
Protective Efficacy of BCG Vaccination in Calves Vaccinated at Different Ages

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Bacteria and Inoculation
2.3. Post-Mortem Examination and Bacteriology
2.4. Antigens
2.5. Tuberculin Skin Tests
2.6. Immunological Assays
2.7. Statistical Analysis
3. Results
3.1. Protection Induced by BCG Vaccination Is Not Significantly Different in Calves Vaccinated at 1 Day or 3 Weeks of Age
3.2. BCG Vaccination of Calves Reduces Bacterial Burden in Calves
3.3. Skin Test Reactivity Did Not Differ between BCG-Vaccinated and Control Calves Following M. bovis Challenge
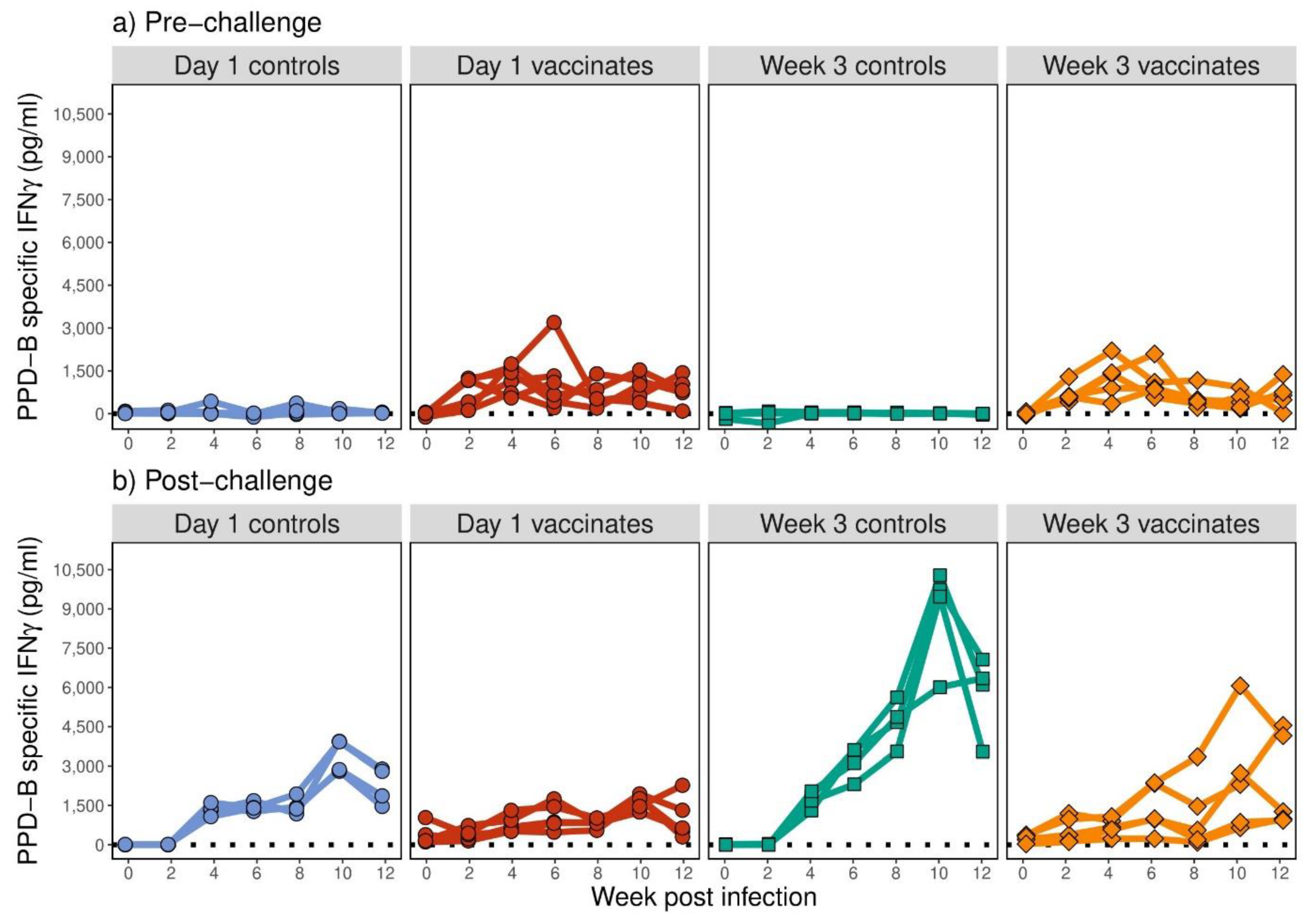
3.4. BCG Induced Similar Kinetics and Intensity of Antigen-Specific IFN-γ Responses in Calves Vaccinated at 1 Day or 3 Weeks of Age
3.5. Antigen-Specific IFN-γ Expression Post-Vaccination Correlated with Protection Whereas Expression Post-Challenge Correlated with Pathology and Bacterial Burden
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Correia, C.N.; McHugo, G.; Browne, J.A.; McLoughlin, K.E.; Nalpas, N.C.; Magee, D.A.; Whelan, A.O.; Villarreal-Ramos, B.; Vordermeier, H.M.; Gormley, E.; et al. High-resolution transcriptomics of bovine purified protein derivative-stimulated peripheral blood from cattle infected with Mycobacterium bovis across an experimental time course. Tuberculosis 2022, 136, 102235. [Google Scholar] [CrossRef]
- Olea-Popelka, F.; Muwonge, A.; Perera, A.; Dean, A.S.; Mumford, E.; Erlacher-Vindel, E.; Forcella, S.; Silk, B.J.; Ditiu, L.; El Idrissi, A.; et al. Zoonotic tuberculosis in human beings caused by Mycobacterium bovis—A call for action. Lancet Infect. Dis. 2017, 17, e21–e25. [Google Scholar] [CrossRef] [PubMed]
- De la Rua-Domenech, R.; Goodchild, A.T.; Vordermeier, H.M.; Hewinson, R.G.; Christiansen, K.H.; Clifton-Hadley, R.S. Ante mortem diagnosis of tuberculosis in cattle: A review of the tuberculin tests, γ-interferon assay and other ancillary diagnostic techniques. Res. Vet. Sci. 2006, 81, 190–210. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V. Tuberculosis: A reemerging disease at the interface of domestic animals and wildlife. In Wildlife and Emerging Zoonotic Diseases: The Biology, Circumstances and Consequences of Cross-Species Transmission; Springer: Berlin/Heidelberg, Germany, 2007; pp. 195–215. [Google Scholar]
- Bayissa, B.; Sirak, A.; Worku, A.; Zewude, A.; Zeleke, Y.; Chanyalew, M.; Gumi, B.; Berg, S.; Conlan, A.; Hewinson, R.G.; et al. Evaluation of the efficacy of BCG in protecting against contact challenge with bovine tuberculosis in holstein-friesian and zebu crossbred calves in ethiopia. Front. Vet. Sci. 2021, 8, 702402. [Google Scholar] [CrossRef] [PubMed]
- Buddle, B.M.; Vordermeier, H.M.; Chambers, M.A.; de Klerk-Lorist, L.M. Efficacy and safety of BCG vaccine for control of tuberculosis in domestic livestock and wildlife. Front. Vet. Sci. 2018, 5, 259. [Google Scholar] [CrossRef]
- Hope, J.; Thom, M.L.; McAulay, M.; Mead, E.; Vordermeier, H.M.; Clifford, D.; Hewinson, R.G.; Villarreal-Ramos, B. Identification of surrogates and correlates of protection in protective immunity against Mycobacterium bovis infection induced in neonatal calves by vaccination with M. bovis BCG Pasteur and M. bovis BCG Danish. Clin. Vaccine Immunol. 2011, 18, 373–379. [Google Scholar] [CrossRef]
- Hope, J.; Thom, M.L.; Villarreal-Ramos, B.; Vordermeier, H.M.; Hewinson, R.G.; Howard, C.J. Vaccination of neonatal calves with Mycobacterium bovis BCG induces protection against intranasal challenge with virulent M. bovis. Clin. Exp. Immunol. 2005, 139, 48–56. [Google Scholar] [CrossRef]
- Vordermeier, H.M.; Villarreal-Ramos, B.; Cockle, P.J.; McAulay, M.; Rhodes, S.G.; Thacker, T.; Gilbert, S.C.; McShane, H.; Hill, A.V.; Xing, Z.; et al. Viral booster vaccines improve Mycobacterium bovis BCG-induced protection against bovine tuberculosis. Infect. Immun. 2009, 77, 3364–3373. [Google Scholar]
- Williams, G.A.; Scott-Baird, E.; Núñez, A.; Salguero, F.J.; Wood, E.; Houghton, S.; Vordermeier, H.M. The safety of BCG vaccination in cattle: Results from good laboratory practice safety studies in calves and lactating cows. Heliyon 2022, 8, e12356. [Google Scholar] [CrossRef]
- Ábalos, P.; Valdivieso, N.; Pérez de Val, B.; Vordermeier, M.; Benavides, M.B.; Alegría-Morán, R.; Saadi, K.; Wistuba, M.; Ortega, C.; Sánchez, N.; et al. Vaccination of calves with the Mycobacterium bovis BCG strain induces protection against bovine tuberculosis in dairy herds under a natural transmission setting. Animals 2022, 12, 1083. [Google Scholar] [CrossRef]
- Buddle, B.; Wedlock, D.N.; Parlane, N.A.; Corner, L.A.; De Lisle, G.W.; Skinner, M.A. Revaccination of neonatal calves with Mycobacterium bovis BCG reduces the level of protection against bovine tuberculosis induced by a single vaccination. Infect. Immun. 2003, 71, 6411–6419. [Google Scholar] [CrossRef]
- Wedlock, D.N.; Denis, M.; Vordermeier, H.M.; Hewinson, R.G.; Buddle, B.M. Vaccination of cattle with Danish and Pasteur strains of Mycobacterium bovis BCG induce different levels of IFNγ post-vaccination, but induce similar levels of protection against bovine tuberculosis. Vet. Immunol. Immunopathol. 2007, 118, 50–58. [Google Scholar] [CrossRef]
- Siddiqui, N.; Price, S.; Hope, J. BCG vaccination of neonatal calves: Potential roles for innate immune cells in the induction of protective immunity. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 219–226. [Google Scholar] [CrossRef]
- Srinivasan, S.; Conlan, A.J.K.; Easterling, L.A.; Herrera, C.; Dandapat, P.; Veerasami, M.; Ameni, G.; Jindal, N.; Raj, G.D.; Wood, J.; et al. A meta-Analysis of the effect of Bacillus Calmette-Guérin vaccination against bovine tuberculosis: Is perfect the enemy of good? Front. Vet. Sci. 2021, 8, 637580. [Google Scholar] [CrossRef] [PubMed]
- Vordermeier, H.M.; Jones, G.J.; Buddle, B.M.; Hewinson, R.G. Development of immune-diagnostic reagents to diagnose bovine tuberculosis in cattle. Vet. Immunol. Immunopathol. 2016, 181, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Thom, M.; McAulay, M.; Vordermeier, H.M.; Clifford, D.; Hewinson, R.G.; Villarreal-Ramos, B.; Hope, J.C. Duration of immunity against Mycobacterium bovis following neonatal vaccination with bacillus Calmette-Guérin Danish: Significant protection against infection at 12, but not 24, months. Clin. Vaccine Immunol. 2012, 19, 1254–1260. [Google Scholar] [CrossRef] [PubMed]
- Bayissa, B.; Sirak, A.; Zewude, A.; Worku, A.; Gumi, B.; Berg, S.; Hewinson, R.G.; Wood, J.L.N.; Jones, G.J.; ETHICOBOTS Consortium; et al. Field evaluation of specific mycobacterial protein-based skin test for the differentiation of Mycobacterium bovis-infected and Bacillus Calmette Guerin-vaccinated crossbred cattle in Ethiopia. Transbound. Emerg. Dis. 2022, 69, e1–e9. [Google Scholar] [CrossRef]
- Srinivasan, S.; Jones, G.; Veerasami, M.; Steinbach, S.; Holder, T.; Zewude, A.; Fromsa, A.; Ameni, G.; Easterling, L.; Bakker, D.; et al. A defined antigen skin test for the diagnosis of bovine tuberculosis. Sci. Adv. 2019, 5, eaax4899. [Google Scholar] [CrossRef]
- Jones, G.J.; Konold, T.; Hurley, S.; Holder, T.; Steinbach, S.; Coad, M.; Wedlock, D.N.; Buddle, B.M.; Singh, M.; Vordermeier, H.M. Test performance data demonstrates utility of a cattle DIVA skin test reagent (DST-F) compatible with BCG vaccination. Sci. Rep. 2022, 12, 12052. [Google Scholar]
- Garnier, T.; Eiglmeier, K.; Camus, J.C.; Medina, N.; Mansoor, H.; Pryor, M.; Duthoy, S.; Grondin, S.; Lacroix, C.; Monsempe, C.; et al. The complete genome sequence of Mycobacterium bovis. Proc. Natl. Acad. Sci. USA 2003, 100, 7877–7882. [Google Scholar] [CrossRef]
- Vordermeier, H.M.; Chambers, M.A.; Cockle, P.J.; Whelan, A.O.; Simmons, J.; Hewinson, R.G. Correlation of ESAT-6-specific gamma interferon production with pathology in cattle following Mycobacterium bovis BCG vaccination against experimental bovine tuberculosis. Infect. Immun. 2002, 70, 3026–3032. [Google Scholar] [CrossRef]
- Gallagher, J.; Horwill, D. A selective oleic acid albumin agar medium for the cultivation of Mycobacterium bovis. Epidemiol. Infect. 1977, 79, 155–160. [Google Scholar]
- Morrison, W.I.; Bourne, F.J.; Cox, D.R.; Donnelly, C.A.; Gettinby, G.; McInerney, J.P.; Woodroffe, R. Pathogenesis and diagnosis of infections with Mycobacterium bovis in cattle. Independent Scientific Group on Cattle TB. Vet. Rec. 2000, 146, 236–242. [Google Scholar] [PubMed]
- Kwong, L.S.; Hope, J.C.; Thom, M.L.; Sopp, P.; Duggan, S.; Bembridge, G.P.; Howard, C.J. Development of an ELISA for bovine IL-10. Vet. Immunol. Immunopathol. 2002, 85, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Wedlock, D.N.; Skinner, M.A.; Parlane, N.A.; Vordermeier, H.M.; Hewinson, R.G.; de Lisle, G.W.; Buddle, B.M. Vaccination with DNA vaccines encoding MPB70 or MPB83 or a MPB70 DNA prime-protein boost does not protect cattle against bovine tuberculosis. Tuberculosis 2003, 83, 339–349. [Google Scholar] [CrossRef]
- Buddle, B.M.; Wards, B.J.; Aldwell, F.E.; Collins, D.M.; de Lisle, G.W. Influence of sensitisation to environmental mycobacteria on subsequent vaccination against bovine tuberculosis. Vaccine 2002, 20, 1126–1133. [Google Scholar] [CrossRef]
- Roy, A.; Eisenhut, M.; Harris, R.J.; Rodrigues, L.C.; Sridhar, S.; Habermann, S.; Snell, L.; Mangtani, P.; Adetifa, I.; Lalvani, A.; et al. Effect of BCG vaccination against Mycobacterium tuberculosis infection in children: Systematic review and meta-analysis. BMJ 2014, 349, g4643. [Google Scholar] [CrossRef]
- Hope, J.C.; Villarreal-Ramos, B. Bovine TB and the development of new vaccines. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 77–100. [Google Scholar] [CrossRef] [PubMed]
- Hope, J.C.; Vordermeier, H.M. Vaccines for bovine tuberculosis: Current views and future prospects. Expert Rev. Vaccines 2005, 4, 891–903. [Google Scholar] [CrossRef]
- Vordermeier, H.M.; Jones, G.J.; Buddle, B.M.; Hewinson, R.G.; Villarreal-Ramos, B. Bovine Tuberculosis in Cattle: Vaccines, DIVA Tests, and Host Biomarker Discovery. Annu. Rev. Anim. Biosci. 2016, 4, 87–109. [Google Scholar] [CrossRef]
- Olsen, I.; Boysen, P.; Kulberg, S.; Hope, J.C.; Jungersen, G.; Storset, A.K. Bovine NK cells can produce gamma interferon in response to the secreted mycobacterial proteins ESAT-6 and MPP14 but not in response to MPB70. Infect. Immun. 2005, 73, 5628–5635. [Google Scholar] [CrossRef]
- Sopp, P.; Howard, C.J.; Hope, J.C. Flow cytometric detection of gamma interferon can effectively discriminate Mycobacterium bovis BCG-vaccinated cattle from M. bovis-infected cattle. Clin. Vaccine Immunol. 2006, 13, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Whelan, A.O.; Coad, M.; Upadhyay, B.L.; Clifford, D.J.; Hewinson, R.G.; Vordermeier, H.M. Lack of correlation between BCG-induced tuberculin skin test sensitisation and protective immunity in cattle. Vaccine 2011, 29, 5453–5458. [Google Scholar]
- Subramanian, S.; Srinivasan, S.; Ramaiyan Selvaraju, K.; Vinoli, P.M.; Selladurai, S.; Ramasamy, B.; Kumaragurubaran, K.; Bakker, D.; Vordermeier, M.; Kapur, V.; et al. Defined Antigen Skin Test for Bovine Tuberculosis Retains Specificity on Revaccination With Bacillus Calmette–Guérin. Front. Vet. Sci. 2022, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Subramanian, S.; Shankar Balakrishnan, S.; Ramaiyan Selvaraju, K.; Manomohan, V.; Selladurai, S.; Jothivelu, M.; Kandasamy, S.; Gopal, D.R.; Kathaperumal, K.; et al. A defined antigen skin test that enables implementation of BCG vaccination for control of bovine tuberculosis: Proof of concept. Front. Vet. Sci. 2020, 7, 391. [Google Scholar] [CrossRef]
- Mills, G. Bovine TB vaccine trials move to next phase. Vet. Rec. 2023, 192, 146–147. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal | Treatment | Age at Vaccination | Sex | BCG Titre (×106) | M. bovis Titre (×104) |
|---|---|---|---|---|---|
| 201540 | BCG | 1 day | Female | 1.2 | 3.2 |
| 301548 | BCG | 1 day | Female | 0.94 | 3.2 |
| 201554 | BCG | 1 day | Male | 1.2 | 3.2 |
| 401633 | BCG | 1 day | Female | 0.95 | 3.2 |
| 601642 | BCG | 1 day | Female | 1.1 | 3.2 |
| 401549 | Control | 1 day | Male | 3.2 | |
| 301555 | Control | 1 day | Male | 3.2 | |
| 601558 | Control | 1 day | Female | 3.2 | |
| 501564 | Control | 1 day | Male | 3.2 | |
| 301583 | BCG | 3 weeks | Female | 1.2 | 5.6 |
| 201582 | BCG | 3 weeks | Male | 1.2 | 5.6 |
| 601600 | BCG | 3 weeks | Male | 1.2 | 5.6 |
| 701601 | BCG | 3 weeks | Female | 1.2 | 5.6 |
| 301604 | BCG | 3 weeks | Female | 1.2 | 5.6 |
| 101623 | Control | 3 weeks | Female | 5.6 | |
| 201624 | Control | 3 weeks | Male | 5.6 | |
| 601628 | Control | 3 weeks | Female | 5.6 | |
| 701629 | Control | 3 weeks | Male | 5.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hope, J.C.; Khalid, H.; Thom, M.L.; Howard, C.J.; Shaw, D.J. Protective Efficacy of BCG Vaccination in Calves Vaccinated at Different Ages. Pathogens 2023, 12, 789. https://doi.org/10.3390/pathogens12060789
Hope JC, Khalid H, Thom ML, Howard CJ, Shaw DJ. Protective Efficacy of BCG Vaccination in Calves Vaccinated at Different Ages. Pathogens. 2023; 12(6):789. https://doi.org/10.3390/pathogens12060789
Chicago/Turabian StyleHope, Jayne C., Hamza Khalid, Michelle L. Thom, Chris J. Howard, and Darren J. Shaw. 2023. "Protective Efficacy of BCG Vaccination in Calves Vaccinated at Different Ages" Pathogens 12, no. 6: 789. https://doi.org/10.3390/pathogens12060789