
Putrescine Detected in Strains of Staphylococcus aureus

Abstract
:1. Introduction
2. Materials and Methods
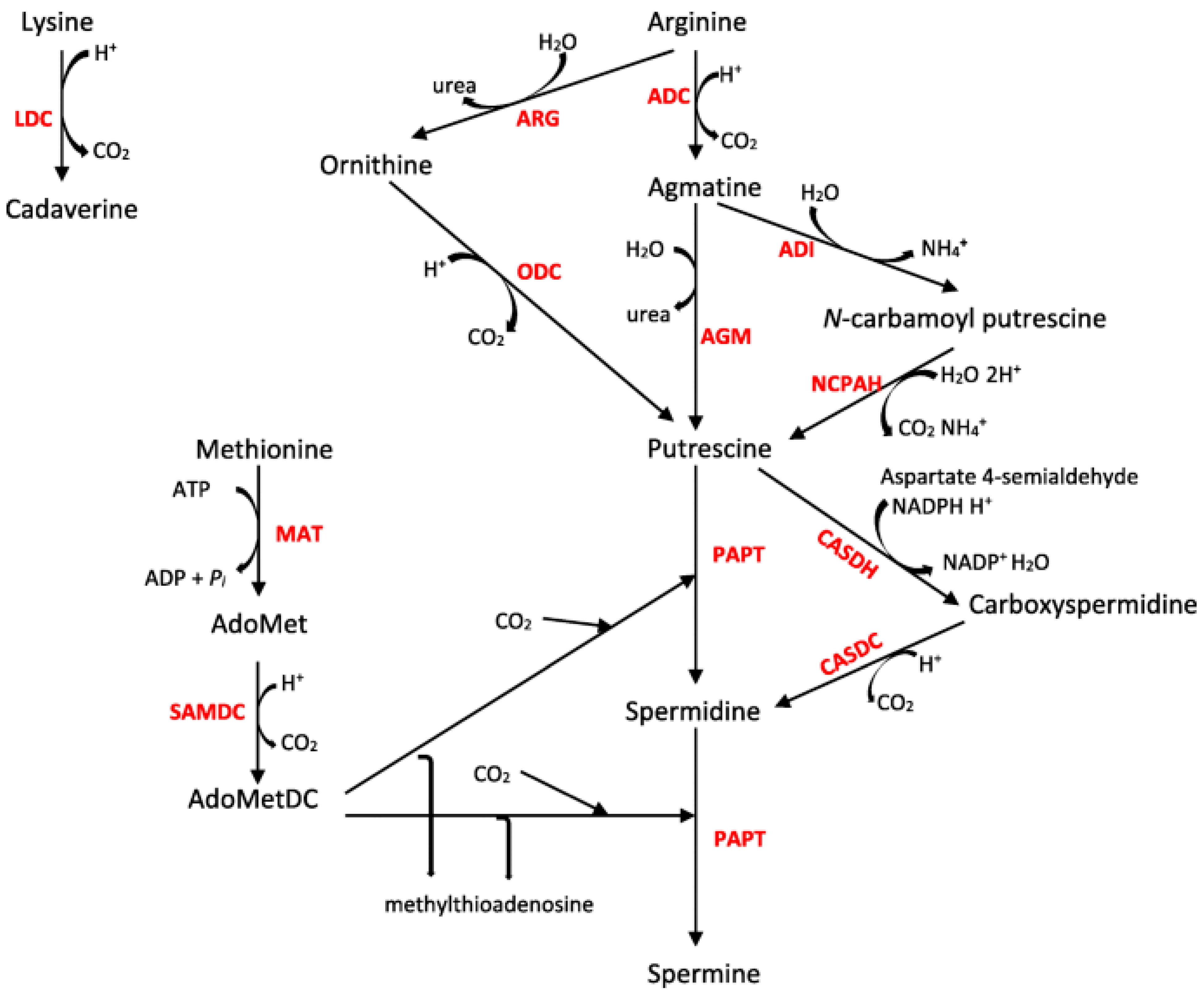
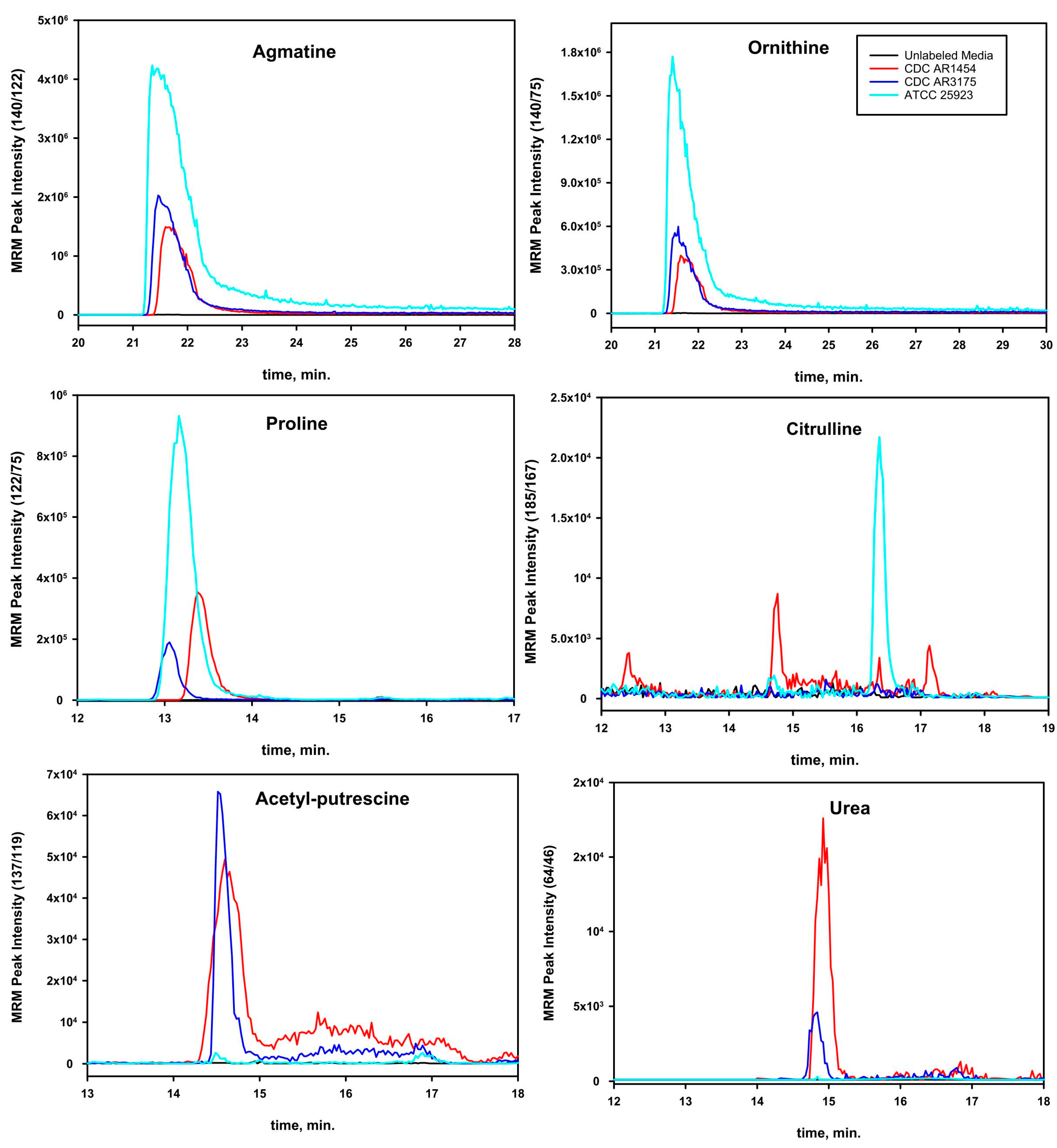
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Igarashi, K.; Kashiwag, K. The Functional Role of Polyamines in Eukaryotic Cells. Intl. J. Biochem. Cell Biol. 2018, 107, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Coton, E.; Mulder, N.; Coton, M.; Pochet, S.; Trip, H.; Lolkema, J.S. Origin of the Putrescine-Producing Ability of the Coagulase-Negative Bacterium Staphylococcus epidermis 2015B. Appl. Environ. Microbiol. 2010, 76, 5570–5576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, G.S.; Spontak, J.S.; Klapper, D.G.; Richardson, A.R. ACME Encoded speG Abrogates the Unique Hypersensitivity of Staphylococcus aureus to Exogenous Polyamines. Mol. Microbiol. 2011, 82, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Maezato, Y.; Kim, S.H.; Kurihara, S.; Liang, J.; Michael, A.J. Polyamine Independent Growth and Biofilm Formation, and Functional Spermidine/Spermine N-acetyltransferases in Staphylococcus aureus and Enterococcus faecalis. Mol. Microbiol. 2018, 111, 159–175. [Google Scholar] [CrossRef] [Green Version]
- El-Halfawy, O.; Valvano, M.A. Putrescine Reduces Antibiotic-Induced Oxidative Stress as a Mechanism of Modulation of Antibiotic Resistance in Burkholderia cenocepacia. Antimicrob. Agents Chemother. 2014, 58, 4162–4171. [Google Scholar] [CrossRef] [Green Version]
- Banerji, R.; Kanojiya, P.; Patil, A.; Saroj, S.D. Polyamines in the Virulence of Bacterial Pathogens of Respiratory Tract. Mol. Oral Microbiol. 2021, 36, 1–11. [Google Scholar] [CrossRef]
- Miller, R.D.; Fung, D.Y.C. Amino Acid Requirements for the Production of Enterotoxin B by Staphylococcus aureus S-6 in a Chemically Defined Medium. Appl. Microbiol. 1973, 25, 800–806. [Google Scholar] [CrossRef]
- Carreau, A.; Hafny-Rahbi, B.E.; Matejuk, A.; Grillon, C.; Kleda, C. Why Is the Partial Oxygen Pressure of Human Tissues a Crucial Parameter? Small Molecules and Hypoxia. J. Cell. Mol. Med. 2011, 15, 1239–1253. [Google Scholar] [CrossRef] [Green Version]
- Yarwood, J.M.; Schlievert, P.M. Oxygen and Carbon Dioxide Regulation of Toxic Shock Syndrome Toxin 1 Production by Staphylococcus aureus MN8. J. Clin. Microbiol. 2000, 38, 1797–1803. [Google Scholar] [CrossRef] [Green Version]
- George, S.E.; Hrubesch, J.; Breuing, I.; Vetter, N.; Korn, N.; Hennemann, K.; Bleul, L.; Willmann, M.; Ebner, P.; Götz, F.; et al. Oxidative Stress Drives the Selection of Quorum Sensing Mutants in the Staphylococcus aureus population. Proc. Natl. Acad. Sci. USA 2019, 116, 19145–19154. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.C. Polyamines in Soybeans. Plant Physiol. 1972, 50, 152–156. [Google Scholar] [CrossRef] [Green Version]
- Hanfrey, C.; Sommer, S.; Mayer, M.J.; Burtin, D.; Michael, A.J. Araidopsis Polyamine Biosynthesis: Absence of Ornithine Decarboxylase and the Mechanism of Arginine Decarboxylase Activity. Plant J. 2001, 27, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.; Santiago, J.C.; Rodriguez-Paez, L.; Ibanez, M.; Baeza, I. Synthesis of Putrescine Under Possible Primitive Earth Conditions. Origin Life Evol. Biosph. 1991, 21, 145–156. [Google Scholar] [CrossRef]
- Guerra, P.R.; Herrero-Fresno, A.; Ladero, V.; Pires dos Santos, T.; Spiegelhauer, M.R.; Jelsbak, L.; Olsen, J.E. Putrescine Biosynthesis and Export Genes are Essential for Normal Growth of Avian Pathogenic Escherichia coli. BMC Microbiol. 2018, 18, 226–237. [Google Scholar] [CrossRef]
- Antti, H.; Fahlgren, A.; Näsström, E.; Kouremenos, K.; Sundén-Cullberg, J.; Guo, Y.; Moritz, T.; Wolf-Watz, H.; Johansson, A.; Fallman, M. Metabolic Profiling for Detection of Staphylococcus aureus Infection and Antibiotic Resistance. PLoS ONE 2013, 8, e56971. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Zhang, S.; Chen, J.; Han, B. Metabolic Profiling of Staphylococcus aureus Cultivated Under Aerobic and Anaerobic Conditions with 1H NMR-based Nontargeted Analysis. Can. J. Microbiol. 2012, 58, 709–718. [Google Scholar] [CrossRef]
- Ammons, M.C.B.; Tripet, B.P.; Carlson, R.P.; Kirker, K.R.; Gross, M.A.; Stanisich, J.J.; Copie, V. Quantitative NMR Metabolite Profiling of Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus Discriminates Between Biofilm and Planktonic Phenotypes. J. Proteome Res. 2014, 13, 2973–2985. [Google Scholar] [CrossRef] [Green Version]
- Freudenberg, R.A.; Wittemeier, L.; Einhaus, A.; Baier, T.; Kruse, O. Advanced Pathway Engineering for Phototrophic Putrescine Production. Plant Biotech. J. 2022, 20, 1968–1982. [Google Scholar] [CrossRef]
- Rosenthal, S.M.; Dubin, D.T. Metabolism of Polyamines by Staphylococcus. J. Bacteriol. 1962, 84, 859–863. [Google Scholar] [CrossRef] [Green Version]
- Michael, A.J. Polyamines in Eukaryotes, Bacteria, and Archaea. J. Biol. Chem. 2016, 291, 14896–14903. [Google Scholar] [CrossRef] [Green Version]
- Lodish, H.; Berk, A.; Zipursky, S.L.; Matsudaira, P.; Baltimore, D.; Darnell, J. Molecular Cell Biology, 4th ed.; W.H. Freeman and Company: New York, NY, USA, 2000. [Google Scholar] [CrossRef]
- Kong, X.; Wang, X.; Yin, Y.; Li, X.; Gao, H.; Bazer, F.W.; Wu, G. Putrescine Stimulates the mTOR Signaling Pathway and Protein Synthesis in Porcine Trophectoderm Cells. Biol. Reprod. 2014, 91, 106. [Google Scholar] [CrossRef] [PubMed]
- Pollitt, E.J.G.; Szkuta, P.T.; Burns, N.; Foster, S.J. Staphylococcus aureus infection dynamics. PLoS Pathog. 2018, 14, e1007112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.; Wang, Q.; Li, Y.; Liang, Z.; Xu, L.; Zhou, J.; Cui, Z.; Zhang, L.-H. Putrescine Is an Intraspecies and Interkingdom Cell-Cell Communication Signal Modulating the Virulence of Dickeya zeae. Front. Microbiol. 2019, 10, 1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegg, A.E.; Casero, R.A. Current Status of the Polyamine Research Field. Methods Mol. Biol. 2011, 720, 3–35. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.; Mandal, A.; Johansson, H.E.; Orjalo, A.V.; Park, M.H. Depletion of Cellular Polyamines, Spermidine, and Spermine, Cause a Total Arrest in Translation and Growth in Mammalian Cells. Proc. Natl. Acad. Sci. USA 2013, 110, 2169–2174. [Google Scholar] [CrossRef] [Green Version]
- Nanduri, B.; Swiatlo, E. The expansive effects of polyamines on the metabolism and virulence of Streptococcus pneumoniae. Pneumonia 2021, 13, 4. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strain | Putrescine | Putrescine (nM) | N-Acetyl-Putrescine | Acetyl Putrescine (nM) |
|---|---|---|---|---|
| ATCC 25923 | 4.17 ± 3.03 | 0.4 | 17.2 ± 15.14 | 1.65 |
| CDC AR1454 | 2.66 ± 0.91 b | 6 | 12.59 ± 11.09 b | 28.4 |
| CDC AR3175 | 1.78 ± 0.58 | 3.95 | --- | |
| CDC AR7067 | --- | --- | 149.98 ± 123.37 a | 14.4 |
| Strain | Antibiotic Resistance | Hemolytic Activity (24 h) | Concentration CFU/mL |
|---|---|---|---|
| ATCC BAA-44 | Multiple 1 | None | 3.69 × 109 |
| NCBI COL-S | None | None | 1.7 × 109 |
| ATCC 43300 | Methicillin, Oxacillin | None | 2 × 109 |
| ATCC 25923 | Cefoxitin, Penicillin, Mupirocin | None | 1.8 × 109 |
| CDC AR1454 | Oxacillin, Penicillin, Erythromycin | None | 4.35 × 109 |
| CDC AR2789 | Oxacillin, Penicillin, Clindamycin, Erythromycin, Levofloxacin | None | 2.9 × 109 |
| CDC AR3175 | Oxacillin, Penicillin, Clindamycin, Erythromycin | None | 2.87 × 109 |
| CDC AR42208 | Oxacillin, Penicillin, Erythromycin, Levofloxacin | Some | 1.3 × 109 |
| CDC AR 5738 | Oxacillin, Penicillin, Erythromycin | Some | 2.21 × 109 |
| CDC AR6498 | Oxacillin, Penicillin, Clindamycin, Erythromycin, Levofloxacin | None | 1.9 × 109 |
| CDC AR7067 | Oxacillin, Penicillin, Erythromycin, Levofloxacin | Some | 2.23 × 109 |
| Baylor MRSA USA300 | Erythromycin, Vancomycin, Methicillin | Strong | 2.7 × 109 |
| GENE | 25923 | 43300 |
|---|---|---|
| Hypothetical protein 132469..132870 | 1 | 0 |
| Hypothetical protein 135487..136044 | 1 | 2 |
| Hypothetical protein 156985..157179 | 2 | 0 |
| Hypothetical protein 157404..158372 | 2 | 0 |
| Hypothetical protein 158649..159218 | 3 | 2 |
| Hypothetical protein 159559..160086 | 4 | 3 |
| Hypothetical protein 160137..160355 | 2 | 1 |
| Hypothetical protein 160356..160952 | 3 | 2 |
| Hypothetical protein 160964..161305 | 1 | 0 |
| Hypothetical protein 161652..162293 | 3 | 2 |
| Hypothetical protein 162290..162574 | 1 | 2 |
| Hypothetical protein 162576..162938 | 2 | 3 |
| Hypothetical protein 164721..165590 | 1 | 0 |
| Hypothetical protein 165974..166183 | 2 | 0 |
| Hypothetical protein 166176..166322 | 1 | 0 |
| Hypothetical protein 166650..167291 | 1 | 0 |
| Hypothetical protein 167295..167507 | 3 | 2 |
| Hypothetical protein 167661..168323 | 1 | 0 |
| Hypothetical protein 177188..177328 | 3 | 4 |
| Hypothetical protein 434789..435115 | 1 | 2 |
| Hypothetical protein 460317..460508 | 1 | 0 |
| Hypothetical protein 490900..491820 | 1 | 2 |
| Hypothetical protein 513477..515150 | 2 | 3 |
| Hypothetical protein 637833..638594 | 1 | 2 |
| Hypothetical protein 689131..690099 | 2 | 1 |
| Hypothetical protein 690618..691028 | 1 | 3 |
| Hypothetical protein 734010..734387 | 2 | 6 |
| Hypothetical protein 734404..735225 | 2 | 3 |
| Hypothetical protein 945446..945742 | 1 | 0 |
| Hypothetical protein 992307..993047 | 1 | 2 |
| Hypothetical protein 1004508..1005284 | 3 | 2 |
| Hypothetical protein 1182854..1183327 | 2 | 1 |
| Hypothetical protein 1183342..1183716 | 2 | 1 |
| Hypothetical protein 1183734..1184123 | 2 | 1 |
| Hypothetical protein 1184125..1184517 | 1 | 0 |
| Hypothetical protein 1750363..1750452 | 5 | 4 |
| Hypothetical protein 1254171..1255004 | 1 | 0 |
| Hypothetical protein 1256675..1257370 | 1 | 0 |
| Hypothetical protein 1257514..1257726 | 3 | 2 |
| Hypothetical protein 1257727..1257999 | 1 | 0 |
| Hypothetical protein 1258150..1258359 | 2 | 0 |
| Hypothetical protein 1258750..1259619 | 2 | 3 |
| Hypothetical protein 1259633..1261342 | 1 | 0 |
| Hypothetical protein 1262031..1262672 | 3 | 2 |
| Hypothetical protein 1263189..1263530 | 2 | 3 |
| Hypothetical protein 1263542..1264138 | 3 | 2 |
| Hypothetical protein 1264139..1264357 | 2 | 1 |
| Hypothetical protein 1264408..1264935 | 4 | 3 |
| Hypothetical protein 1265276..1265845 | 3 | 2 |
| Hypothetical protein 1266122..1267090 | 2 | 0 |
| Hypothetical protein 1267386..1267667 | 1 | 0 |
| Hypothetical protein 1267785..1268297 | 1 | 0 |
| Hypothetical protein 1274829..1275455 | 1 | 2 |
| Hypothetical protein 1488328..1488564 | 2 | 1 |
| Hypothetical protein 1521968..1522474 | 3 | 4 |
| Hypothetical protein 1587960..1588349 | 4 | 6 |
| Hypothetical protein 1653173..1654051 | 4 | 3 |
| Hypothetical protein 1744950..1745777 | 2 | 1 |
| Hypothetical protein 1850234..1850428 | 1 | 0 |
| Hypothetical protein 1975041..1976078 | 4 | 6 |
| Hypothetical protein 2009292..2009777 | 1 | 0 |
| Hypothetical protein 2020584..2021309 | 3 | 2 |
| Hypothetical protein 2021419..2022144 | 2 | 1 |
| Hypothetical protein 2187375..2187581 | 2 | 3 |
| Hypothetical protein 2197213..2197338 | 1 | 0 |
| Hypothetical protein 2268153..2268722 | 1 | 0 |
| Hypothetical protein 2292291..2292692 | 1 | 2 |
| Hypothetical protein 2314681..2314902 | 2 | 3 |
| Hypothetical protein 2321684..2322388 | 6 | 5 |
| Hypothetical protein 2413521..2414975 | 2 | 1 |
| Hypothetical protein 2414986..2415288 | 2 | 1 |
| Hypothetical protein 2415414..2415788 | 1 | 0 |
| Hypothetical protein 2420121..2421611 | 2 | 1 |
| Hypothetical protein 2421611..2426260 | 1 | 0 |
| Hypothetical protein 2426498..2426944 | 1 | 0 |
| Hypothetical protein 2427009..2427962 | 5 | 3 |
| Hypothetical protein 2427963..2428343 | 2 | 1 |
| Hypothetical protein 2428340..2428717 | 1 | 0 |
| Hypothetical protein 2428717..2429052 | 1 | 0 |
| Hypothetical protein 2429039..2429371 | 1 | 0 |
| Hypothetical protein 2429380..2429538 | 2 | 1 |
| Hypothetical protein 2429574..2430821 | 2 | 1 |
| Hypothetical protein 2430909..2431493 | 1 | 0 |
| Hypothetical protein 2431486..2432736 | 1 | 0 |
| Hypothetical protein 2432742..2432981 | 1 | 0 |
| Hypothetical protein 2432956..2434650 | 1 | 0 |
| Hypothetical protein 2434653..2435120 | 2 | 1 |
| Hypothetical protein 2435250..2435603 | 1 | 0 |
| Hypothetical protein 2435610..2436062 | 1 | 0 |
| Hypothetical protein 2436177..2436611 | 1 | 0 |
| Hypothetical protein 2437643..2437795 | 2 | 1 |
| Hypothetical protein 2437795..2437995 | 1 | 0 |
| Hypothetical protein 2438155..2438400 | 1 | 0 |
| Hypothetical protein 2439228..2439476 | 2 | 1 |
| Hypothetical protein 2439477..2439836 | 1 | 0 |
| Hypothetical protein 2439837..2440022 | 1 | 0 |
| Hypothetical protein 2440027..2440431 | 1 | 0 |
| Hypothetical protein 2440441..2440662 | 1 | 0 |
| Hypothetical protein 2440675..2440833 | 2 | 1 |
| Hypothetical protein 2440827..2441606 | 1 | 0 |
| Hypothetical protein 2441616..2442386 | 1 | 0 |
| Hypothetical protein 2442452..2442733 | 1 | 0 |
| Hypothetical protein 2442872..2443543 | 1 | 0 |
| Hypothetical protein 2444140..2444919 | 1 | 0 |
| Hypothetical protein 2444912..2445133 | 1 | 0 |
| Hypothetical protein 2445143..2445403 | 1 | 0 |
| Hypothetical protein 2445814..2445975 | 1 | 0 |
| Hypothetical protein 2446877..2447104 | 1 | 0 |
| Hypothetical protein 2447106..2447297 | 1 | 0 |
| Hypothetical protein 2447354..2447893 | 1 | 0 |
| Hypothetical protein 2450801..2450926 | 1 | 0 |
| Strain | Transport | Ornithine Decarboxylase | Agmatinase | Agmatine Deiminase | N-Carbamoyl- Putrescine Amidohydrolase | Arginine Decarboxylase |
|---|---|---|---|---|---|---|
| Streptococcus pyogenes | ||||||
| NCTC12064 | Spermidine/putrescine transport permease | No | No | No | No | No |
| HKU488 | Spermidine/putrescine transport permease | No | No | No | No | No |
| MGAS29326 | Spermidine/putrescine transport permease | No | No | No | No | No |
| TJ11-001 | No transport | No | No | No | No | No |
| BSAC_bs1388 | ABC transporter permease | No | No | No | No | No |
| ABC221 | ABC transporter permease | No | No | No | No | No |
| 37-97S | ABC transporter permease | No | No | No | No | No |
| NCTC8324 | Spermidine/putrescine transport permease | No | No | No | No | No |
| HKU419 | No transport | No | No | No | No | No |
| MGAS10786 | ABC transporter permease | No | No | No | No | No |
| 21SPY7071 | ABC transporter permease | No | No | No | No | No |
| PS003 | No transport | No | No | No | No | No |
| Pseudomonas aeruginosa | ||||||
| Pa58 | Spermidine/putrescine ABC transporter | Yes | Yes | No | Yes | Yes |
| AR_0353 | Polyamine antiporter | No | Yes | Yes | Yes | Yes |
| AG1 | Putrescine ABC transporter | No | No | Yes | Yes | Yes |
| PALA9 | Spermidine/putrescine transport system permease | Yes | No | Yes | No | Yes |
| WTJH36 | Spermidine/putrescine ABC transporter | No | Yes | Yes | Yes | Yes |
| H05 | Spermidine/putrescine ABC transporter | No | Yes | Yes | Yes | Yes |
| Bacillus subtilis | ||||||
| SRCM102754 | Putative ABC transporter ATP-binding protein, ABC transporter permease | No | Yes | No | No | Yes |
| PRO112 | ABC transporter permease, ABC transporter ATP-binding protein | No | Yes | No | No | Yes |
| ms-2 | No transport | No | No | No | No | No |
| SRCM103581 | ABC transporter permease | No | Yes | No | No | No |
| 29R7-12 | Spermidine/putrescine ABC transporter | No | Yes | No | No | Yes |
| TR21 | ABC transporter permease | No | Yes | No | No | Yes |
| ATC-3 | ABC transporter permease | No | Yes | No | No | Yes |
| MB9_B4 | ABC transporter permease | No | Yes | No | No | Yes |
| Staphylococcus aureus | ||||||
| ATCC BAA-44 | Putrescine export system ATP-binding protein | No | No | No | No | Yes |
| ATCC 6538 | Putrescine export system ATP-binding protein | No | No | No | No | Yes |
| ATCC 9144 | Putrescine export system ATP-binding protein | No | No | No | No | Yes |
| ATCC 12600 | Putrescine export system ATP-binding protein | No | No | No | No | Yes |
| ATCC BAA-1680 | Putrescine export system ATP-binding protein | No | No | No | No | Yes |
| ATCC 27660 | Putrescine export system ATP-binding protein | No | No | No | No | Yes |
| ATCC25923 | Putrescine export system ATP-binding protein | No | No | No | No | Yes |
| ATCC43300 | Putrescine export system ATP-binding protein | No | No | No | No | Yes |
| KG-22 | Spermidine/putrescine ABC transporter | No | No | No | No | No |
| JK3137 | Spermidine/putrescine ABC transporter | No | No | No | No | No |
| MRSA—AMRF 5 | Spermidine/putrescine ABC transporter | No | No | No | No | No |
| AR_0216 | Spermidine/putrescine import ATP-binding protein | No | No | No | No | No |
| USA300-SUR16 | Spermidine/putrescine ABC transporter | No | No | No | No | No |
| 110900 | Spermidine/putrescine ABC transporter | No | No | No | No | No |
| CMRSA-3 | Spermidine/putrescine ABC transporter | No | No | No | No | No |
| Escherichia coli | ||||||
| 2010C-3347 | No transport | No | No | No | No | No |
| W3110 | Putrescine/proton symporter, putrescine transport protein | Yes | Yes | No | No | Yes |
| K-12 (MG1655) | Putrescine-ornithine antiporter, putrescine transporter, putative amine transport | Yes | Yes | No | No | Yes |
| 117 | Spermidine/putrescine ABC transporter, putrescine ABC transporter permease, putrescine-ornithine antiporter | Yes | Yes | No | No | Yes |
| DSM 103246 | Spermidine/putrescine ABC transporter | Yes | Yes | No | No | Yes |
| CV261 | Putrescine-ornithine antiporter, spermidine/putrescine ABC transporter, putrescine ABC transporter permease | Yes | Yes | No | No | Yes |
| STEC2018-553 | Putrescine/proton symporter, putrescine/proton symporter, putrescine ABC transporter permease, putrescine-ornithine antiporter | Yes | Yes | No | No | Yes |
| 2013C-3277 | No transport | No | No | No | No | No |
| CFSAN027350 | Putrescine-ornithine antiporter, putrescine/proton symporter, putrescine/proton symporter, spermidine/putrescine ABC transporter, putrescine ABC transporter permease | Yes | Yes | No | No | Yes |
| Ecol_AZ146 | Putrescine-ornithine antiporter | Yes | Yes | No | No | Yes |
| 2014C-3655 | No transport | No | No | No | No | No |
| Mycobacterium tuberculosis | ||||||
| SEA02010036P6C4 | ABC transporter permease | No | No | No | No | No |
| 1-0110P6c4 | ABC transporter permease | No | No | No | No | No |
| MTB2 | ABC transporter permease | No | No | No | No | No |
| H54 | Spermidine/putrescine ABC transporter, ABC transporter permease | No | No | No | No | No |
| BLR-31d | ABC transporter permease, ABC transporter permease | No | No | No | No | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seravalli, J.; Portugal, F. Putrescine Detected in Strains of Staphylococcus aureus. Pathogens 2023, 12, 881. https://doi.org/10.3390/pathogens12070881
Seravalli J, Portugal F. Putrescine Detected in Strains of Staphylococcus aureus. Pathogens. 2023; 12(7):881. https://doi.org/10.3390/pathogens12070881
Chicago/Turabian StyleSeravalli, Javier, and Frank Portugal. 2023. "Putrescine Detected in Strains of Staphylococcus aureus" Pathogens 12, no. 7: 881. https://doi.org/10.3390/pathogens12070881