

The Low Variability of Tc24 in Trypanosoma cruzi TcI as an Advantage for Chagas Disease Prophylaxis and Diagnosis in Mexico
 , , , , , , , , , , add
Show full author list
, , , , , , , , , , add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection, Maintenance, and Proliferation of Isolates
2.2. Development of Primers
2.3. Phylogenetic Analysis
2.4. Genetic Analysis
3. Results
3.1. Identification of the Isolates
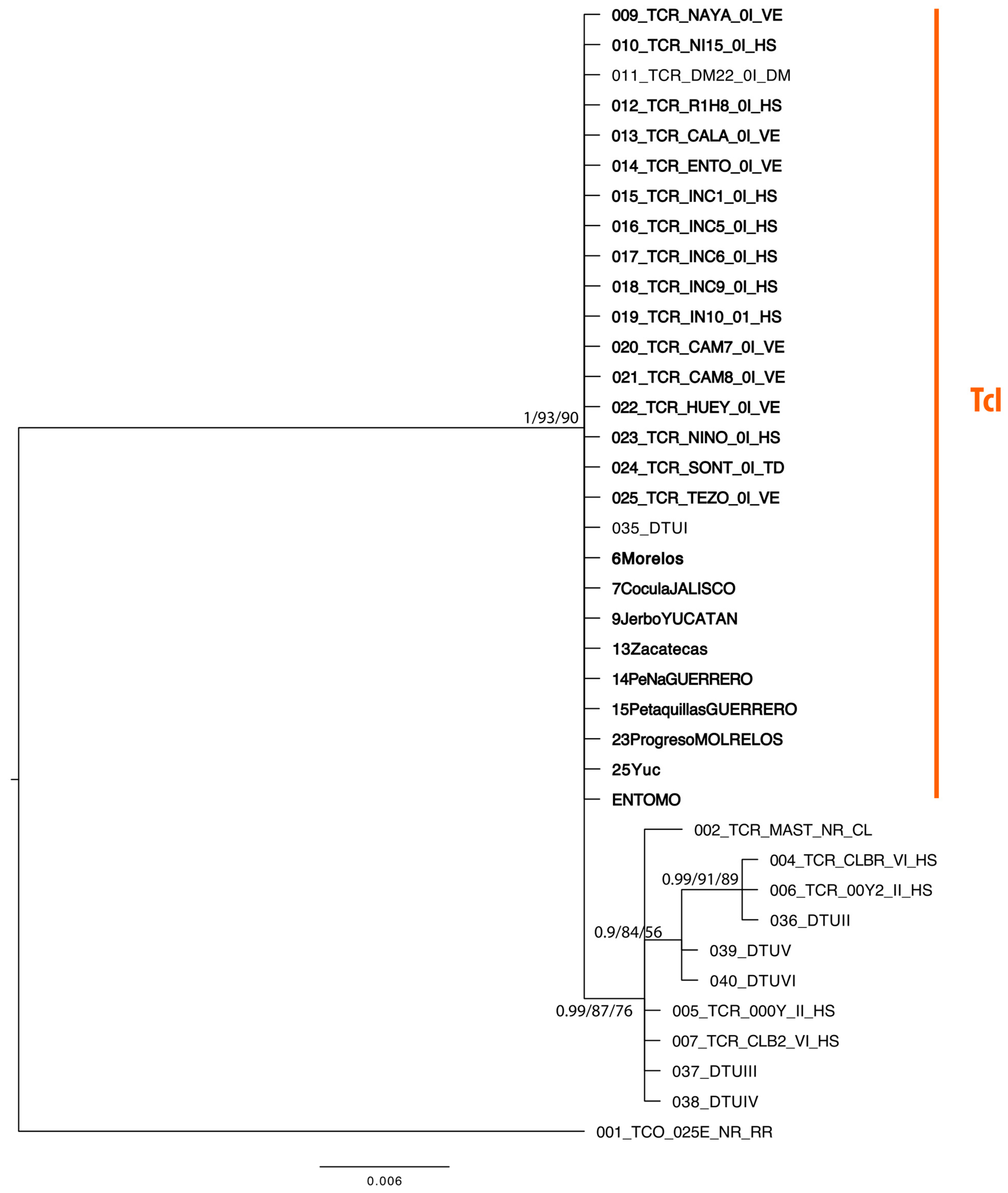
3.2. Phylogenetic Analysis
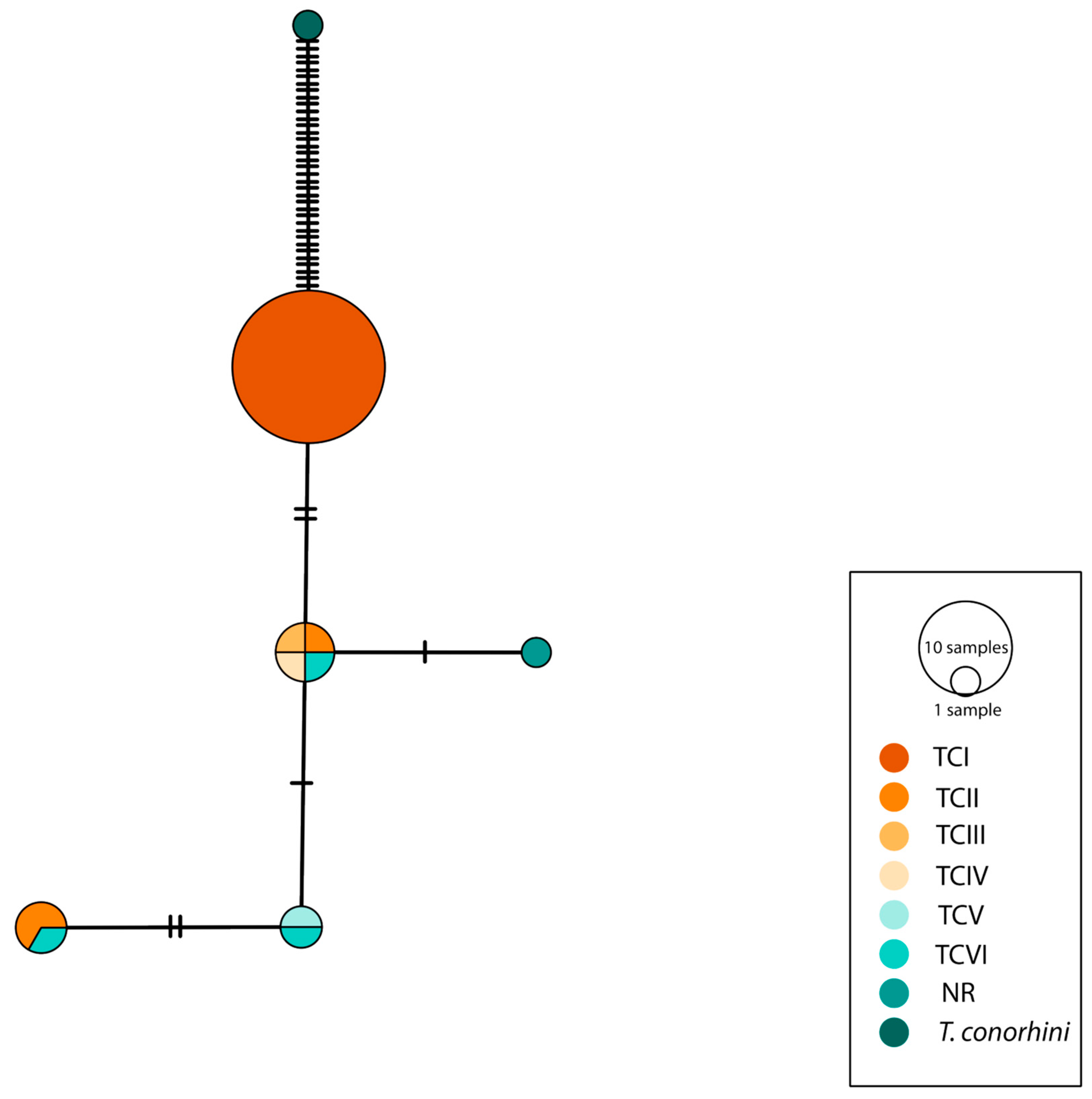
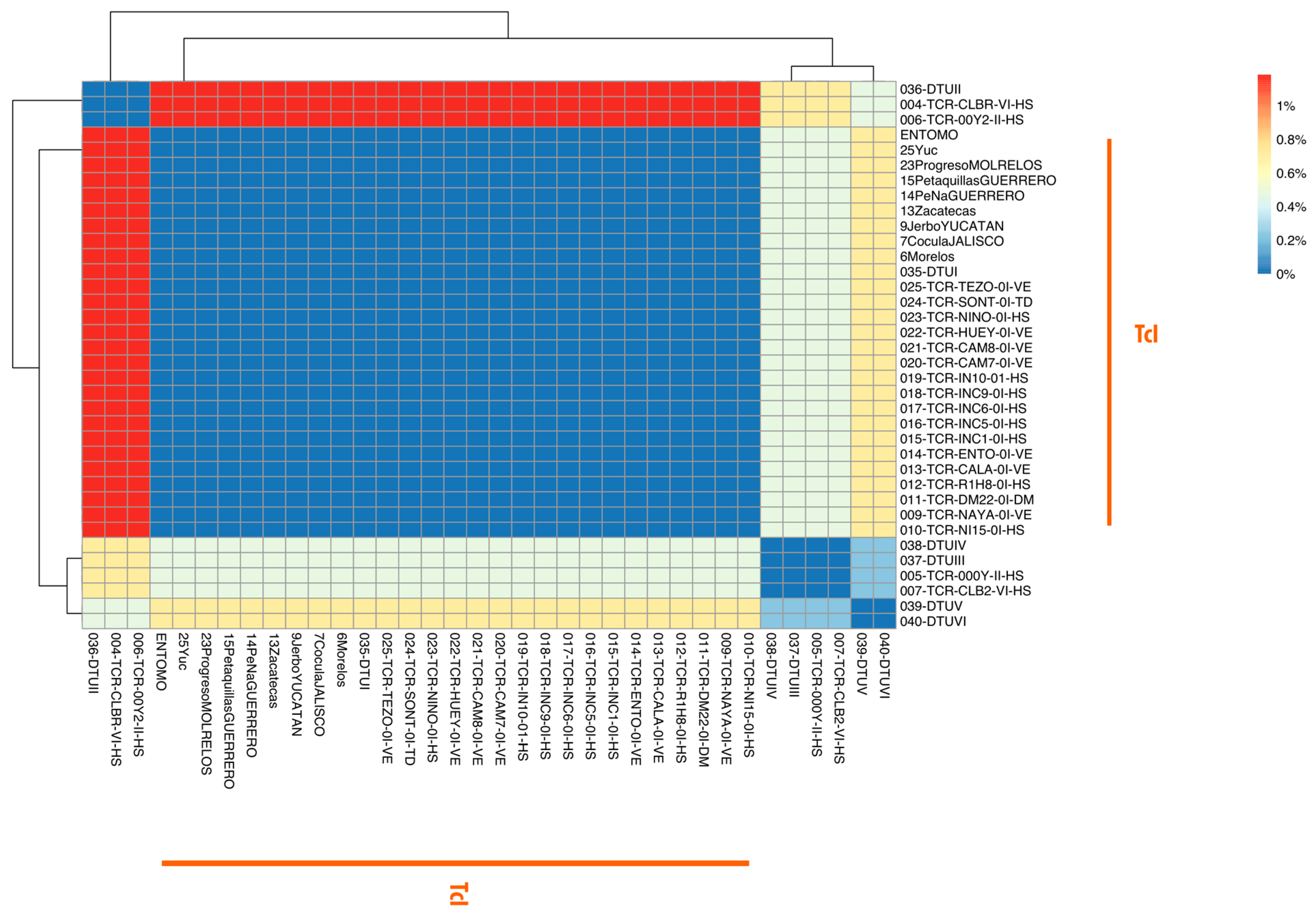
3.3. Genetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Chagas disease in Latin America: An epidemiological update based on 2010 estimates. Wkly. Epidemiol. Rec. 2015, 90, 33–43. [Google Scholar]
- Rassi, A.J.; Rassi, A.; Marin-Neto, J. Chagas disease. Lancet 2010, 375, 1388–1402. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Bacon, K.; Bottazzi, M.; Hotez, P. Global economic burden of Chagas disease: A computational simulation model. Lancet Infect. Dis. 2013, 13, 342–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Requena-Méndez, A.; Albajar-Viñas, P.; Angheben, A.; Chiodini, P.; Gascón, J.; Muñoz, J. Health policies to control Chagas disease transmission in European countries. PLoS Negl. Trop. Dis. 2014, 8, 3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angheben, A.; Boix, L.; Buonfrate, D.; Gobbi, F.; Bisoffi, Z.; Pupella, S.; Gandini, G.; Aprili, G. Chagas disease and transfusion medicine: A perspective from non-endemic countries. Blood Transfus. 2015, 13, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, J.D.; Hernández, C.; Montilla, M.; Zambrano, P.; Flórez, A.C.; Parra, E.; Cucunubá, Z.M. First report of human Trypanosoma cruzi infection attributed to TcBat genotype. Zoonoses Public Health 2014, 61, 477–479. [Google Scholar] [CrossRef]
- Zingales, B.; Andrade, S.; Briones, M.; Campbell, D.; Chiari, E.; Fernandes, O.; Guhl, F.; Lages-Silva, E.; Macedo, A.M.; Machado, C.R.; et al. A new consensus for Trypanosoma cruzi intraspecific nomenclature: Second revision meeting recommends TcI to TcVI. Mem. Inst. Oswaldo. Cruz. 2009, 104, 1051–1054. [Google Scholar] [CrossRef]
- Brenière, S.; Waleckx, E.; Barnabé, C. Over six thousand Trypanosoma cruzi strains classified into Discrete Typing Units (DTUs): Attempt at an inventory. PLoS Negl. Trop. Dis. 2016, 10, e0004792. [Google Scholar] [CrossRef] [Green Version]
- Centro Nacional de Vigilancia Epidemiológica y Control de Enfermedades (CENAVECE). Anuarios de Morbilidad Durante el Periodo 1995–2010. In Anuarios de Morbilidad, 1984–2017; Secretaría de Salud: Ciudad de Mexico, Mexico, 2022; Available online: https://www.gob.mx/salud/acciones-y-programas/anuarios-de-morbilidad-1984-a-2020 (accessed on 8 March 2021).
- SSA—Secretaría de Salud. Norma Oficial Mexicana NOM-032-SSA2-2014, Para la Vigilancia Epidemiológica, Prevención y Control de Enfermedades Transmitidas por Vector; Diario Oficial de la Federación: Ciudad de Mexico, Mexico, 2014. [Google Scholar]
- Ibarra-Cerdeña, C.; Sánchez-Cordero, V.; Townsend, P.A.; Ramsey, J. Ecology of North American Triatominae. Acta Trop. 2009, 110, 178–186. [Google Scholar] [CrossRef]
- Magallón-Gastélum, E.; Lozano-Kasten, F.; Gutierréz, M.; Flores-Pérez, A.; Sánchez, B.; Espinoza, B.; Bosseno, M.; Brenière, S. Epidemiological risk for Trypanosoma cruzi transmission by species of Phyllosoma complex in the occidental part of Mexico. Acta Trop. 2006, 97, 331–338. [Google Scholar] [CrossRef]
- Sandoval-Ruiz, C.; Cervantes-Peredo, L.; Mendoza-Palmero, F.; Ibáñez-Bernal, S. The Triatominae (Hemiptera: Heteroptera: Reduviidae) of Veracruz, Mexico: Geographic distribution, taxonomic redescriptions, and a key. Zootaxa 2012, 3487, 1–23. [Google Scholar] [CrossRef]
- Arce-Fonseca, M.; Carrillo-Sánchez, S.C.; Molina-Barrios, R.M.; Martínez-Cruz, M.; Cedillo-Cobián, J.R.; Henao-Díaz, Y.A.; Rodríguez-Morales, O. Seropositivity for Trypanosoma cruzi in domestic dogs from Sonora, Mexico. Infect. Dis. Poverty 2017, 6, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buekens, P.; Cafferata, M.; Alger, J.; Althabe, F.; Belizán, J.; Carlier, Y.; Ciganda, A.; Del Cid, J.H.; Dumonteil, E.; Gamboa-León, R.; et al. Congenital transmission of Trypanosoma cruzi in Argentina, Honduras, and Mexico: An Observational Prospective Study. Am. J. Trop. Med. Hyg. 2018, 98, 478–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montes-Rincón, L.; Galaviz-Silva, L.; Molina-Garza, Z. Anticuerpos anti-Trypanosoma cruzi en migrantes latinoamericanos en tránsito por el cruce fronterizo entre México y los Estados Unidos. Biomédica 2018, 38, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Newton-Sánchez, O.; Espinoza-Gómez, F.; Melnikov, V.; Delgado-Enciso, I.; Rojas-Larios, F.; Dumonteil, E.; Trujillo-Hernández, B.; De La Cruz-Ruíz, M. Seroprevalence of Trypanosoma cruzi (TC) and risk factors in Colima, Mexico. Gac. Med. Mex. 2017, 153, 179–184. [Google Scholar]
- Zumaya-Estrada, F.; Messenger, L.; Lopez-Ordonez, T.; Lewis, M.; Flores-Lopez, C.; Martínez-Ibarra, A.; Pennington, P.M.; Cordon-Rosales, C.; Carrasco, H.V.; Segovia, M.; et al. North American import? Charting the origins of an enigmatic Trypanosoma cruzi domestic genotype. Parasit. Vectors 2012, 5, 226. [Google Scholar] [CrossRef] [Green Version]
- Rivas-García, L.; Carballo-Amador, M.; Flores-López, C. Design of a AFLP-PCR and PCR-RFLP test that identify the majority of discrete typing units of Trypanosoma cruzi. PLoS ONE 2020, 15, e0237180. [Google Scholar] [CrossRef]
- Díaz-Valdez, J.; Martínez, I.; Rodríguez-Moreno, Á.; Gutiérrez-Granados, G.; León-Villegas, R.; Sánchez-Cordero, V.; Fraga-Nodarse, J.; Ángles-Chimal, J.; Espinoza, E. Multiple discrete typing units of Trypanosoma cruzi infect sylvatic Triatoma dimidiata and Panstrongylus rufotuberculatus in Southeast Mexico. Am. J. Trop. Med. Hyg. 2021, 105, 1042–1049. [Google Scholar] [CrossRef]
- Bosseno, M.; Barnabé, C.; Magallón-Gastélum, E.; Lozano-Kasten, F.; Ramsey, J.; Espinoza, B.; Brenière, S. Predominance of Trypanosoma cruzi lineage I in Mexico. J. Clin. Microbiol. 2002, 40, 627–632. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Guillén, M.; Bernabé, C.; Tibayrenc, M.; Zavala-Castro, J.; Totolhua, J.; Méndez-López, J.; González-Mejía, M.E.; Torres-Rasgado, E.; López-Colombo, A.; Pérez-Fuentes, R. Trypanosoma cruzi strains isolated from human, vector, and animal reservoir in the same endemic region in Mexico and typed as T. cruzi I, discrete typing unit 1 exhibit considerable biological diversity. Mem. Inst. Oswaldo. Cruz. 2006, 101, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez-Cervantes, G.; Martínez-Ibarra, A.; Nogueda-Torres, B.; López-Orduña, E.; Alonso, A.; Perea, C.; Maldonado, T.; Hernández, J.M.; León-Avila, G. Identification by Q-PCR of Trypanosoma cruzi lineage and determination of blood meal sources in triatomine gut samples in México. Parasitol. Int. 2013, 62, 36–43. [Google Scholar] [CrossRef]
- Ramos-Ligonio, A.; Torres-Montero, J.; López-Monteon, A.; Dumonteil, E. Extensive diversity of Trypanosoma cruzi discrete typing units circulating in Triatoma dimidiata from central Veracruz, Mexico. Infect. Genet. Evol. 2012, 12, 1341–1353. [Google Scholar] [CrossRef] [PubMed]
- López-Cancino, S.; Tun-Ku, E.; De la Cruz-Felix, H.K.; Ibarra-Cerdeña, C.; Izeta-Alberdi, A.; Pech-May, A.; Mazariegos-Hidalgo, C.; Valdez-Tah, A.; Ramsey, J. Landscape ecology of Trypanosoma cruzi in the southern Yucatan Peninsula. Acta Trop. 2015, 151, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Monteón, V.; Triana-Chávez, O.; Mejía-Jaramillo, A.; Pennignton, P.; Ramos-Ligonio, Á.; Acosta, K.; Lopez, R. Circulation of Tc Ia discrete type unit Trypanosoma cruzi in Yucatan Mexico. J. Parasit. Dis. 2016, 40, 550–554. [Google Scholar] [CrossRef] [Green Version]
- Pech-May, A.; Mazariegos-Hidalgo, C.; Izeta-Alberdi, A.; López-Cancino, S.; Tun-Ku, E.; De la Cruz-Félix, K.; Ibarra-Cerdeña, C.; Ittig, R.G.; Ramsey, J. Genetic variation and phylogeography of the Triatoma dimidiata complex evidence a potential center of origin and recent divergence of haplogroups having differential Trypanosoma cruzi and DTU infections. PLoS Negl. Trop. Dis. 2019, 13, e0007044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murillo-Solano, C.; Ramos-Ligonio, A.; López-Monteon, A.; Guzmán-Gómez, D.; Torres-Montero, J.; Herrera, C.; Dumonteil, E. Diversity of Trypanosoma cruzi parasites infecting Triatoma dimidiata in Central Veracruz, Mexico, and their One Health ecological interactions. Infect. Genet. Evol. 2021, 95, 105050. [Google Scholar] [CrossRef]
- Dorn, P.; McClure, A.; Gallaspy, M.; Waleckx, E.; Woods, A.; Monroy, M.; Stevens, L. The diversity of the Chagas parasite, Trypanosoma cruzi, infecting the main Central American vector, Triatoma dimidiata, from Mexico to Colombia. PLoS Negl. Trop. Dis. 2017, 11, e0005878. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Hernández, C.; Pérez, S.; Rezende-Oliveira, K.; Barbosa, C.; Lages-Silva, E.; Ramírez, L.; Ramírez, J. Evaluation of the multispecies coalescent method to explore intra- Trypanosoma cruzi I relationships and genetic diversity. Parasitology 2019, 146, 1063–1074. [Google Scholar] [CrossRef]
- Rovirosa-Hernández, M.; López-Monteon, A.; García-Orduña, F.; Torres-Montero, J.; Guzmán-Gómez, D.; Dumonteil, E.; Waleckx, E.; Lagunes-Merino, O.; Canales-Espinoza, D.; Ramos-Ligonio, A. Natural infection with Trypanosoma cruzi in three species of non-human primates in southeastern Mexico: A contribution to reservoir knowledge. Acta Trop. 2021, 213, 105754. [Google Scholar] [CrossRef]
- Herrera, C.; Truyens, C.; Dumonteil, E.; Alger, J.; Sosa-Estani, S.; Cafferata, M.L.; Gibbons, L.; Ciganda, A.; Matute, M.L.; Zuniga, C.; et al. Phylogenetic analysis of Trypanosoma cruzi from pregnant women and newborns from Argentina, Honduras, and Mexico suggests an association of parasite haplotypes with congenital transmission of the parasite. J. Mol. Diagn. 2019, 21, 1095–1105. [Google Scholar] [CrossRef]
- Rawal, K.; Sinha, R.; Abbasi, B.A.; Chaudhary, A.; Nath, S.K.; Kumari, P.; Preeti, P.; Saraf, D.; Singh, S.; Mishra, K.; et al. Identification of vaccine targets in pathogens and design of a vaccine using computational approaches. Sci. Rep. 2021, 11, 17626. [Google Scholar] [CrossRef]
- Limon-Flores, A.; Cervera-Cetina, R.; Tzec-Arjona, J.; Ek-Macias, L.; Sánchez-Burgos, G.; Ramirez-Sierra, M.; Cruz-Chan, V.; VanWynsberghe, N.R.; Dumonteil, E. Effect of a combination DNA vaccine for the prevention and therapy of Trypanosoma cruzi infection in mice: Role of CD4+ and CD8+ T cells. Vaccine 2010, 28, 7414–7419. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Dumonteil, E.; Heffernan, M.J.; Bottazzi, M.E. Innovation for the `Bottom 100 Million’: Eliminating Neglected Tropical Diseases in the Americas. Adv. Exp. Med. Biol. 2013, 764, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dumonteil, E.; Bottazzi, M.; Zhan, B.; Heffernan, M.; Jones, K.; Valenzuela, J.; Kamhawi, S.; Ortega, J.; Ponce de Leon, S.R.; Lee, B.Y.; et al. Accelerating the development of a therapeutic vaccine for human Chagas disease: Rationale and prospects. Expert. Rev. Vaccines 2012, 11, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnal, A.; Villanueva-Lizama, L.; Teh-Poot, C.; Herrera, C.; Dumonteil, E. Extent of polymorphism and selection pressure on the Trypanosoma cruzi vaccine candidate antigen Tc24. Evol. Appl. 2020, 13, 2663–2672. [Google Scholar] [CrossRef]
- Espinoza, B.; Rico, T.; Sosa, S.; Oaxaca, E.; Vizcaino-Castillo, A.; Caballero, M.; Martínez, I. Mexican Trypanosoma cruzi T. cruzi I strains with different degrees of virulence induce diverse humoral and cellular immune responses in a murine experimental infection model. J. Biomed. Biotechnol. 2010, 2010, 890672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Olmos, V.; Pérez-Nasser, N.; Piñero, D.; Ortega, E.; Hernandez, R.; Espinoza, B. Biological characterization and genetic diversity of Mexican isolates of Trypanosoma cruzi. Acta Trop. 1998, 69, 239–254. [Google Scholar] [CrossRef]
- Da Cruz Moreira, O.; Ramirez, J.C. Genotyping of Trypanosoma cruzi from Clinical Samples by Multilocus Conventional PCR. Methods Mol. Biol. 2019, 1955, 227–238. [Google Scholar] [CrossRef]
- Colunga-Salas, P.; Hernández-Canchola, G. Bats and humans during the SARS-CoV-2 outbreak: The case of bat-coronaviruses from Mexico. Transbound. Emerg. Dis. 2021, 68, 987–992. [Google Scholar] [CrossRef]
- Sánchez-Montes, S.; Salceda-Sánchez, B.; Bermúdez, S.; Aguilar-Tipacamú, G.; Ballados-González, G.; Huerta, H.; Aguilar-Domínguez, M.; Delgado-de la Mora, J.; Licona-Enríquez, J.D.; Delgado-de la Mora, D.; et al. Rhipicephalus sanguineus complex in the Americas: Systematic, genetic diversity, and geographic insights. Pathogens 2021, 10, 1118. [Google Scholar] [CrossRef]
- Nguyen, L.; Schmidt, H.; Von Haeseler, A.; Minh, B. IQ-TREE: A fast and effective stochastic algorithm for estimating Maximum-Likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.; Wong, T.; Von Haeseler, A.; Jermiin, L. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 4, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.; Wright, A.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate Maximum-Likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.; Xie, D.; Baele, G.; Suchard, M. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, N.S.; AbdElbagi, H.; Elsadig, A.R.; Ahmed, A.E.; Mohammed, O.Y.; Elssir, L.T.; Elnour, M.A.; Ali, Y.; Ali, M.S.; Altahir, O.; et al. Assessment of genetic diversity of Plasmodium falciparum circumsporozoite protein in Sudan: The RTS,S leading malaria vaccine candidate. Malar. J. 2021, 20, 436. [Google Scholar] [CrossRef]
- Flores-Alanis, A.; González-Cerón, L.; Santillán-Valenzuela, F.; Ximenez, C.; Sandoval-Bautista, M.A.; Cerritos, R. Spatiotemporal Changes in Plasmodium vivax msp142 Haplotypes in Southern Mexico: From the Control to the Pre-Elimination Phase. Microorganisms 2022, 10, 186. [Google Scholar] [CrossRef]
- Dzul-Huchim, V.M.; Ramirez-Sierra, M.J.; Martinez-Vega, P.P.; Rosado-Vallado, M.E.; Arana-Argaez, V.E.; Ortega-Lopez, J.; Gusovsky, F.; Dumonteil, E.; Cruz-Chan, J.V.; Hotez, P.; et al. Vaccine-linked chemotherapy with a low dose of benznidazole plus a bivalent recombinant protein vaccine prevents the development of cardiac fibrosis caused by Trypanosoma cruzi in chronically-infected BALB/c mice. PLoS Negl. Trop. Dis. 2022, 16, e0010258. [Google Scholar] [CrossRef]
- González-López, C.; Chen, W.H.; Alfaro-Chacón, A.; Villanueva-Lizama, L.E.; Rosado-Vallado, M.; Ramirez-Sierra, M.J.; Teh-Poot, C.F.; Pollet, J.; Asojo, O.; Jones, K.M.; et al. A novel multi-epitope recombinant protein elicits an antigen-specific CD8+ T cells response in Trypanosoma cruzi-infected mice. Vaccine 2022, 40, 6445–6449. [Google Scholar] [CrossRef] [PubMed]
- Quijano-Hernández, I.A.; Castro-Barcena, A.; Vázquez-Chagoyán, J.C.; Bolio-González, M.E.; Ortega-López, J.; Dumonteil, E. Preventive and therapeutic DNA vaccination partially protect dogs against an infectious challenge with Trypanosoma cruzi. Vaccine 2013, 31, 2246–2252. [Google Scholar] [CrossRef]
- Dumonteil, E.; Herrera, C.; Marx, P.A. Safety and preservation of cardiac function following therapeutic vaccination against Trypanosoma cruzi in rhesus macaques. J. Microbiol. Immunol. Infect. 2022, S1684-1182(22)00154–2. [Google Scholar] [CrossRef]
- Viettri, M.; Lares, M.; Medina, M.; Herrera, L.; Ferrer, E. Evaluation of commercial kits for the immunological and molecular diagnosis of Chagas disease in endemic areas of Venezuela. Enferm. Infecc. Microbiol. Clin. 2022, 40, 82–85. [Google Scholar] [CrossRef]
- Chakravarti, I.; Miranda-Schaeubinger, M.; Ruiz-Remigio, A.; Briones-Garduño, C.; Fernández-Figueroa, E.A.; Villanueva-Cabello, C.C.; Borge-Villareal, A.; Bejar-Ramírez, Y.; Pérez-González, A.; Rivera-Benitez, C.; et al. Chagas Disease in Pregnant Women from Endemic Regions Attending the Hospital General de Mexico, Mexico City. Trop. Med. Infect. Dis. 2022, 7, 8. [Google Scholar] [CrossRef]
- Mita-Mendoza, N.K.; McMahon, E.; Kenneson, A.; Barbachano-Guerrero, A.; Beltran-Ayala, E.; Cueva, C.; King, C.A.; Lupone, C.D.; Castro-Sesquen, Y.E.; Gilman, R.H.; et al. Chagas Disease in Southern Coastal Ecuador: Coinfections with Arboviruses and a Comparison of Serological Assays for Chagas Disease Diagnosis. Am. J. Trop. Med. Hyg. 2018, 99, 1530–1533. [Google Scholar] [CrossRef] [Green Version]
- Llano, M.; Pavía, P.; Flórez, A.C.; Cuéllar, A.; González, J.M.; Puerta, C. Evaluación preliminar de la prueba comercial Chagas (Trypanosoma cruzi) IgG-ELISA ® en individuos colombianos [Preliminary evaluation of the commercial kit Chagas (Trypanosoma cruzi) IgG-ELISA ® in Colombian individuals]. Biomedica 2014, 34, 228–236. [Google Scholar] [CrossRef]
- Luquetti, A.O.; Espinoza, B.; Martínez, I.; Hernández-Becerril, N.; Ponce, C.; Ponce, E.; Reyes, P.A.; Hernández, O.; López, R.; Monteón, V. Performance levels of four Latin American laboratories for the serodiagnosis of Chagas disease in Mexican sera samples. Mem. Inst. Oswaldo Cruz 2009, 104, 797–800. [Google Scholar] [CrossRef] [Green Version]
- Vexenat, A.D.C.; Santana, J.M.; Teixeira, A.R. Cross-reactivity of antibodies in human infections by the kinetoplastid protozoa Trypanosoma cruzi, Leishmania chagasi and Leishmania (viannia) braziliensis. Rev. Inst. Med. Trop. Sao Paulo 1996, 38, 177–185. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DTU | Vector | State | Reference |
|---|---|---|---|
| TcI | |||
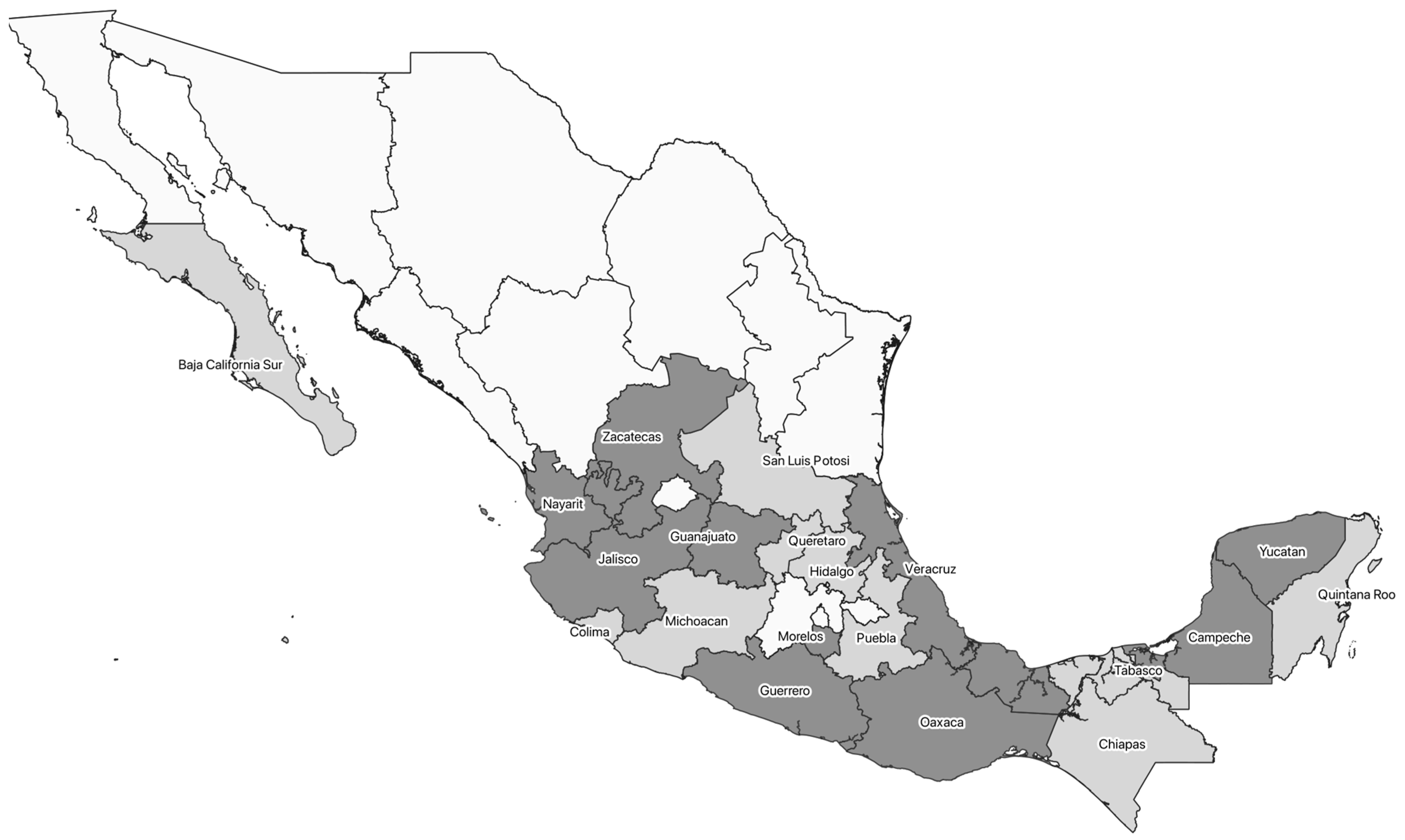
| Dipetalogaster maxima | Baja California Sur | [8,18,19,20] | |
| Panstrongylus rufotuberculatus | Veracruz | [20] | |
| Triatoma barberi | Jalisco, Michoacán, Oaxaca, Puebla, Querétaro | [8,18,21,22,23] | |
| Triatoma dimidiata | Campeche, Chiapas, Michoacán, Quintana Roo, San Luis Potosí, Veracruz, Yucatán | [8,18,20,24,25,26,27,28,29] | |
| Triatoma longipennis | Colima, Jalisco, Michoacán, Nayarit, Zacatecas | [8,21,23,30] | |
| Triatoma pallidipennis | Colima, Jalisco, Michoacán, Morelos, Yucatán, | [8,18,21,23,30] | |
| Triatoma picturata | Jalisco, Nayarit, Sonora | [8,21,26,30] | |
| Triatoma phyllosoma | Nayarit, Zacatecas | [8,21] | |
| Triatoma sp. | Yucatán | [8] | |
| TcII | |||
| T. dimidiata | Veracruz | [20,24,28] | |
| T. pallidipennis | Michoacán | [23] | |
| TcIII | |||
| T. dimidiata | Veracruz | [24] | |
| T. pallidipennis | Michoacán | [23] | |
| TcIV | |||
| T. dimidiata | Quintana Roo, Veracruz | [20,24,29] | |
| T. pallidipennis | Michoacán | [23] | |
| TcV | |||
| T. dimidiata | Veracruz | [24,28] | |
| TcVI | |||
| T. dimidiata | Veracruz | [20,27,28] | |
| Mixed infection | |||
| TcI/TcII | |||
| T. dimidiata | Campeche | [25] | |
| TcI/TcII/TcIV | |||
| T. pallidipennis | Michoacán | [23] |
| DTU | Host/Vector | State | Reference |
|---|---|---|---|
| TcI | |||
| Mammals | |||
| Artiodactyla | Sus scrofa domesticus | Campeche | [25] |
| Carnivora | Canis lupus familiaris | Campeche | [25] |
| Chiroptera | Artibeus jamaicensis | Campeche | [25] |
| Carollia brevicauda | Campeche | [25] | |
| Dermanura phaeotis | Campeche | [25] | |
| Sturnira lilium | Campeche | [25] | |
| Sturnira ludovici | Campeche | [25] | |
| Didelpimorphia | Didelphis marsupialis | Yucatán | [8] |
| Didelphis virginiana | Campeche, Morelos, Veracruz | [8,21,25] | |
| Didelphis sp. | Yucatán | [8,21] | |
| Philander opossum | Veracruz | [8,21] | |
| Rodentia | Peromyscus yucatanicus | Campeche | [25] |
| Peromyscus peromyscus | Puebla | [22] | |
| Heteromys gaumeri | Campeche | [25] | |
| Sigmodon toltecus | Campeche | [25] | |
| Mus musculus | Campeche | [25] | |
| Primates | Alouatta palliata | Tabasco | [31] |
| Alouatta pigra | Tabasco | [31] | |
| Ateles geoffroyi | Veracruz | [31] | |
| Homo sapiens | Guanajuato, Guerrero, Hidalgo, Jalisco, Morelos, Oaxaca, Puebla, San Luis Potosí, Yucatán, Zacatecas | [8,21,22,26,30,32] | |
| TcII | |||
| Artiodactyla | Ovis aries | Campeche | [25] |
| Carnivora | C. lupus familiaris | Campeche | [25] |
| Chiroptera | Myotis keaysi | Campeche | [25] |
| Didelphimorphia | D. virginiana | Veracruz | [21] |
| P. oppossum | Veracruz | [21] | |
| Primates | A. geoffroyi | Veracruz | [31] |
| H. sapiens | Yucatán | [32] | |
| Rodentia | S. toltecus | Campeche | [25] |
| TcV | |||
| Primates | A. pigra | Campeche | [31] |
| TcVI | |||
| Primates | A. geoffroyi | Yucatán | [31] |
| Mixed infection | |||
| TcI/TcII | |||
| Artiodactyla | O. aries | Campeche | [25] |
| S. scrofa domesticus | Campeche | [25] | |
| Carnivora | C. lupus familiaris | Campeche | [25] |
| Chiroptera | Artibeus lituratus | Campeche | [25] |
| Rodentia | S. toltecus | Campeche | [25] |
| Sequence ID | DTU | Host | Country | State | Isolate | GenBank Accession Number |
|---|---|---|---|---|---|---|
| 009-TCR-NAYA-0I-VE | TcI | Triatominae | Mexico | Nayarit | NAYARIT | OL781152 |
| 010-TCR-NI15-0I-HS | TcI | Homo sapiens | Mexico | Oaxaca | NINOA 1.5 | OL781153 |
| 012-TCR-R1H8-0I-HS | TcI | Homo sapiens | Mexico | Yucatán | R1H8 | OL781155 |
| 013-TCR-CALA-0I-VE | TcI | Triatoma dimidiata | Mexico | Campeche | CALAKMUL | OL781156 |
| 014-TCR-ENTO-0I-VE | TcI | Triatominae | Mexico | Morelos | ENTO | OL781157 |
| 015-TCR-INC1-0I-HS | TcI | Homo sapiens | Mexico | Oaxaca | INC-1 | OL781158 |
| 016-TCR-INC5-0I-HS | TcI | Homo sapiens | Mexico | Guanajuato | INC-5 | OL781159 |
| 017-TCR-INC6-0I-HS | TcI | Homo sapiens | Mexico | Oaxaca | INC-6 | OL781160 |
| 018-TCR-INC9-0I-HS | TcI | Homo sapiens | Mexico | Guerrero | INC-9 | OL781161 |
| 019-TCR-IN10-01-HS | TcI | Homo sapiens | Mexico | Guanajuato | INC-10 | OL781162 |
| 020-TCR-CAM7-0I-VE | TcI | Triatoma dimidiata | Mexico | Campeche | CAM-7 | OL781163 |
| 021-TCR-CAM8-0I-VE | TcI | Triatoma dimidiata | Mexico | Campeche | CAM-8 | OL781164 |
| 022-TCR-HUEY-0I-VE | TcI | Triatoma barberi | Mexico | Estado de México | HUEYPOXTLA | OL781165 |
| 023-TCR-NINO-0I-HS | TcI | Homo sapiens | Mexico | Oaxaca | NINOA | OL781166 |
| 024-TCR-SONT-0I-TD | TcI | Triatoma dimidiata | Mexico | Veracruz | SONTECOMAPAN | OL781167 |
| 025-TCR-TEZO-0I-VE | TcI | Triatoma barberi | Mexico | Estado de México | TEZONTLALPAN | OL781168 |
| 6Morelos | TcI | Homo sapiens | Mexico | Morelos | MORELOS | OL781175 |
| 7CoculaJALISCO | TcI | Triatominae | Mexico | Jalisco | COCULA | OL781176 |
| 9JerboYUCATAN | TcI | Homo sapiens | Mexico | Yucatán | JERBO | OL781177 |
| 13Zacatecas | TcI | Homo sapiens | Mexico | Zacatecas | ZACATECAS | OL781178 |
| 14PeNaGUERRERO | TcI | Homo sapiens | Mexico | Guerrero | PEÑA | OL781179 |
| 15PetaquillasGUERRER | TcI | Triatominae | Mexico | Guerrero | PETAQUILLAS | OL781180 |
| 23ProgresoMOLRELOS | TcI | Triatominae | Mexico | Morelos | PROGRESO | OL781181 |
| 25Yuc | TcI | Triatominae | Mexico | Yucatán | YUC | OL781182 |
| ENTOMO | TcI | ND | ND | ND | ENTOMO | OL781183 |
| 011-TCR-DM22-0I-DM | TcI | Didelphis marsupialis | Brazil | ND | DM28 | OL781154 |
| 035-DTUI | TcI | Didelphis marsupialis | Venezuela | ND | DM28 | OL781169 |
| 036-DTUII | TcII | Homo sapiens | Brazil | ND | 000Y | OL781170 |
| 037-DTUIII | TcIII | Panstrongylus geniculatus | Brazil | Manaus | 3663 | OL781171 |
| 038-DTUIV | TcIV | Rhodnius brethesi | Brazil | Barcelos | 4167 | OL781172 |
| 039-DTUV | TcV | Triatoma infestans | Argentina | Chaco | LL014 | OL781173 |
| 040-DTUVI | TcVI | Triatoma infestans | Brazil | Rio Grande do Sul | CL Brener | OL781174 |
| 002-TCR-MAST-NR-CL | ND | Canis lupus familiaris | US | Texas | MASTIFF | DQ183066 |
| 004-TCR-CLBR-VI-HS | TcVI | Triatoma infestans | Brazil | ND | CLBRENER | XM_800482.1 |
| 005-TCR-000Y-II-HS | TcII | Homo sapiens | Brazil | ND | 000Y | D87512.1 |
| 006-TCR-00Y2-II-HS | TcII | Homo sapiens | Brazil | ND | 000Y | S43664.1 |
| 007-TCR-CLB2-VI-HS | TcVI | Triatoma infestans | Brazil | ND | CL BRENER | AF192980.2 |
| 001-TCO-025E-NR-RR | Trypanosoma conorhini | Rattus rattus | Brazil | NR | 025E | XM_029375354.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, I.; Miranda-Ortiz, H.; Fernández-Figueroa, E.A.; Sánchez-Montes, S.; Colunga-Salas, P.; Grostieta, E.; Juárez-Gabriel, J.; Lozano-Sardaneta, Y.N.; Arce-Fonseca, M.; Rodríguez-Morales, O.; et al. The Low Variability of Tc24 in Trypanosoma cruzi TcI as an Advantage for Chagas Disease Prophylaxis and Diagnosis in Mexico. Pathogens 2023, 12, 368. https://doi.org/10.3390/pathogens12030368
Becker I, Miranda-Ortiz H, Fernández-Figueroa EA, Sánchez-Montes S, Colunga-Salas P, Grostieta E, Juárez-Gabriel J, Lozano-Sardaneta YN, Arce-Fonseca M, Rodríguez-Morales O, et al. The Low Variability of Tc24 in Trypanosoma cruzi TcI as an Advantage for Chagas Disease Prophylaxis and Diagnosis in Mexico. Pathogens. 2023; 12(3):368. https://doi.org/10.3390/pathogens12030368
Chicago/Turabian StyleBecker, Ingeborg, Haydee Miranda-Ortiz, Edith A. Fernández-Figueroa, Sokani Sánchez-Montes, Pablo Colunga-Salas, Estefanía Grostieta, Javier Juárez-Gabriel, Yokomi N. Lozano-Sardaneta, Minerva Arce-Fonseca, Olivia Rodríguez-Morales, and et al. 2023. "The Low Variability of Tc24 in Trypanosoma cruzi TcI as an Advantage for Chagas Disease Prophylaxis and Diagnosis in Mexico" Pathogens 12, no. 3: 368. https://doi.org/10.3390/pathogens12030368