
Uncovering a Complex Virome Associated with the Cacao Pathogens Ceratocystis cacaofunesta and Ceratocystis fimbriata

 , , , ,
, , , ,  , , , and
, , , and 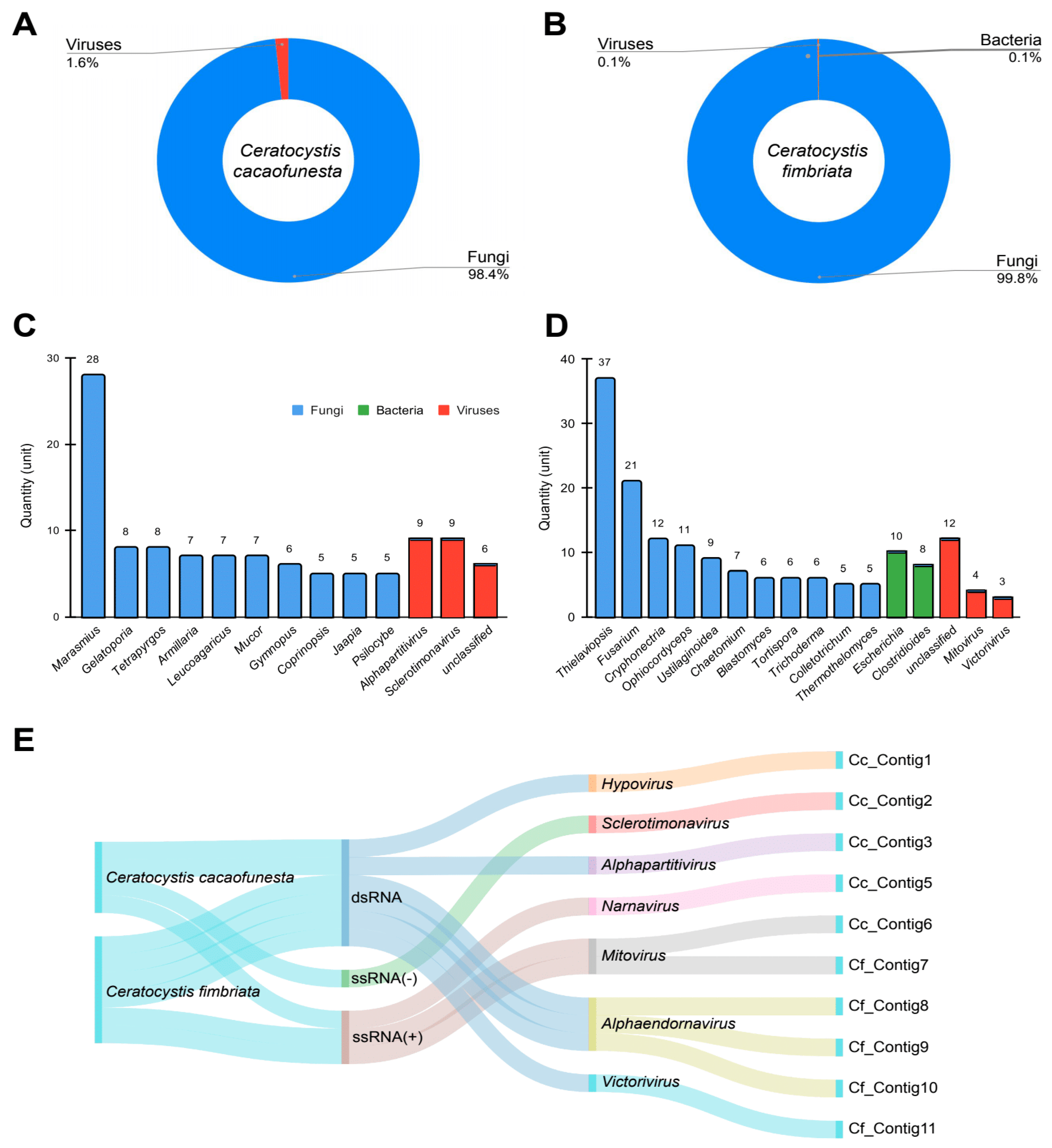{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Acquisition of RNA Libraries
2.2. Bioinformatic Analysis
2.2.1. Transcriptome Assembly
2.2.2. Identification of Viral Sequences
2.2.3. Manual Curation of Viral Genomes
2.2.4. Integrative Genome Assembly
2.3. Phylogenetic Analysis
2.4. Genetic Diversity of Species and Abundance of Viral Sequences
2.5. Characterization of RNAi-Related Proteins in C. fimbriata
3. Results
3.1. Transcriptome Assembly
3.2. Metagenomic Analysis
3.3. Virome Characterization
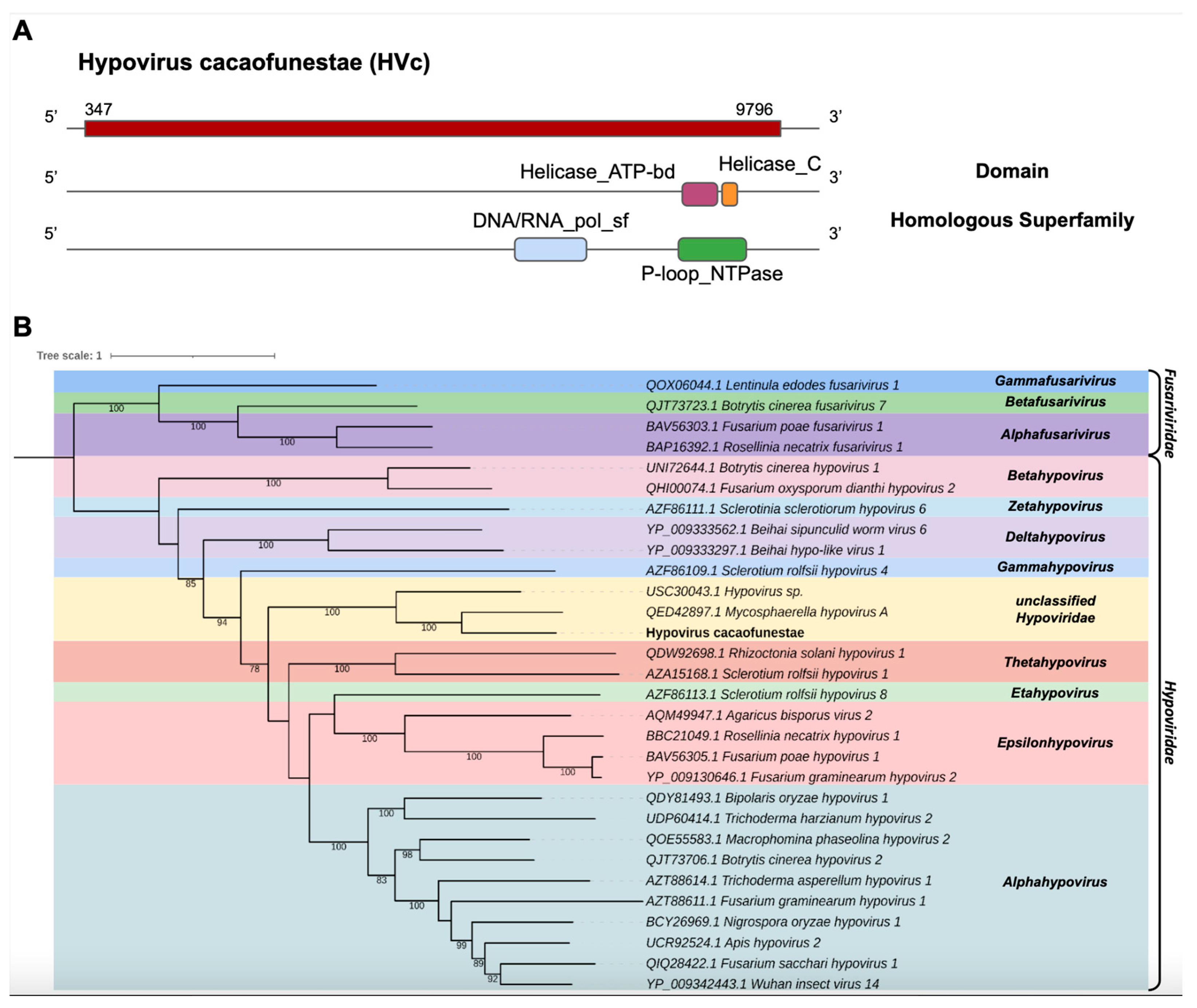
- Hypoviridae
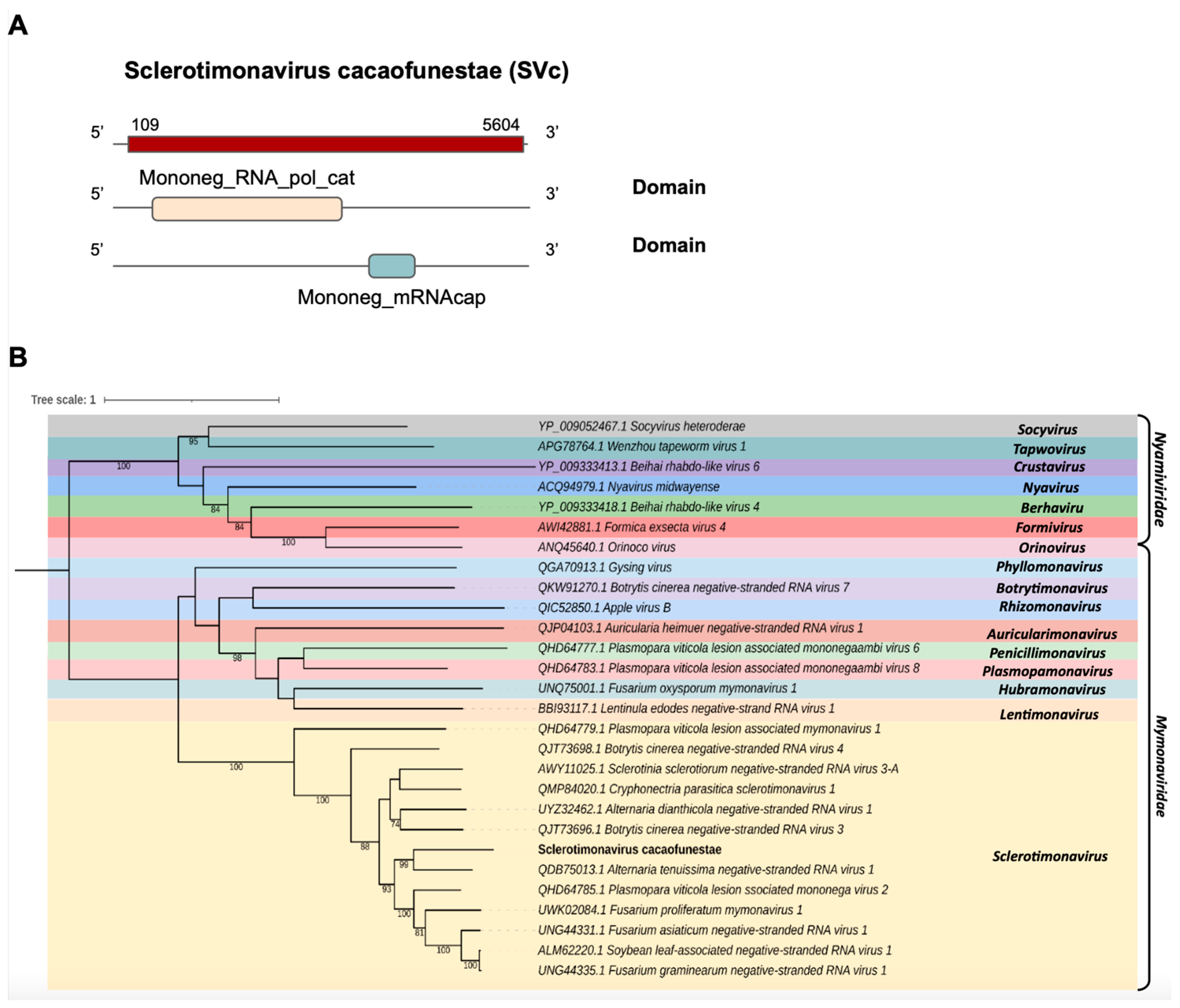
- Mymonaviridae
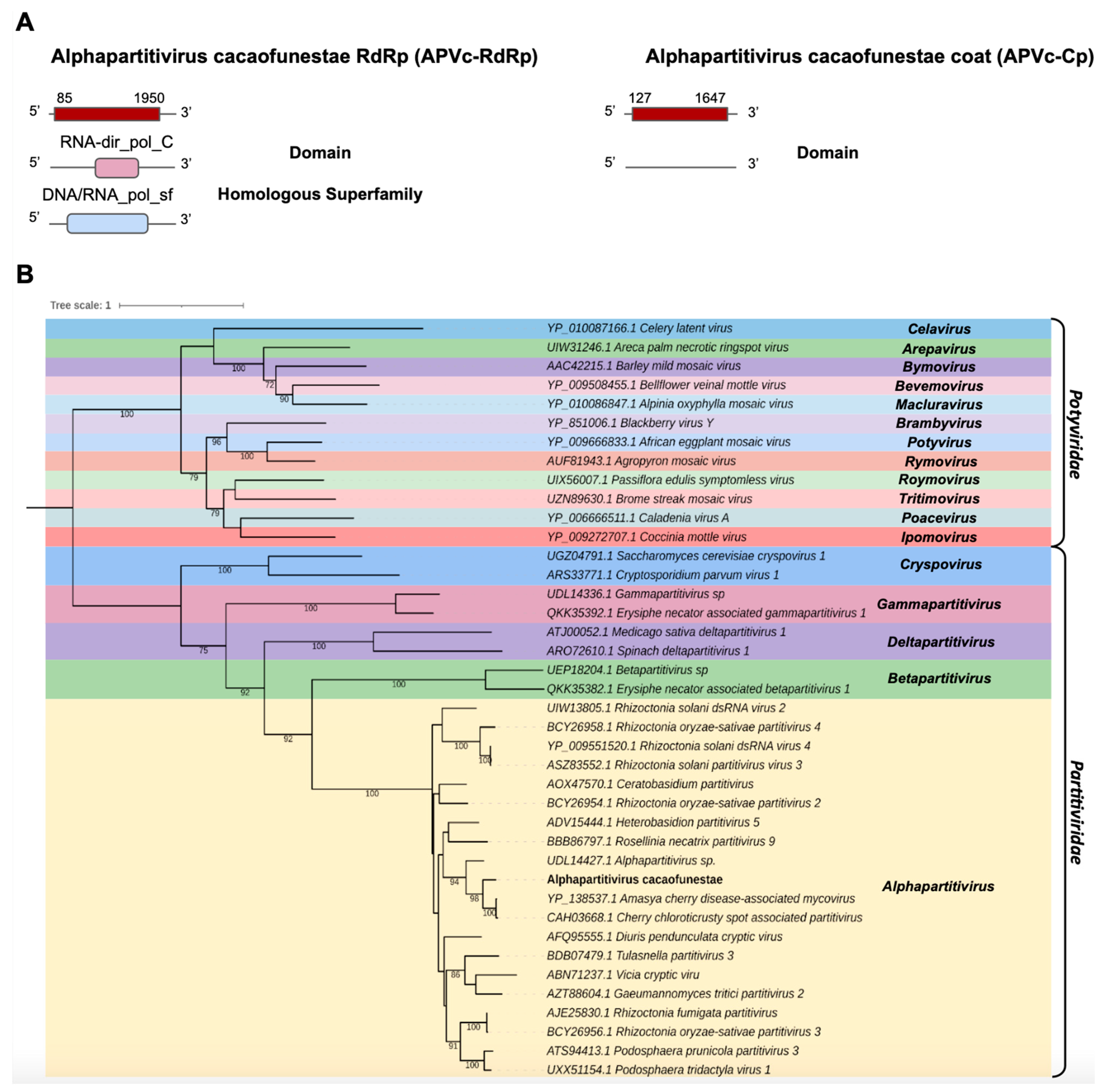
- Partitiviridae
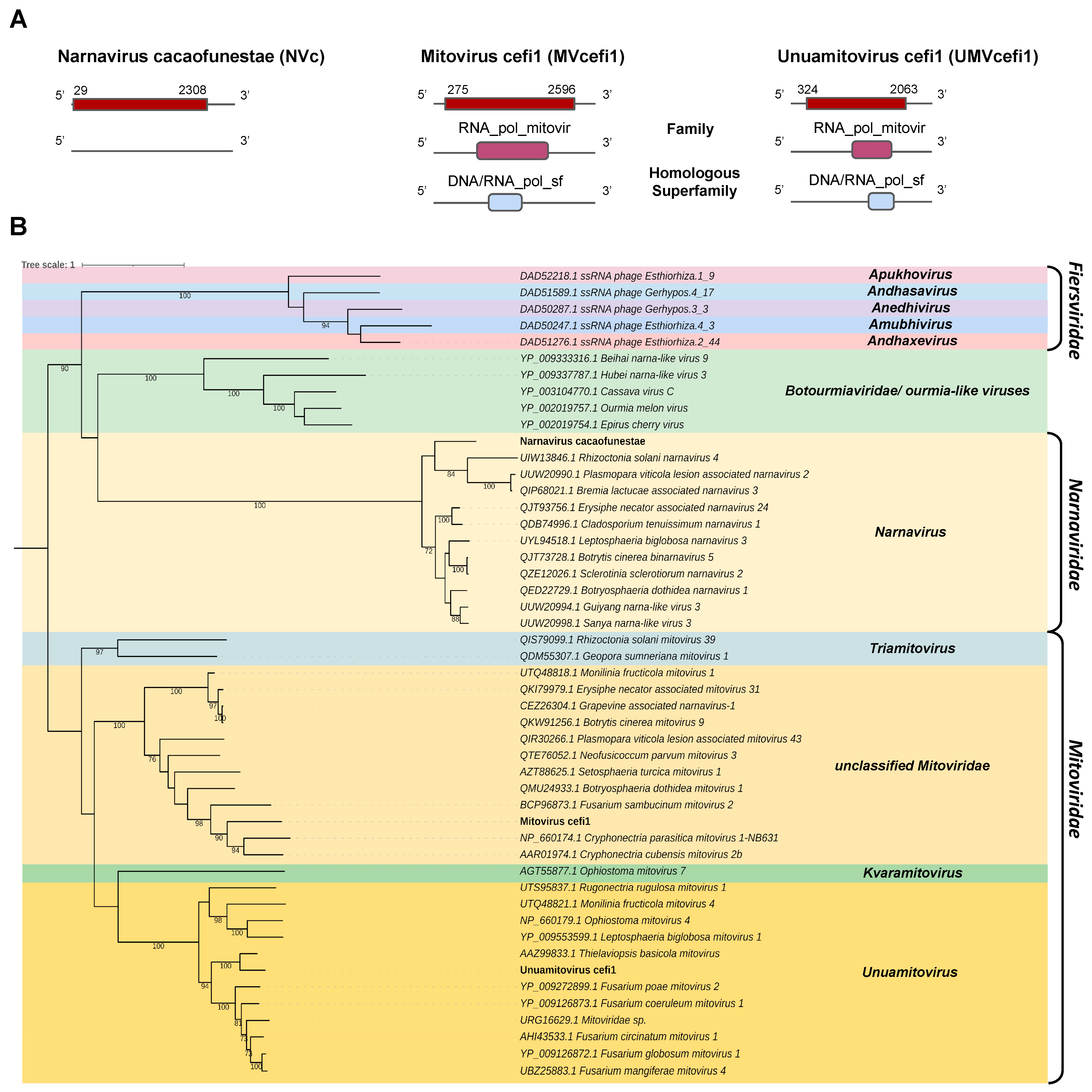
- Narnaviridae and Mitoviridae
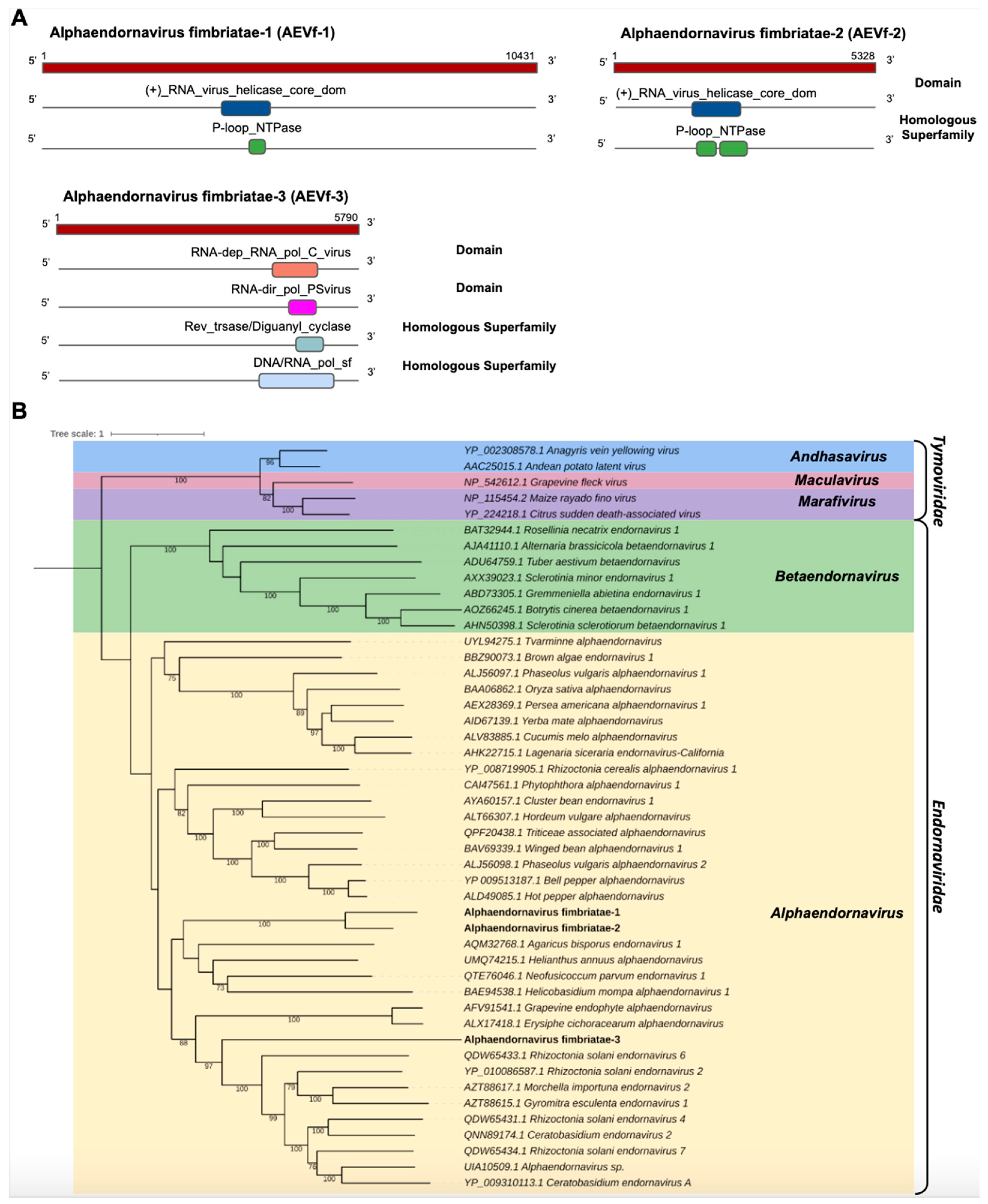
- Endornaviridae
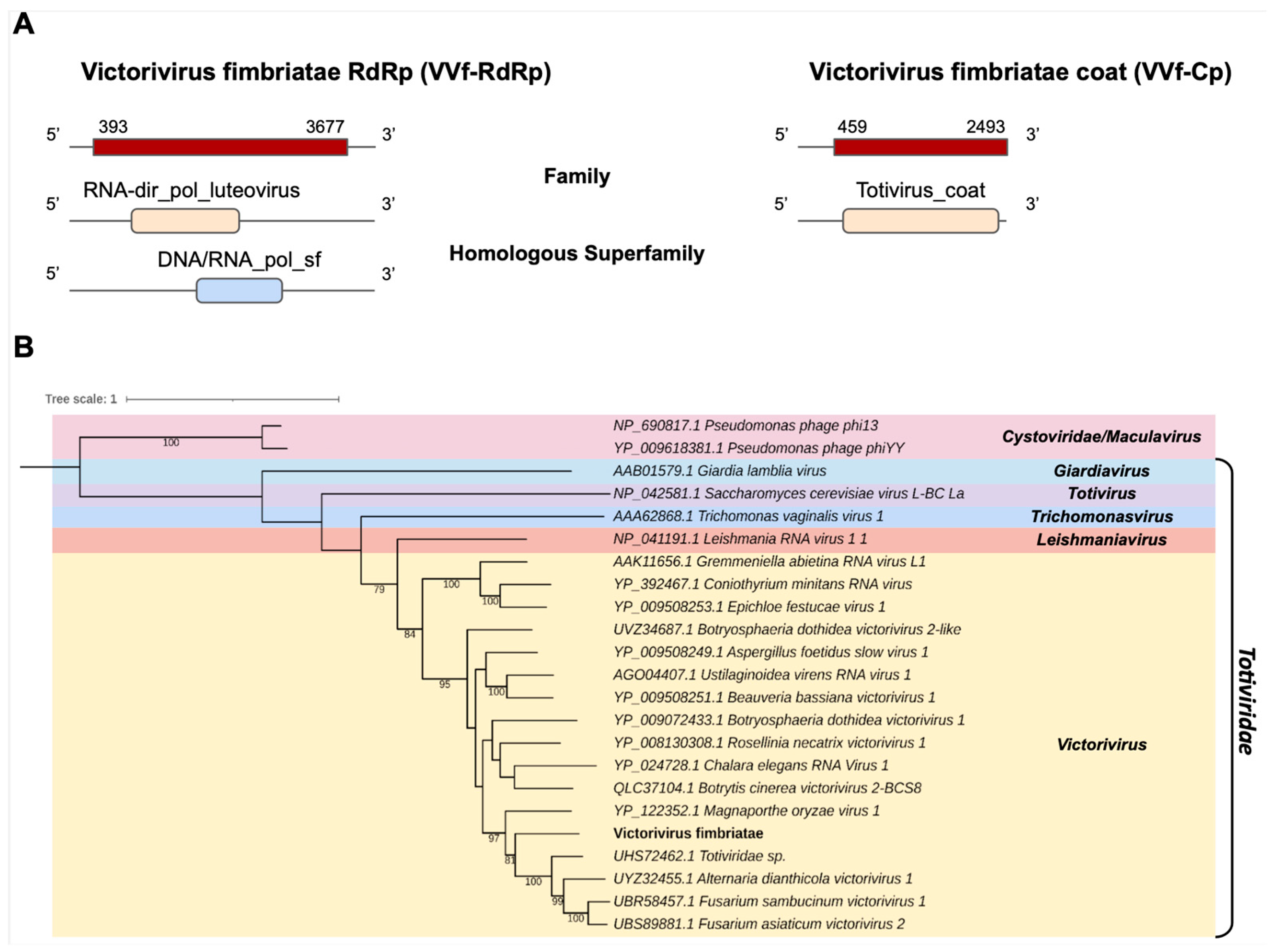
- Totiviridae
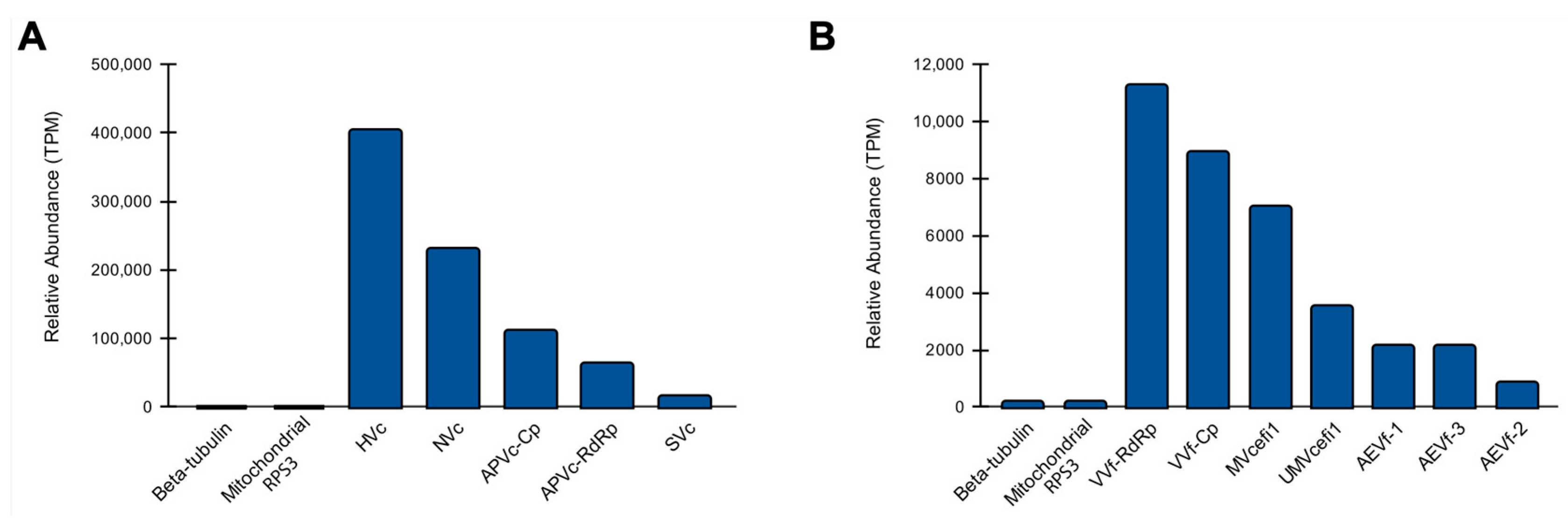
3.4. Abundance of Viral Sequences
3.5. Characterization of RNA-Interference-Related Genes in Ceratocystis Fimbriata
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bekele, F.; Phillips-Mora, W. Cacao (Theobroma Cacao L.) Breeding. In Advances in Plant Breeding Strategies: Industrial and Food Crops: Volume 6; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 409–487. [Google Scholar] [CrossRef]
- Fernandes, L.D.S.; Royaert, S.; Corrêa, F.M.; Mustiga, G.M.; Marelli, J.-P.; Corrêa, R.X.; Motamayor, J.C. Mapping of a Major QTL for Ceratocystis Wilt Disease in an F1 Population of Theobroma Cacao. Front. Plant Sci. 2018, 9, 155. [Google Scholar] [CrossRef] [PubMed]
- Marelli, J.-P.; Guest, D.I.; Bailey, B.A.; Evans, H.C.; Brown, J.K.; Junaid, M.; Barreto, R.W.; Lisboa, D.O.; Puig, A.S. Chocolate Under Threat from Old and New Cacao Diseases. Phytopathology 2019, 109, 1331–1343. [Google Scholar] [CrossRef] [PubMed]
- International Cocoa Organization—Statistics 2021. Available online: https://www.icco.org/statistics/ (accessed on 17 October 2021).
- Hussain, B. Modernization in Plant Breeding Approaches for Improving Biotic Stress Resistance in Crop Plants. Turk. J. Agric. For. 2015, 39, 515–530. [Google Scholar] [CrossRef]
- Ploetz, R. The Impact of Diseases on Cacao Production: A Global Overview. In Cacao Diseases: A History of Old Enemies and New Encounters; Bailey, B.A., Meinhardt, L.W., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 33–59. [Google Scholar] [CrossRef]
- Cabrera, O.G.; Molano, E.P.L.; José, J.; Álvarez, J.C.; Pereira, G.A.G. Ceratocystis Wilt Pathogens: History and Biology—Highlighting C. Cacaofunesta, the Causal Agent of Wilt Disease of Cacao. In Cacao Diseases: A History of Old Enemies and New Encounters; Bailey, B.A., Meinhardt, L.W., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 383–428. [Google Scholar] [CrossRef]
- Delgado-Ospina, J.; Molina-Hernández, J.B.; Chaves-López, C.; Romanazzi, G.; Paparella, A. The Role of Fungi in the Cocoa Production Chain and the Challenge of Climate Change. J. Fungi 2021, 7, 202. [Google Scholar] [CrossRef]
- Santos, S.A.; Vidigal, P.M.P.; Thrimawithana, A.; Betancourth, B.M.L.; Guimarães, L.M.S.; Templeton, M.D.; Alfenas, A.C. Comparative Genomic and Transcriptomic Analyses Reveal Different Pathogenicity-Related Genes among Three Eucalyptus Fungal Pathogens. Fungal Genet. Biol. 2020, 137, 103332. [Google Scholar] [CrossRef]
- Molano, E.P.L.; Cabrera, O.G.; Jose, J.; do Nascimento, L.C.; Carazzolle, M.F.; Teixeira, P.J.P.L.; Alvarez, J.C.; Tiburcio, R.A.; Tokimatu Filho, P.M.; de Lima, G.M.A.; et al. Ceratocystis Cacaofunesta Genome Analysis Reveals a Large Expansion of Extracellular Phosphatidylinositol-Specific Phospholipase-C Genes (PI-PLC). BMC Genom. 2018, 19, 58. [Google Scholar] [CrossRef]
- Engelbrecht, C.J.B.; Harrington, T.C. Intersterility, Morphology and Taxonomy of Ceratocystis Fimbriata on Sweet Potato, Cacao and Sycamore. Mycologia 2005, 97, 57–69. [Google Scholar] [CrossRef]
- Raffa, K.F.; Hobson, K.R.; Lafontaine, S.; Aukema, B.H. Can Chemical Communication Be Cryptic? Adaptations by Herbivores to Natural Enemies Exploiting Prey Semiochemistry. Oecologia 2007, 153, 1009–1019. [Google Scholar] [CrossRef]
- Smith, S.M.; Hulcr, J. Chapter 12—Scolytus and Other Economically Important Bark and Ambrosia Beetles. In Bark Beetles; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 495–531. [Google Scholar] [CrossRef]
- da Cruz, M.B.; de Sousa, D.F.; de Almeida Oliveira, L.; de França, J.P.; de França, L.P.; Conceição, A.O.; Fonseca Chaves, A.L. Interference of Aqueous and Ethanolic Solutions of Adiantum Latifolium Lam. (Pteridaceae) Leaves on in Vitro Ceratocystis Cacaofunesta Mycelial Growth. Arq. Inst. Biol. 2019, 86. [Google Scholar] [CrossRef]
- Mora-Ocampo, I.Y.; Pirovani, C.P.; Luz, E.D.M.N.; Rêgo, A.P.B.; Silva, E.M.A.; Rhodes-Valbuena, M.; Corrêa, R.X. Ceratocystis Cacaofunesta Differentially Modulates the Proteome in Xylem-Enriched Tissue of Cocoa Genotypes with Contrasting Resistance to Ceratocystis Wilt. Planta 2021, 254, 94. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Xu, M.; Guo, J.; Zhang, C.; Feng, Z.; Peng, X.; Li, Z.; Xing, K.; Qin, S. Antifungal Effect of Volatile Organic Compounds Produced by Pseudomonas Chlororaphis Subsp. Aureofaciens SPS-41 on Oxidative Stress and Mitochondrial Dysfunction of Ceratocystis Fimbriata. Pestic. Biochem. Physiol. 2021, 173, 104777. [Google Scholar] [CrossRef] [PubMed]
- Aime, M.C.; Phillips-Mora, W. The Causal Agents of Witches’ Broom and Frosty Pod Rot of Cacao (Chocolate, Theobroma Cacao) Form a New Lineage of Marasmiaceae. Mycol. 2005, 97, 1012–1022. [Google Scholar] [CrossRef]
- Surujdeo-Maharaj, S.; Sreenivasan, T.N.; Motilal, L.A.; Umaharan, P. Black Pod and Other Phytophthora Induced Diseases of Cacao: History, Biology, and Control. In Cacao Diseases: A History of Old Enemies and New Encounters; Bailey, B.A., Meinhardt, L.W., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 213–266. [Google Scholar] [CrossRef]
- Rodrigues, G.d.S.; Magalhães, D.M.A.; da Costa, A.M.; Luz, E.D.M.N. Antagonism of Trichoderma Spp. To the Etiological Agent of Ceratocystis Wilt in Cacao. Summa Phytopathol. 2018, 44, 72–78. [Google Scholar] [CrossRef]
- Torres-Trenas, A.; Prieto, P.; Cañizares, M.C.; García-Pedrajas, M.D.; Pérez-Artés, E. Mycovirus Fusarium Oxysporum f. Sp. Dianthi Virus 1 Decreases the Colonizing Efficiency of Its Fungal Host. Front Cell Infect. Microbiol. 2019, 9, 51. [Google Scholar] [CrossRef]
- Sharma, M.; Guleria, S.; Singh, K.; Chauhan, A.; Kulshrestha, S. Mycovirus Associated Hypovirulence, a Potential Method for Biological Control of Fusarium Species. Virus Dis. 2018, 29, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Wagemans, J.; Holtappels, D.; Vainio, E.; Rabiey, M.; Marzachì, C.; Herrero, S.; Ravanbakhsh, M.; Tebbe, C.C.; Ogliastro, M.; Ayllón, M.A.; et al. Going Viral: Virus-Based Biological Control Agents for Plant Protection. Annu. Rev. Phytopathol. 2022, 60, 21–42. [Google Scholar] [CrossRef]
- Wen, C.; Wan, X.; Zhang, Y.; Du, H.; Wei, C.; Zhong, R.; Zhang, H.; Shi, Y.; Xie, J.; Fu, Y.; et al. Molecular Characterization of the First Alternavirus Identified in Fusarium Oxysporum. Viruses 2021, 13, 2026. [Google Scholar] [CrossRef]
- Xiao, X.; Cheng, J.; Tang, J.; Fu, Y.; Jiang, D.; Baker, T.S.; Ghabrial, S.A.; Xie, J. A Novel Partitivirus That Confers Hypovirulence on Plant Pathogenic Fungi. J. Virol. 2014, 88, 10120–10133. [Google Scholar] [CrossRef]
- Zhu, J.Z.; Guo, J.; Hu, Z.; Zhang, X.T.; Li, X.G.; Zhong, J. A Novel Partitivirus That Confer Hypovirulence to the Plant Pathogenic Fungus Colletotrichum Liriopes. Front Microbiol 2021, 12, 653809. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.Z.; Zhu, H.J.; Gao, B.D.; Zhou, Q.; Zhong, J. Diverse, Novel Mycoviruses From the Virome of a Hypovirulent Sclerotium Rolfsii Strain. Front Plant Sci. 2018, 9, 1738. [Google Scholar] [CrossRef]
- Segers, G.C.; Zhang, X.; Deng, F.; Sun, Q.; Nuss, D.L. Evidence That RNA Silencing Functions as an Antiviral Defense Mechanism in Fungi. Proc. Natl. Acad. Sci. USA 2007, 104, 12902–12906. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lee, K.-M.; Cho, W.K.; Park, J.Y.; Kim, K.-H. Differential Contribution of RNA Interference Components in Response to Distinct Fusarium Graminearum Virus Infections. J. Virol. 2018, 92, e01756-e17. [Google Scholar] [CrossRef] [PubMed]
- Nuss, D.L. Mycoviruses, RNA Silencing, and Viral RNA Recombination. Adv. Virus Res. 2011, 80, 25–48. [Google Scholar] [CrossRef] [PubMed]
- Nicolás, F.E.; Ruiz-Vázquez, R.M. Functional Diversity of RNAi-Associated SRNAs in Fungi. Int. J. Mol. Sci. 2013, 14, 15348–15360. [Google Scholar] [CrossRef]
- Nicolás, F.E.; Vila, A.; Moxon, S.; Cascales, M.D.; Torres-Martínez, S.; Ruiz-Vázquez, R.M.; Garre, V. The RNAi Machinery Controls Distinct Responses to Environmental Signals in the Basal Fungus Mucor Circinelloides. BMC Genom. 2015, 16, 237. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.; Yang, Q.; Xue, Z.; Liu, Y. RNA Interference in Fungi: Pathways, Functions, and Applications. Eukaryot. Cell 2011, 10, 1148–1155. [Google Scholar] [CrossRef]
- Liu, Y. RNA Interference Pathways in Filamentous Fungi. Cell Mol. Life Sci. 2010, 67, 3849–3863. [Google Scholar] [CrossRef]
- Khatri, M.; Rajam, M.V. Targeting Polyamines of Aspergillus Nidulans by SiRNA Specific to Fungal Ornithine Decarboxylase Gene. Med. Mycol. 2007, 45, 211–220. [Google Scholar] [CrossRef]
- Mukherjee, K.; Campos, H.; Kolaczkowski, B. Evolution of Animal and Plant Dicers: Early Parallel Duplications and Recurrent Adaptation of Antiviral RNA Binding in Plants. Mol. Biol. Evol. 2013, 30, 627–641. [Google Scholar] [CrossRef]
- Nakayashiki, H.; Kadotani, N.; Mayama, S. Evolution and Diversification of RNA Silencing Proteins in Fungi. J. Mol. Evol. 2006, 63, 127–135. [Google Scholar] [CrossRef]
- Roossinck, M.J. Metagenomics of Plant and Fungal Viruses Reveals an Abundance of Persistent Lifestyles. Front Microbiol. 2015, 5, 767. [Google Scholar] [CrossRef] [PubMed]
- Mokili, J.L.; Rohwer, F.; Dutilh, B.E. Metagenomics and Future Perspectives in Virus Discovery. Curr. Opin. Virol. 2012, 2, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.C.; Wilken, P.M.; Coetzee, M.P.A.; Wingfield, M.J.; Wingfield, B.D. Analysis of Microsatellite Markers in the Genome of the Plant Pathogen Ceratocystis Fimbriata. Fungal. Biol. 2013, 117, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Wilken, P.M.; Steenkamp, E.T.; Wingfield, M.J.; de Beer, Z.W.; Wingfield, B.D. IMA Genome-F 1: Ceratocystis Fimbriata: Draft Nuclear Genome Sequence for the Plant Pathogen, Ceratocystis Fimbriata. IMA Fungus 2013, 4, 357–358. [Google Scholar] [CrossRef]
- Wingett, S.W.; Andrews, S. FastQ Screen: A Tool for Multi-Genome Mapping and Quality Control. F1000Res 2018, 7, 1338. [Google Scholar] [CrossRef] [PubMed]
- BI Ageing Clock Team; Stubbs, T.M.; Bonder, M.J.; Stark, A.-K.; Krueger, F.; von Meyenn, F.; Stegle, O.; Reik, W. Multi-Tissue DNA Methylation Age Predictor in Mouse. Genome Biol. 2017, 18, 68. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Prjibelski, A.D.; Vasilinetc, I.; Bankevich, A.; Gurevich, A.; Krivosheeva, T.; Nurk, S.; Pham, S.; Korobeynikov, A.; Lapidus, A.; Pevzner, P.A. ExSPAnder: A Universal Repeat Resolver for DNA Fragment Assembly. Bioinformatics 2014, 30, i293–i301. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Huang, X.; Madan, A. CAP3: A DNA Sequence Assembly Program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment Using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Kyran, A. Fasta Statistics: Display Summary Statistics for a Fasta File 2021. Available online: https://github.com/galaxyproject/tools-iuc (accessed on 5 November 2022).
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2018 Update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Rombel, I.T.; Sykes, K.F.; Rayner, S.; Johnston, S.A. ORF-FINDER: A Vector for High-Throughput Gene Identification. Gene 2002, 282, 33–41. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro Protein Families and Domains Database: 20 Years On. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER Web Server: Interactive Sequence Similarity Searching. Nucleic Acids Res. 2011, 39 (Suppl. S2), W29–W37. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Arndt, W.; Miller, B.L.; Wheeler, T.J.; Schreiber, F.; Bateman, A.; Eddy, S.R. HMMER Web Server: 2015 Update. Nucleic Acids Res. 2015, 43, W30–W38. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER Web Server: 2018 Update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Almeida, A.; Beracochea, M.; Boland, M.; Burgin, J.; Cochrane, G.; Crusoe, M.R.; Kale, V.; Potter, S.C.; Richardson, L.J.; et al. MGnify: The Microbiome Analysis Resource in 2020. Nucleic Acids Res. 2020, 48, D570–D578. [Google Scholar] [CrossRef]
- Schulz, M.H.; Zerbino, D.R.; Vingron, M.; Birney, E. Oases: Robust de Novo RNA-Seq Assembly across the Dynamic Range of Expression Levels. Bioinformatics 2012, 28, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Afiahayati; Sato, K.; Sakakibara, Y. MetaVelvet-SL: An Extension of the Velvet Assembler to a de Novo Metagenomic Assembler Utilizing Supervised Learning. DNA Res. 2015, 22, 69–77. [Google Scholar] [CrossRef]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de Novo Assembler for Single-Cell and Metagenomic Sequencing Data with Highly Uneven Depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. Cd-Hit: A Fast Program for Clustering and Comparing Large Sets of Protein or Nucleotide Sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. AliView: A Fast and Lightweight Alignment Viewer and Editor for Large Datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The Conserved Domain Database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and Sensitive Taxonomic Classification for Metagenomics with Kaiju. Nat. Commun 2016, 7, 11257. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon Provides Fast and Bias-Aware Quantification of Transcript Expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003, Unit 2.3. [Google Scholar] [CrossRef]
- Han, M.V.; Zmasek, C.M. PhyloXML: XML for Evolutionary Biology and Comparative Genomics. BMC Bioinform. 2009, 10, 356. [Google Scholar] [CrossRef] [PubMed]
- Marincowitz, S.; Barnes, I.; de Beer, Z.W.; Wingfield, M.J. Epitypification of Ceratocystis Fimbriata. Fungal Syst. Evol. 2020, 6, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, M.; Wang, Y.; Li, L.; Wang, M.; Li, X.; Xu, M.; Loake, G.J.; Guo, M.; Jiang, J. Ceratocystis Fimbriata Employs a Unique Infection Strategy Targeting Peltate Glandular Trichomes of Sweetpotato (Ipomoea Batatas) Plants. Phytopathology 2020, 110, 1923–1933. [Google Scholar] [CrossRef]
- Stahr, M.; Quesada-Ocampo, L.M. Assessing the Role of Temperature, Inoculum Density, and Wounding on Disease Progression of the Fungal Pathogen Ceratocystis Fimbriata Causing Black Rot in Sweetpotato. Plant Dis. 2020, 104, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Harrington, T.C.; Thorpe, D.J.; Alfenas, A.C. Genetic Variation and Variation in Aggressiveness to Native and Exotic Hosts among Brazilian Populations of Ceratocystis Fimbriata. Phytopathology 2011, 101, 555–566. [Google Scholar] [CrossRef]
- Kumari, N.; Shukla, P.K.; Singh, H.; Fatima, T.; Bajpai, A. Development of Species-Specific PCR Based Detection Assay for Ceratocystis Fimbriata, Mango Wilt Pathogen. Indian Phytopathol. 2022, 75, 215–220. [Google Scholar] [CrossRef]
- Fourie, A.; Wingfield, M.J.; Wingfield, B.D.; Barnes, I. Molecular Markers Delimit Cryptic Species in Ceratocystis Sensu Stricto. Mycol. Prog. 2014, 14, 1020. [Google Scholar] [CrossRef]
- Harrington, T.C.; Kazmi, M.R.; Al-Sadi, A.M.; Ismail, S.I. Intraspecific and Intragenomic Variability of ITS RDNA Sequences Reveals Taxonomic Problems in Ceratocystis Fimbriata Sensu Stricto. Mycologia 2014, 106, 224–242. [Google Scholar] [CrossRef] [PubMed]
- Yakushiji, H.; Morita, T.; Jikumaru, S.; Ikegami, H.; Azuma, A.; Koshita, Y. Interspecific Hybridization of Fig (Ficus Carica L.) and Ficus Erecta Thunb., a Source of Ceratocystis Canker Resistance. Euphytica 2012, 183, 39–47. [Google Scholar] [CrossRef]
- Li, Q.; Harrington, T.C.; McNew, D.; Li, J.; Huang, Q.; Somasekhara, Y.M.; Alfenas, A.C. Genetic Bottlenecks for Two Populations of Ceratocystis Fimbriata on Sweet Potato and Pomegranate in China. Plant Dis. 2016, 100, 2266–2274. [Google Scholar] [CrossRef]
- Huang, Q.; Zhu, Y.Y.; Chen, H.R.; Wang, Y.Y.; Liu, Y.L.; Lu, W.J.; Ruan, X.Y. First Report of Pomegranate Wilt Caused by Ceratocystis Fimbriata in Yunnan, China. Plant Dis. 2003, 87, 1150. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, Y.Y.; Zhao, Y.Y.; Jiao, Y.X.; Li, X.F.; Chen, H.R.; Zhu, Y.Y. First Report of Taro Black Rot Caused by Ceratocystis Fimbriata in China. Plant Pathol. 2008, 57, 780. [Google Scholar] [CrossRef]
- Harrington, T.C.; Thorpe, D.J.; Marinho, V.L.A.; Furtado, E.L. First Report of Black Rot of Colocasia Esculenta Caused by Ceratocystis Fimbriata in Brazil. Fitopatol. Bras. 2005, 30, 88–89. [Google Scholar] [CrossRef]
- Baker, C.J.; Harrington, T.C.; Krauss, U.; Alfenas, A.C. Genetic Variability and Host Specialization in the Latin American Clade of Ceratocystis Fimbriata. Phytopathology 2003, 93, 1274–1284. [Google Scholar] [CrossRef]
- Méndez-Álvarez, D.; de Souza Cândido, T.; Couto Alfenas, A.; Murillo, O.; Badilla, Y.; Ferreira Alfenas, R. First Report of Ceratocystis Fimbriata Causing Wilt on Gmelina Arborea in Costa Rica. For. Pathol. 2020, 50, e12628. [Google Scholar] [CrossRef]
- Thorpe, D.J.; Harrington, T.C.; Uchida, J.Y. Pathogenicity, Internal Transcribed Spacer-RDNA Variation, and Human Dispersal of Ceratocystis Fimbriata on the Family Araceae. Phytopathology 2005, 95, 316–323. [Google Scholar] [CrossRef]
- Walker, J.; Tesoriero, L.; Pascoe, I.; Forsberg, L.I. Basal Rot of Syngonium Cultivars and the First Record of Ceratocystis Fimbriata from Australia. Australas. Plant Pathol. 1988, 17, 22–23. [Google Scholar] [CrossRef]
- De Brito, N.M.; Duarte, H.S.S.; Bühner, C. de B.; Auer, C.G.; Santos, Á.F. dos. Morphophysiological Characterization of Ceratocystis Fimbriata Isolates from Yerba Mate. Cienc. Rural 2021, 51. [Google Scholar] [CrossRef]
- De Brito, N.M.; Duarte, H.d.S.S.; Auer, C.G.; Wendling, I.; Alfenas, R.F.; Dos Santos, Á.F. Aggressiveness and Screening for Host Responses in Yerba Mate (Ilex Paraguariensis) Clones to a Wilt Disease Caused by Ceratocystis Fimbriata. For. Pathol. 2021, 51, e12682. [Google Scholar] [CrossRef]
- Engelbrecht, C.J.; Harrington, T.C.; Alfenas, A. Ceratocystis Wilt of Cacao—A Disease of Increasing Importance. Phytopathology 2007, 97, 1648–1649. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, C.J.B.; Harrington, T.C.; Alfenas, A.C.; Suarez, C. Genetic Variation in Populations of the Cacao Wilt Pathogen, Ceratocystis Cacaofunesta. Plant Pathol. 2007, 56, 923–933. [Google Scholar] [CrossRef]
- Suzuki, N.; Ghabrial, S.A.; Kim, K.-H.; Pearson, M.; Marzano, S.-Y.L.; Yaegashi, H.; Xie, J.; Guo, L.; Kondo, H.; Koloniuk, I.; et al. ICTV Virus Taxonomy Profile: Hypoviridae. J. Gen. Virol. 2018, 99, 615–616. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Bian, R.; Liu, Q.; Yang, L.; Pang, T.; Salaipeth, L.; Andika, I.B.; Kondo, H.; Sun, L. Identification of a Novel Hypovirulence-Inducing Hypovirus From Alternaria Alternata. Front. Microbiol. 2019, 10, 1076. [Google Scholar] [CrossRef] [PubMed]
- De Miccolis Angelini, R.M.; Raguseo, C.; Rotolo, C.; Gerin, D.; Faretra, F.; Pollastro, S. The Mycovirome in a Worldwide Collection of the Brown Rot Fungus Monilinia Fructicola. J. Fungi 2022, 8, 481. [Google Scholar] [CrossRef]
- Velasco, L.; Arjona-Girona, I.; Ariza-Fernández, M.T.; Cretazzo, E.; López-Herrera, C. A Novel Hypovirus Species From Xylariaceae Fungi Infecting Avocado. Front. Microbiol. 2018, 9, 778. [Google Scholar] [CrossRef]
- Li, Q.; Huang, W.; Hai, D.; Wang, Y.; Xie, J.; Wang, M. The Complete Genome Sequence of a Novel Hypovirus Infecting Bipolaris Oryzae. Arch. Virol. 2020, 165, 1027–1031. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, J.; Nzabanita, C.; Zhang, M.; Nie, J.; Guo, L. Fungal Virus, FgHV1-Encoded P20 Suppresses RNA Silencing through Single-Strand Small RNA Binding. J. Fungi 2022, 8, 1171. [Google Scholar] [CrossRef]
- Aulia, A.; Hyodo, K.; Hisano, S.; Kondo, H.; Hillman, B.I.; Suzuki, N. Identification of an RNA Silencing Suppressor Encoded by a Symptomless Fungal Hypovirus, Cryphonectria Hypovirus 4. Biology 2021, 10, 100. [Google Scholar] [CrossRef]
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.-M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Pénzes, J.J.; et al. ICTV Virus Taxonomy Profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Q.; Li, S.; Sossah, F.L.; Han, X.; Zhu, G.; Li, Y.; Li, C.; Fu, Y. A Novel Fungal Negative-Stranded RNA Virus Related to Mymonaviruses in Auricularia Heimuer. Arch. Virol. 2022, 167, 2223–2227. [Google Scholar] [CrossRef]
- Hao, F.; Wu, M.; Li, G. Molecular Characterization and Geographic Distribution of a Mymonavirus in the Population of Botrytis Cinerea. Viruses 2018, 10, 432. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M. ICTV Virus Taxonomy Profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yang, B.; Liu, J.; Wang, Q.; Zhang, Q.; Yu, W.; Hsiang, T. A Novel Mycovirus Infecting Aspergillus Nidulans That Is Closely Related to Viruses in a New Genus of the Family Partitiviridae. Arch. Virol. 2021, 166, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, H.; Kotta-Loizou, I.; Kim, K.-H.; Xie, J. Editorial: Mycoviruses and Related Viruses Infecting Fungi, Lower Eukaryotes, Plants and Insects. Front. Microbiol. 2021, 12, 798598. [Google Scholar] [CrossRef]
- Deng, F.; Boland, G.J. Natural Occurrence of a Partitivirus in the Sapstaining Fungus Ceratocystis Resinifera. Can. J. Plant Pathol. 2007, 29, 182–189. [Google Scholar] [CrossRef]
- Botella, L.; Jung, M.H.; Rost, M.; Jung, T. Natural Populations from the Phytophthora Palustris Complex Show a High Diversity and Abundance of SsRNA and DsRNA Viruses. J. Fungi 2022, 8, 1118. [Google Scholar] [CrossRef]
- Kinsella, C.M.; Deijs, M.; Gittelbauer, H.; van der Hoek, L.; van Dijk, K. Human Clinical Isolates of Pathogenic Fungi Are Host to Diverse Mycoviruses. Microbiol. Spectr. 2022, 10, e01610–e01622. [Google Scholar] [CrossRef]
- Zou, Q.; Gao, Y.; Wang, Q.; Yang, Y.; Wang, F.; Hong, N.; Wang, G.; Wang, L. The Full-Length Genome Sequence of a Novel Mitovirus from Botryosphaeria Dothidea, the Causal Agent of Pear Ring Rot Disease. Arch. Virol. 2021, 166, 2881–2885. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.; Zhou, Q. The Complete Genome Sequence of a New Mitovirus from the Phytopathogenic Fungus Colletotrichum Higginsianum. Arch. Virol. 2021, 166, 1481–1484. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ni, Y.; Liu, X.; Zhao, H.; Xiao, Y.; Xiao, X.; Li, S.; Liu, H. Divergent RNA Viruses in Macrophomina Phaseolina Exhibit Potential as Virocontrol Agents. Virus Evol. 2021, 7, veaa095. [Google Scholar] [CrossRef] [PubMed]
- de Rezende, R.R.; de Oliveira Souza, F.; Leal, L.L.; Morgan, T.; Bermudez, J.M.M.; Cascardo, R.d.S.; de Barros, D.R.; Alfenas-Zerbini, P. Characterization of a New Mitovirus Infecting the Phytopathogenic Fungus Microdochium Albescens. Arch. Virol. 2021, 166, 3455–3459. [Google Scholar] [CrossRef] [PubMed]
- Shafik, K.; Umer, M.; You, H.; Aboushedida, H.; Wang, Z.; Ni, D.; Xu, W. Characterization of a Novel Mitovirus Infecting Melanconiella Theae Isolated From Tea Plants. Front Microbiol. 2021, 12, 757556. [Google Scholar] [CrossRef]
- Koonin, E.V.; Dolja, V.V.; Morris, T.J. Evolution and Taxonomy of Positive-Strand RNA Viruses: Implications of Comparative Analysis of Amino Acid Sequences. Crit. Rev. Biochem. Mol. Biol. 1993, 28, 375–430. [Google Scholar] [CrossRef]
- Nishikiori, M.; Sugiyama, S.; Xiang, H.; Niiyama, M.; Ishibashi, K.; Inoue, T.; Ishikawa, M.; Matsumura, H.; Katoh, E. Crystal Structure of the Superfamily 1 Helicase from Tomato Mosaic Virus. J. Virol. 2012, 86, 7565–7576. [Google Scholar] [CrossRef]
- Luo, X.; Jiang, D.; Xie, J.; Jia, J.; Duan, J.; Cheng, J.; Fu, Y.; Chen, T.; Yu, X.; Li, B.; et al. Genome Characterization and Phylogenetic Analysis of a Novel Endornavirus That Infects Fungal Pathogen Sclerotinia Sclerotiorum. Viruses 2022, 14, 456. [Google Scholar] [CrossRef]
- Cao, C.T.H.; Derbyshire, M.C.; Regmi, R.; Li, H.; Jones, M.G.K.; Wylie, S.J. Small RNA Analyses of a Ceratobasidium Isolate Infected with Three Endornaviruses. Viruses 2022, 14, 2276. [Google Scholar] [CrossRef]
- Marais, A.; Faure, C.; Comont, G.; Candresse, T.; Stempien, E.; Corio-Costet, M.-F. Characterization of the Mycovirome of the Phytopathogenic Fungus, Neofusicoccum Parvum. Viruses 2021, 13, 375. [Google Scholar] [CrossRef]
- Koonin, E.V.; Gorbalenya, A.E.; Chumakov, K.M. Tentative Identification of RNA-Dependent RNA Polymerases of DsRNA Viruses and Their Relationship to Positive Strand RNA Viral Polymerases. FEBS Lett. 1989, 252, 42–46. [Google Scholar] [CrossRef]
- Zanotto, P.M.; Gibbs, M.J.; Gould, E.A.; Holmes, E.C. A Reevaluation of the Higher Taxonomy of Viruses Based on RNA Polymerases. J. Virol. 1996, 70, 6083–6096. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Nuss, D.L. A Host Dicer Is Required for Defective Viral RNA Production and Recombinant Virus Vector RNA Instability for a Positive Sense RNA Virus. Proc. Natl. Acad. Sci. USA 2008, 105, 16749–16754. [Google Scholar] [CrossRef] [PubMed]
- Honda, S.; Eusebio-Cope, A.; Miyashita, S.; Yokoyama, A.; Aulia, A.; Shahi, S.; Kondo, H.; Suzuki, N. Establishment of Neurospora Crassa as a Model Organism for Fungal Virology. Nat. Commun. 2020, 11, 5627. [Google Scholar] [CrossRef] [PubMed]
- Maiti, M.; Lee, H.-C.; Liu, Y. QIP, a Putative Exonuclease, Interacts with the Neurospora Argonaute Protein and Facilitates Conversion of Duplex SiRNA into Single Strands. Genes Dev. 2007, 21, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, H.; Liao, X.L.; Gao, B.; Lu, X.; Sun, D.; Gong, W.; Zhong, J.; Zhu, H.; Pan, X.; et al. Mycoviral Gene Integration Converts a Plant Pathogenic Fungus into a Biocontrol Agent. Proc. Natl. Acad. Sci. USA 2022, 119, e2214096119. [Google Scholar] [CrossRef]
- Lee, H.-C.; Chang, S.-S.; Choudhary, S.; Aalto, A.P.; Maiti, M.; Bamford, D.H.; Liu, Y. QiRNA Is a New Type of Small Interfering RNA Induced by DNA Damage. Nature 2009, 459, 274–277. [Google Scholar] [CrossRef]
- de Beer, Z.W.; Duong, T.A.; Barnes, I.; Wingfield, B.D.; Wingfield, M.J. Redefining Ceratocystis and Allied Genera. Stud. Mycol 2014, 79, 187–219. [Google Scholar] [CrossRef]
- Nasution, A.; Glen, M.; Beadle, C.; Mohammed, C. Ceratocystis Wilt and Canker—A Disease That Compromises the Growing of Commercial Acacia-Based Plantations in the Tropics. Aust. For. 2019, 82 (Suppl. S1), 80–93. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espinal, R.B.A.; de Santana, S.F.; Santos, V.C.; Lizardo, G.N.R.; Silva, R.J.S.; Corrêa, R.X.; Loguercio, L.L.; Góes-Neto, A.; Pirovani, C.P.; Fonseca, P.L.C.; et al. Uncovering a Complex Virome Associated with the Cacao Pathogens Ceratocystis cacaofunesta and Ceratocystis fimbriata. Pathogens 2023, 12, 287. https://doi.org/10.3390/pathogens12020287
Espinal RBA, de Santana SF, Santos VC, Lizardo GNR, Silva RJS, Corrêa RX, Loguercio LL, Góes-Neto A, Pirovani CP, Fonseca PLC, et al. Uncovering a Complex Virome Associated with the Cacao Pathogens Ceratocystis cacaofunesta and Ceratocystis fimbriata. Pathogens. 2023; 12(2):287. https://doi.org/10.3390/pathogens12020287
Chicago/Turabian StyleEspinal, Roy Bogardid Ardón, Sabrina Ferreira de Santana, Vinícius Castro Santos, Gabriela Nicolle Ramos Lizardo, Raner José Santana Silva, Ronan Xavier Corrêa, Leandro Lopes Loguercio, Aristóteles Góes-Neto, Carlos Priminho Pirovani, Paula Luize Camargos Fonseca, and et al. 2023. "Uncovering a Complex Virome Associated with the Cacao Pathogens Ceratocystis cacaofunesta and Ceratocystis fimbriata" Pathogens 12, no. 2: 287. https://doi.org/10.3390/pathogens12020287