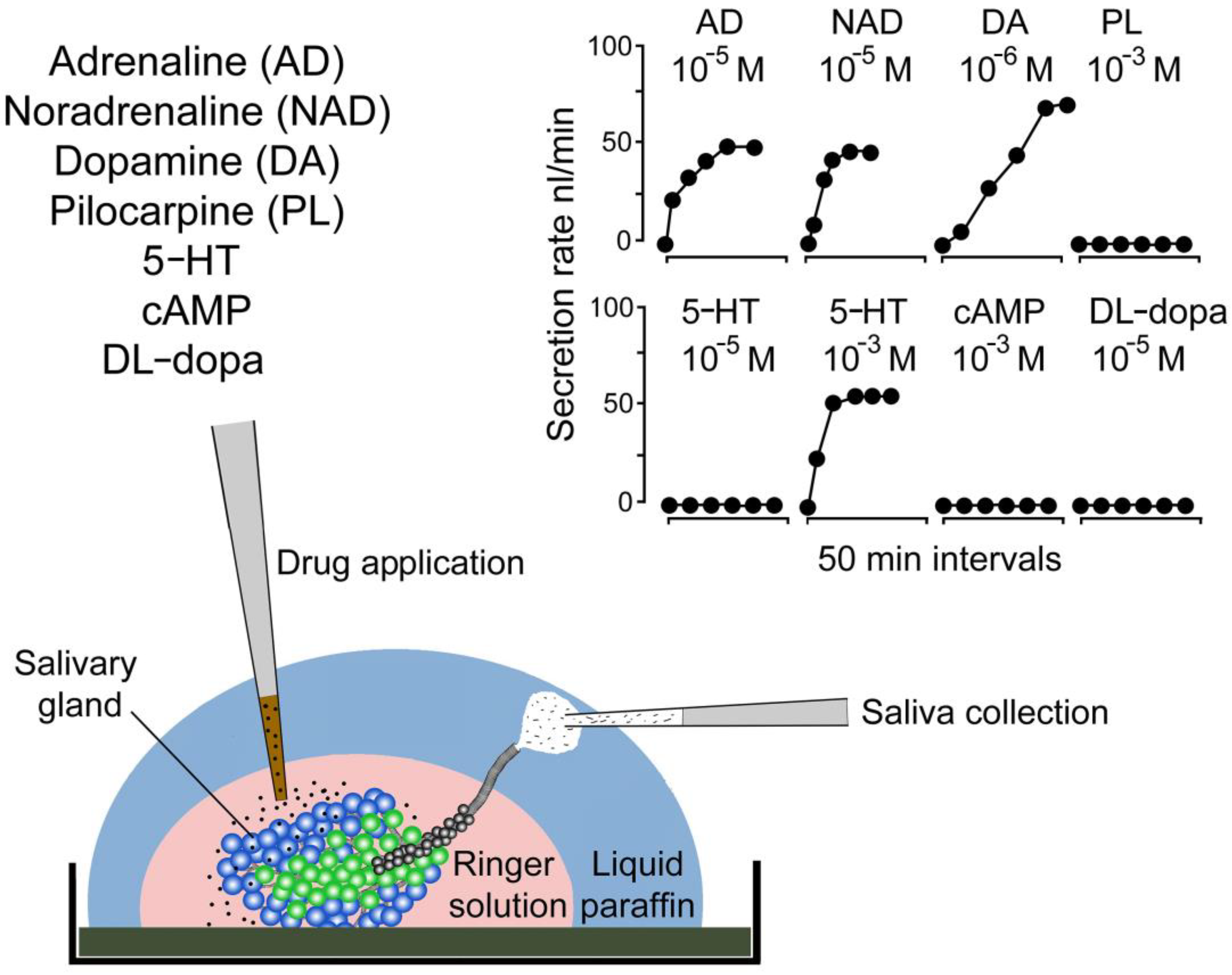
2. Discoveries
3. Impact and Follow-Up Research
Funding
Acknowledgments
Conflicts of Interest
References
- Ramsay, J.A. Active Transport of Potassium by the Malpighian Tubules of Insects. J. Exp. Bio. 1953, 30, 358–369. [Google Scholar] [CrossRef]
- Ramsay, J.A. The Excretion of Sodium, Potassium and Water by the Malpighian Tubules of the Stick Insect, Dixippus morosus (Orthoptera, Phasmidae). J. Exp. Bio. 1955, 32, 200–216. [Google Scholar] [CrossRef]
- Wigglesworth, V. The Physiology of Excretion in a Blood-Sucking Insect, Rhodnius prolixus (Hemiptera, Reduviidae). J. Exp. Bio. 1931, 8, 411–427. [Google Scholar] [CrossRef]
- Benoit, J.B.; Denlinger, D.L. Meeting the Challenges of On-Host and off-Host Water Balance in Blood-Feeding Arthropods. J. Insect Physiol. 2010, 56, 1366–1376. [Google Scholar] [CrossRef] [Green Version]
- Orchard, I.; Lange, A.B. Hormonal Control of Diuresis in Insects. In Advances in Invertebrate (Neuro) Endocrinology, 1st ed.; Saleuddin, S., Ed.; Apple Academic Press: Florida, FL, USA, 2020; Volume 2, pp. 225–282. [Google Scholar]
- Park, Y. Endocrine Regulation of Insect Diuresis in the Early Postgenomic Era1. Can. J. Zool. 2012, 90, 507–520. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, S.E.; Kaufman, W.R.; Phillips, J.E. Mechanism and Characteristics of Coxal Fluid Excretion in the Argasid Tick Ornithodoros moubata. J. Exp. Bio. 1982, 98, 343–352. [Google Scholar] [CrossRef]
- Binnington, K.C. Secretory Coxal Gland, Active during Apolysis in Ixodid and Argasid Ticks (Acarina). Int. J. Insect Morphol. Embryol. 1975, 4, 183–191. [Google Scholar] [CrossRef]
- Kaufman, W.R. Ticks: Physiological Aspects with Implications for Pathogen Transmission. Ticks Tick Borne Dis. 2010, 1, 11–22. [Google Scholar] [CrossRef]
- Sonenshine, D.E. Biology of Ticks, 1st ed.; Oxford University Press: New York, NY, USA, 1991. [Google Scholar]
- Kaufman, W. Tick-Host Interaction: A Synthesis of Current Concepts. Parasitol. Today 1989, 5, 47–56. [Google Scholar] [CrossRef]
- Kaufman, W.R.; Flynn, P.C.; Reynolds, S.E. Cuticular Plasticization in the Tick, Amblyomma hebraeum (Acari: Ixodidae): Possible Roles of Monoamines and Cuticular PH. J. Exp. Bio. 2010, 213, 2820–2831. [Google Scholar] [CrossRef] [Green Version]
- Okura, N.; Kitaura, H.; Möri, T.; Shiraishi, S. Cuticular Plasticization Induced by Copulatory Stimuli in Female Haemaphysalis longicornis (Acari: Ixodidae). J. Med. Entomol. 1996, 33, 702–705. [Google Scholar] [CrossRef] [PubMed]
- Lees, A.D. The Water Balance in Ixodes Ricinus l. and Certain Other Species of Ticks. Parasitology 1946, 37, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Gregson, J.D. Observations on the Movement of Fluids in the Vicinity of the Mouthparts of Naturally Feeding Dermacentor andersoni Stiles. Parasitology 1967, 57, 1–8. [Google Scholar] [CrossRef]
- Tatchell, R.J. Salivary Secretion in the Cattle Tick as a Means of Water Elimination. Nature 1967, 213, 940–941. [Google Scholar] [CrossRef]
- Belozerov, V.N. Water Content and Regulation of Water Balance in Tick, Females (Acarina, Ixodidae) during Feeding and after Engorgement. Zool. Zhurnal 1967, 46, 1182–1187. [Google Scholar]
- Kaufman, W.R.; Phillips, J.E. Ion and Water Balance in the Ixodid Tick Dermacentor andersoni. I. Routes of Ion and Water Excretion. J. Exp. Bio. 1973. [Google Scholar] [CrossRef]
- Kaufman, W.R.; Phillips, J.E. Ion and Water Balance in the Ixodid Tick Dermacentor andersoni: II. Mechanism and Control of Salivary Secretion. J. Exp. Bio. 1973, 58, 537–547. [Google Scholar] [CrossRef]
- Kaufman, W.R.; Phillips, J.E. Ion and Water Balance in the Ixodid Tick Dermacentor andersoni: III. Influence of Monovalent Ions and Osmotic Pressure on Salivary Secretion. J. Exp. Bio. 1973, 58, 549–564. [Google Scholar] [CrossRef]
- Kaufman, S.E. Ion and Water Regulation during Feeding in the Female Tick Ornithodorus moubata. Ph.D. Thesis, University of British Columbia, Vancover, BC, Canada, 1971. [Google Scholar]
- Balashov, Y.S. The Excretion Processes and Function of Malpighian Tubules of Ixodid Ticks. Parasit. Sborn. Zool. Inst. Akad. Nauk. USSR 1958, 18, 120–128. [Google Scholar]
- Purnell, R.E.; Branagan, D.; Radley, D.E. The Use of Parasympathomimetic Drugs to Stimulate Salivation in the Tick Rhipicephalus appendiculatus, and the Transmission of Theileria parva Using Saliva Obtained by This Method from Infected Ticks. Parasitology 1969, 59, 709–718. [Google Scholar] [CrossRef]
- Needham, G.R.; Sauer, J.R. Control of Fluid Secretion by Isolated Salivary Glands of the Lone Star Tick. J. Insect Physiol. 1975, 21, 1893–1898. [Google Scholar] [CrossRef]
- Kaufman, W.R.; Sauer, J.R. Ion and Water Balance in Feeding Ticks: Mechanisms of Tick Excretion. In Physiology of Ticks; Obenchain, F.D., Galun, R., Eds.; Pergamon: Oxford, UK, 1982; pp. 213–244. ISBN 978-0-08-024937-7. [Google Scholar]
- Bowman, A.S.; Sauer, J.R. Tick Salivary Glands: Function, Physiology and Future. Parasitology 2004, 129, S67–S81. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.-H.; Sauer, J.R. Ion and Water Balance in the Feeding Lone Star Tick. Comp. Biochem. Physiol A Physiol. 1975, 52, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Sauer, J.R.; Essenberg, R.C.; Bowman, A.C. Salivary Glands in Ixodid Ticks: Control and Mechanism of Secretion. J. Insect Physiol. 2000, 46, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Sauer, J.R.; McSwain, J.L.; Bowman, A.S.; Essenberg, R.C. Tick Salivary Gland Physiology. Annu. Rev. Entomol. 1995, 40, 245–267. [Google Scholar] [CrossRef]
- Sauer, J.R.; McSwain, J.L.; Essenberg, R.C. Cell Membrane Receptors and Regulation of Cell Function in Ticks and Blood-Sucking Insects. Int. J. Parasitol. 1994, 24, 33–52. [Google Scholar] [CrossRef]
- Kaufman, W. The Influence of Various Factors on Fluid Secretion by in Vitro Salivary Glands of Ixodid Ticks. J. Exp. Bio. 1976, 64, 727–742. [Google Scholar] [CrossRef]
- Kaufman, W.R. The Influence of Adrenergic Agonists and Their Antagonists on Isolated Salivary Glands of Ixodid Ticks. Eur. J. Pharmacol. 1977, 45, 61–68. [Google Scholar] [CrossRef]
- Kaufman, W.R. Actions of Some Transmitters and Their Antagonists on Salivary Secretion in a Tick. Am. J. Physiol. 1978, 235, R76–R81. [Google Scholar] [CrossRef]
- Kaufman, W.R.; Harris, R.A. Neural Pathways Mediating Salivary Fluid Secretion in the Ixodid Tick Amblyomma hebraeum. Can. J. Zool. 1983, 61, 1976–1980. [Google Scholar] [CrossRef]
- Kaufman, W.R.; Wong, D.L. Evidence for Multiple Receptors Mediating Fluid Secretion in Salivary Glands of Ticks. Eur. J. Pharmacol. 1983, 87, 43–52. [Google Scholar] [CrossRef]
- Kaufman, W.R.; Minion, J.L. Pharmacological Characterization of the Ergot Alkaloid Receptor in the Salivary Gland of the Ixodid Tick Amblyomma hebraeum. J. Exp. Bio. 2006, 209, 2525–2534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meredith, J.; Kaufman, W.R. A Proposed Site of Fluid Secretion in the Salivary Gland of the Ixodid Tick, Dermacentor andersoni. Parasitology 1973, 67, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, D.W.; Doxsey, S.J.; Buscher, G. Salivary Gland of the Tick Vector (R. appediculatus) of East Coast Fever. II. Cellular Basis for Fluid Secretion in the Type III Acinus. Tissue Cell 1981, 13, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Binnington, K.C. Sequential Changes in Salivary Gland Structure during Attachment and Feeding of the Cattle Tick, Boophilus microplus. Int. J. Parasitol. 1978, 8, 97–115. [Google Scholar] [CrossRef]
- Coons, L.B.; Kaufman, W.R. Evidence That Developmental Changes in Type III Acini in the Tick Amblyomma hebraeum (Acari: Ixodidae) Are Initiated by a Hemolymph-Borne Factor. Exp. Appl. Acarol. 1988, 4, 117–139. [Google Scholar] [CrossRef]
- Coons, L.B.; Lessman, C.A.; Ward, M.W.; Berg, R.H.; Lamoreaux, W.J. Evidence of a Myoepithelial Cell in Tick Salivary Glands. Int. J. Parasitol. 1994, 24, 551–562. [Google Scholar] [CrossRef]
- Coons, L.B.; Roshdy, M.A. Fine Structure of the Salivary Glands of Unfed Male Dermacentor variabilis (Say) (Ixodoidea: Ixodidae). J. Parasitol. 1973, 59, 900–912. [Google Scholar] [CrossRef]
- Kaufman, W.R.; Aeschlimann, A.A.; Diehl, P.A. Regulation of Body Volume by Salivation in a Tick Challenged with Fluid Loads. Am. J. Physiol. 1980, 238, R102–R112. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, P.J.; Kaufman, W.R. Potentiation of Salivary Fluid Secretion in Ixodid Ticks: A New Receptor System for Gamma-Aminobutyric Acid. Can. J. Physiol. Pharmacol. 1986, 64, 1119–1126. [Google Scholar] [CrossRef]
- Kaufman, W.R.; Sloley, B.D.; Tatchell, R.J.; Zbitnew, G.L.; Diefenbach, T.J.; Goldberg, J.I. Quantification and Cellular Localization of Dopamine in the Salivary Gland of the Ixodid Tick Amblyomma hebraeum. Exp. Appl. Acarol. 1999, 23, 251–265. [Google Scholar] [CrossRef]
- Hill, C.A.; Wikel, S.K. The Ixodes Scapularis Genome Project: An Opportunity for Advancing Tick Research. Trends Parasitol. 2005, 21, 151–153. [Google Scholar] [CrossRef]
- Gulia-Nuss, M.; Nuss, A.B.; Meyer, J.M.; Sonenshine, D.E.; Roe, R.M.; Waterhouse, R.M.; Sattelle, D.B.; De La Fuente, J.; Ribeiro, J.M.; Megy, K.; et al. Genomic Insights into the Ixodes Scapularis Tick Vector of Lyme Disease. Nat. Commun. 2016, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Šimo, L.; Koči, J.; Žitňan, D.; Park, Y. Evidence for D1 Dopamine Receptor Activation by a Paracrine Signal of Dopamine in Tick Salivary Glands. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Koči, J.; Šimo, L.; Park, Y. Autocrine/Paracrine Dopamine in the Salivary Glands of the Blacklegged Tick Ixodes scapularis. J. Insect Physiol. 2014, 62, 39–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimo, L.; Koči, J.; Kim, D.; Park, Y. Invertebrate Specific D1-like Dopamine Receptor in Control of Salivary Glands in the Black-Legged Tick Ixodes scapularis. J. Comp. Neurol. 2014, 522, 2038–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Šimo, L.; Park, Y. Orchestration of Salivary Secretion Mediated by Two Different Dopamine Receptors in the Blacklegged Tick Ixodes scapularis. J. Exp. Bio. 2014, 217, 3656–3663. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Urban, J.; Boyle, D.L.; Park, Y. Multiple Functions of Na/K-ATPase in Dopamine-Induced Salivation of the Blacklegged Tick, Ixodes scapularis. Sci. Rep. 2016, 6, 21047. [Google Scholar] [CrossRef] [Green Version]
- Needham, G.R.; Rosell, R.; Greenwald, L.; Coons, L.B. Ultrastructure of Type-I Salivary-Gland Acini in Four Species of Ticks and the Influence of Hydration States on the Type-I Acini of Amblyomma americanum. Exp. Appl. Acarol. 1990, 10, 83–104. [Google Scholar] [CrossRef]
- Rudolph, D.; Knülle, W. Site and Mechanism of Water Vapour Uptake from the Atmosphere in Ixodid Ticks. Nature 1974, 249, 84–85. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, D.; Knulle, W. Uptake of Water Vapour from the Air: Process, Site and Mechanism in Ticks. In Comparative Physiology: Water Ions and Fluid Mechanics; Schmidt-Nielsen, K., Bolis, L., Maddrell, S.H.P., Eds.; Cambridge University Press: Cambridge, UK, 1978; pp. 97–113. [Google Scholar]
- Kim, D.; Maldonado-Ruiz, P.; Zurek, L.; Park, Y. Water Absorption through Salivary Gland Type I Acini in the Blacklegged Tick, Ixodes scapularis. PeerJ 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Šimo, L.; Vancová, M.; Urban, J.; Park, Y. Neural and Endocrine Regulation of Osmoregulatory Organs in Tick: Recent Discoveries and Implications. Gen. Comp. Endocrinol. 2019, 278, 42–49. [Google Scholar] [CrossRef]
- Mateos-Hernandéz, L.; Defaye, B.; Vancová, M.; Hajdusek, O.; Sima, R.; Park, Y.; Attoui, H.; Šimo, L. Cholinergic Axons Regulate Type I Acini in Salivary Glands of Ixodes ricinus and Ixodes scapularis Ticks. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Šimo, L.; Slovák, M.; Park, Y.; Žitňan, D. Identification of a Complex Peptidergic Neuroendocrine Network in the Hard Tick, Rhipicephalus appendiculatus. Cell Tissue Res. 2008, 335, 639–655. [Google Scholar] [CrossRef] [Green Version]
- Šimo, L.; Žitňan, D.; Park, Y. Two Novel Neuropeptides in Innervation of the Salivary Glands of the Black-Legged Tick, Ixodes scapularis: Myoinhibitory Peptide and SIFamide. J. Comp. Neurol. 2009, 517, 551–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimo, L.; Žitňan, D.; Park, Y. Neural Control of Salivary Glands in Ixodid Ticks. J. Insect Physiol. 2012, 58, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimo, L.; Daniel, S.; Park, Y.; Žitňan, D. The Nervous and Sensory Systems: Structure, Function, Proteomics and Genomics. In Biology of Ticks; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: New York, NZ, USA, 2014; Volume 1, pp. 309–367. [Google Scholar]
- Roller, L.; Šimo, L.; Mizoguchi, L.; Slovák, M.; Park, Y.; Žitňan, D. Orcokinin-like Immunoreactivity in Central Neurons Innervating the Salivary Glands and Hindgut of Ixodid Ticks. Cell Tissue Res. 2015, 360, 209–222. [Google Scholar] [CrossRef]
- Kim, D.; Šimo, L.; Park, Y. Molecular Characterization of Neuropeptide Elevenin and Two Elevenin Receptors, IsElevR1 and IsElevR2, from the Blacklegged Tick, Ixodes scapularis. Insect Biochem. Molec. 2018, 101, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Šimo, L.; Koči, J.; Park, Y. Receptors for the Neuropeptides, Myoinhibitory Peptide and SIFamide, in Control of the Salivary Glands of the Blacklegged Tick Ixodes scapularis. Insect Biochem. Mol. Biol. 2013, 43, 376–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vancová, M.; Bílý, T.; Nebesářová, J.; Grubhoffer, L.; Bonnet, S.; Park, Y.; Šimo, L. Ultrastructural Mapping of Salivary Gland Innervation in the Tick Ixodes ricinus. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Šimo, L.; Kazimirova, M.; Richardson, J.; Bonnet, S.I. The Essential Role of Tick Salivary Glands and Saliva in Tick Feeding and Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7, 281. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šimo, L. 50 Years since Kaufman and Phillips’ Groundbreaking Trilogy Elucidating Ion and Water Homeostasis in Ixodid Ticks. Pathogens 2023, 12, 385. https://doi.org/10.3390/pathogens12030385
Šimo L. 50 Years since Kaufman and Phillips’ Groundbreaking Trilogy Elucidating Ion and Water Homeostasis in Ixodid Ticks. Pathogens. 2023; 12(3):385. https://doi.org/10.3390/pathogens12030385
Chicago/Turabian StyleŠimo, Ladislav. 2023. "50 Years since Kaufman and Phillips’ Groundbreaking Trilogy Elucidating Ion and Water Homeostasis in Ixodid Ticks" Pathogens 12, no. 3: 385. https://doi.org/10.3390/pathogens12030385