

The Role of Cluster of Differentiation 39 (CD39) and Purinergic Signaling Pathway in Viral Infections
 , , , and
, , , and
Abstract
:1. Introduction
2. CD39 Role in the Lymphocytic Subsets’ Functions during Viral Infections
3. CD39 Expression and RNA Virus Infections
3.1. The Proportion and Role of CD39+ Cells during HIV Infection
3.2. CD39/73 in Viral Hepatitis
3.2.1. The Level of CD39+ Cells during Hepatitis C Virus (HCV) Infection
3.2.2. The Level of CD39+ Cells during Hepatitis E Virus (HEV) Infection
3.2.3. CD39+ Cells and Hepatitis D Virus (HDV) Infection
3.3. CD39+ Cells and Influenza Infection
3.4. CD39 and Dengue Virus Infection
3.5. CD39+ Cells and Zika Virus Infection
3.6. CD39+ Cells and Lymphocytic Choriomeningitis Virus (LCMV) Infection
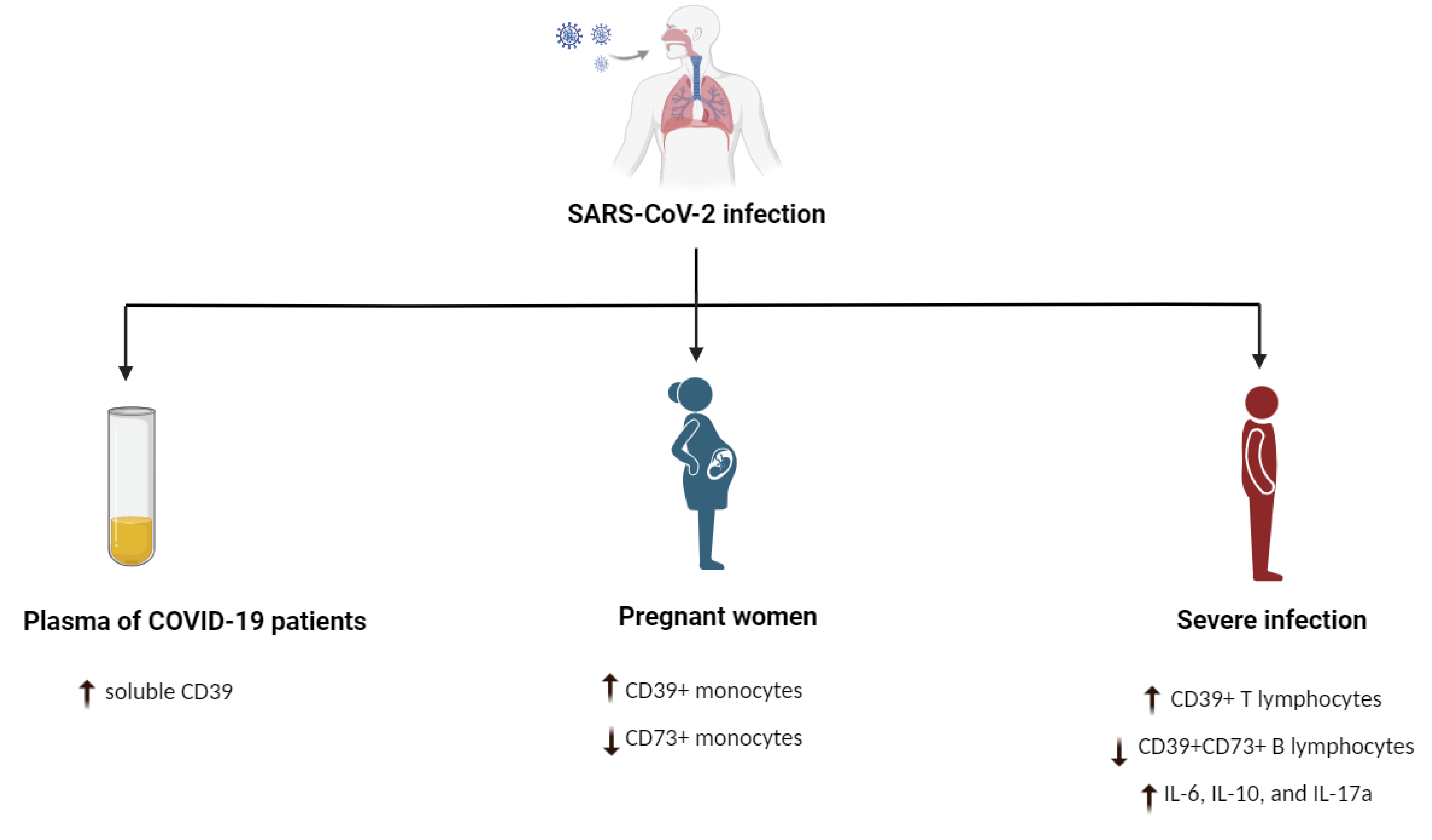
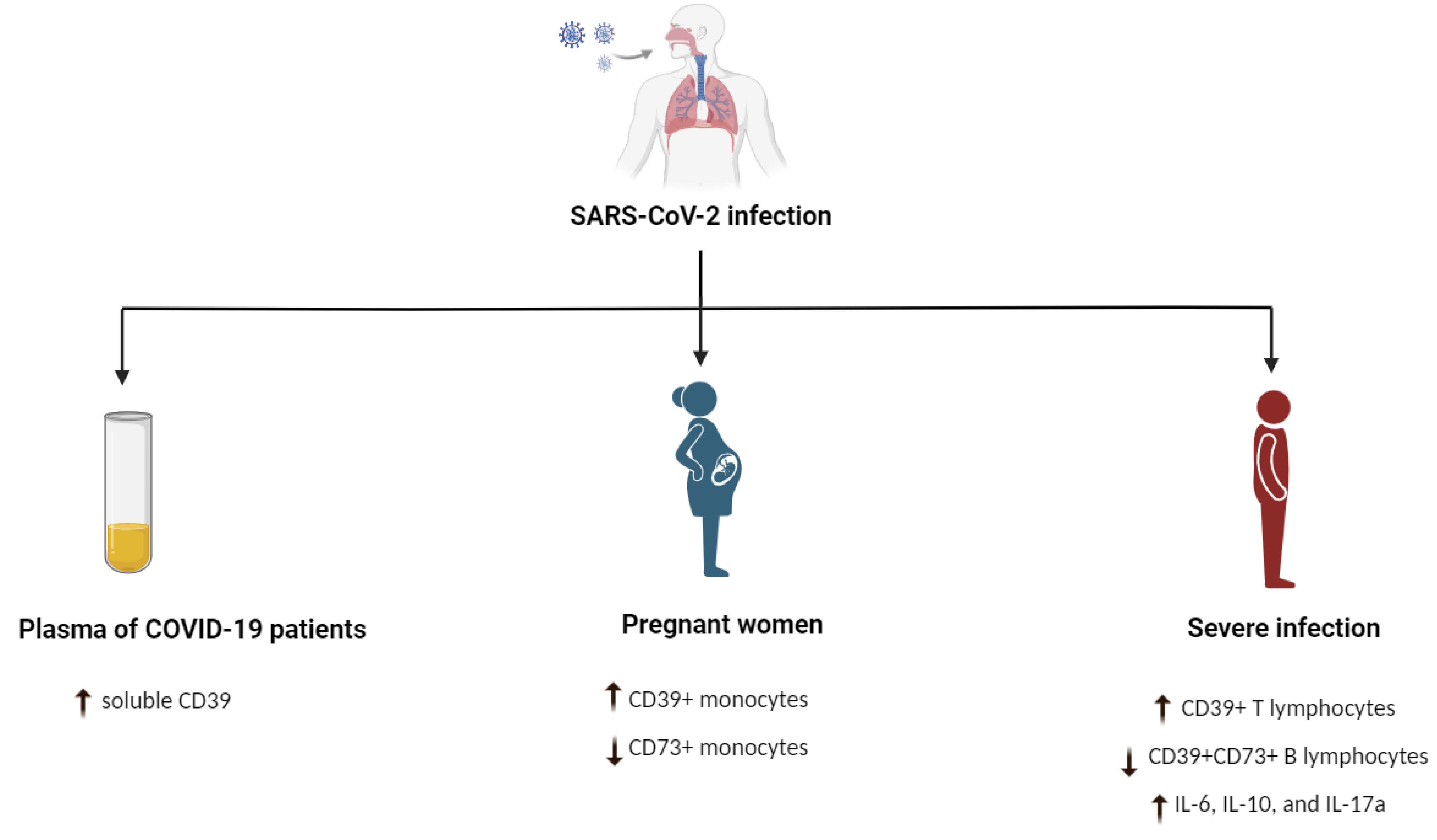
3.7. CD39+ Cells and Severe Acute Respiratory Syndrome (SARS) CoV-2 Infection
4. CD39 + Cells and Infections Caused by DNA Viruses
4.1. CD39+ Cells and Cytomegalovirus (CMV) Infection
4.2. CD39+ Cells and Epstein–Barr Virus (EBV) Infection
4.3. CD39+ Cells and Human Papillomavirus (HPV)
4.4. CD39+ Cells and Hepatitis B Virus (HBV) Infection
5. CD39-Positive Cells and HIV/Tuberculosis (TB) Coinfection
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Robson, S.C.; Sévigny, J.; Zimmermann, H. The E-NTPDase family of ectonucleotidases: Structure function relationships and pathophysiological significance. Purinergic Signal. 2006, 2, 409–430. [Google Scholar] [CrossRef] [PubMed]
- Maliszewski, C.R.; Delespesse, G.J.; Schoenborn, M.A.; Armitage, R.J.; Fanslow, W.C.; Nakajima, T.; Baker, E.; Sutherland, G.R.; Poindexter, K.; Birks, C.; et al. The CD39 lymphoid cell activation antigen. Molecular cloning and structural characterization. J. Immunol. 1994, 153, 3574–3583. [Google Scholar] [CrossRef] [PubMed]
- Kansas, G.S.; Wood, G.S.; Tedder, T.F. Expression, distribution, and biochemistry of human CD39. Role in activation-associated homotypic adhesion of lymphocytes. J. Immunol. 1991, 146, 2235–2244. [Google Scholar] [CrossRef] [PubMed]
- Pulte, D.; Furman, R.R.; Broekman, M.J.; Drosopoulos, J.H.; Ballard, H.S.; Olson, K.E.; Kizer, J.R.; Marcus, A.J. CD39 expression on T lymphocytes correlates with severity of disease in patients with chronic lymphocytic leukemia. Clin. Lymphoma Myeloma Leuk. 2011, 11, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G.; Wieringa, B.; Robson, S.C.; Jalkanen, S. Metabolism of circulating ADP in the bloodstream is mediated via integrated actions of soluble adenylate kinase-1 and NTPDase1/CD39 activities. Faseb J. 2012, 26, 3875–3883. [Google Scholar] [CrossRef]
- Wang, T.-F.; Guidotti, G. CD39 Is an Ecto-(Ca2+, Mg2+)-apyrase (*). J. Biol. Chem. 1996, 271, 9898–9901. [Google Scholar] [CrossRef]
- Burnstock, G. Purine and purinergic receptors. Brain Neurosci. Adv. 2018, 2, 2398212818817494. [Google Scholar] [CrossRef]
- Sueyoshi, K.; Ledderose, C.; Shen, Y.; Lee, A.H.; Shapiro, N.I.; Junger, W.G. Lipopolysaccharide suppresses T cells by generating extracellular ATP that impairs their mitochondrial function via P2Y11 receptors. J. Biol. Chem. 2019, 294, 6283–6293. [Google Scholar] [CrossRef]
- Young, A.; Ngiow, S.F.; Barkauskas, D.S.; Sult, E.; Hay, C.; Blake, S.J.; Huang, Q.; Liu, J.; Takeda, K.; Teng, M.W. Co-inhibition of CD73 and A2AR adenosine signaling improves anti-tumor immune responses. Cancer Cell 2016, 30, 391–403. [Google Scholar] [CrossRef]
- Harvey, J.B.; Phan, L.H.; Villarreal, O.E.; Bowser, J.L. CD73’s potential as an immunotherapy target in gastrointestinal cancers. Front. Immunol. 2020, 11, 508. [Google Scholar] [CrossRef] [Green Version]
- Powderly, J.D.; de Souza, P.L.; Gutierrez, R.; Horvath, L.; Seitz, L.; Ashok, D.; Park, A.; Walters, M.J.; Karakunnel, J.J.; Berry, W. AB928, a novel dual adenosine receptor antagonist, combined with chemotherapy or AB122 (anti-PD-1) in patients (pts) with advanced tumors: Preliminary results from ongoing phase I studies. Am. Soc. Clin. Oncol. 2019, 37, 2604. [Google Scholar] [CrossRef]
- Willingham, S.B.; Ho, P.Y.; Hotson, A.; Hill, C.; Piccione, E.C.; Hsieh, J.; Liu, L.; Buggy, J.J.; McCaffery, I.; Miller, R.A. A2AR Antagonism with CPI-444 Induces Antitumor Responses and Augments Efficacy to Anti–PD-(L) 1 and Anti–CTLA-4 in Preclinical ModelsA2AR Antagonism with CPI-444 Stimulates Antitumor Immunity. Cancer Immunol. Res. 2018, 6, 1136–1149. [Google Scholar] [CrossRef]
- Fredholm, B.B.; AP, I.J.; Jacobson, K.A.; Linden, J.; Müller, C.E. International Union of Basic and Clinical Pharmacology. LXXXI. Nomenclature and classification of adenosine receptors—An update. Pharmacol. Rev. 2011, 63, 1–34. [Google Scholar] [CrossRef]
- Müller, C.E.; Jacobson, K.A. Recent developments in adenosine receptor ligands and their potential as novel drugs. Biochim. Biophys. Acta 2011, 1808, 1290–1308. [Google Scholar] [CrossRef]
- Barletta, K.E.; Ley, K.; Mehrad, B. Regulation of neutrophil function by adenosine. Arter. Thromb Vasc Biol. 2012, 32, 856–864. [Google Scholar] [CrossRef]
- Cohen, H.B.; Ward, A.; Hamidzadeh, K.; Ravid, K.; Mosser, D.M. IFN-γ prevents adenosine receptor (A2bR) upregulation to sustain the macrophage activation response. J. Immunol. 2015, 195, 3828–3837. [Google Scholar] [CrossRef]
- Cohen, H.B.; Briggs, K.T.; Marino, J.P.; Ravid, K.; Robson, S.C.; Mosser, D.M. TLR stimulation initiates a CD39-based autoregulatory mechanism that limits macrophage inflammatory responses. Blood 2013, 122, 1935–1945. [Google Scholar] [CrossRef]
- Haskó, G.; Pacher, P. Regulation of macrophage function by adenosine. Arter. Thromb Vasc Biol. 2012, 32, 865–869. [Google Scholar] [CrossRef]
- Kronlage, M.; Song, J.; Sorokin, L.; Isfort, K.; Schwerdtle, T.; Leipziger, J.; Robaye, B.; Conley, P.B.; Kim, H.-C.; Sargin, S.J. Autocrine purinergic receptor signaling is essential for macrophage chemotaxis. Sci. Signal. 2010, 3, ra55. [Google Scholar] [CrossRef]
- Dombrowski, K.E.; Ke, Y.; Thompson, L.F.; Kapp, J.A. Antigen recognition by CTL is dependent upon ectoATPase activity. J. Immunol. 1995, 154, 6227–6237. [Google Scholar] [CrossRef]
- Gupta, P.K.; Godec, J.; Wolski, D.; Adland, E.; Yates, K.; Pauken, K.E.; Cosgrove, C.; Ledderose, C.; Junger, W.G.; Robson, S.C.; et al. CD39 Expression Identifies Terminally Exhausted CD8+ T Cells. PLOS Pathog. 2015, 11, e1005177. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, M.; Moulana, Z.; Sepidarkish, M.; Bagherzadeh, M.; Rezanejad, M.; Mirzakhani, M.; Jafari, M.; Mohammadnia-Afrouzi, M. Pronounce expression of Tim-3 and CD39 but not PD1 defines CD8 T cells in critical Covid-19 patients. Microb. Pathog. 2021, 153, 104779. [Google Scholar] [CrossRef] [PubMed]
- Canale, F.P.; Ramello, M.C.; Núñez, N.; Araujo Furlan, C.L.; Bossio, S.N.; Gorosito Serrán, M.; Tosello Boari, J.; Del Castillo, A.; Ledesma, M.; Sedlik, C.; et al. CD39 Expression Defines Cell Exhaustion in Tumor-Infiltrating CD8(+) T Cells. Cancer Res. 2018, 78, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Xia, Y.; Lin, Z.; Qu, Y.; Qi, Y.; Chen, Y.; Zhou, Q.; Zeng, H.; Wang, J.; Chang, Y.; et al. Tumor-infiltrating CD39(+)CD8(+) T cells determine poor prognosis and immune evasion in clear cell renal cell carcinoma patients. Cancer Immunol. Immunother. 2020, 69, 1565–1576. [Google Scholar] [CrossRef]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Sakaguchi, N.; Asano, M.; Itoh, M.; Toda, M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 1995, 155, 1151–1164. [Google Scholar] [CrossRef]
- Jung, M.K.; Shin, E.C. Regulatory T Cells in Hepatitis B and C Virus Infections. Immune Netw. 2016, 16, 330–336. [Google Scholar] [CrossRef]
- Dietze, K.K.; Schimmer, S.; Kretzmer, F.; Wang, J.; Lin, Y.; Huang, X.; Wu, W.; Wang, B.; Lu, M.; Dittmer, U. Characterization of the Treg response in the hepatitis B virus hydrodynamic injection mouse model. PLoS ONE 2016, 11, e0151717. [Google Scholar] [CrossRef]
- Tang, Y.; Jiang, L.; Zheng, Y.; Ni, B.; Wu, Y. Expression of CD39 on FoxP3+ T regulatory cells correlates with progression of HBV infection. BMC Immunol. 2012, 13, 17. [Google Scholar] [CrossRef]
- Dwyer, K.M.; Hanidziar, D.; Putheti, P.; Hill, P.A.; Pommey, S.; McRae, J.L.; Winterhalter, A.; Doherty, G.; Deaglio, S.; Koulmanda, M.; et al. Expression of CD39 by human peripheral blood CD4+ CD25+ T cells denotes a regulatory memory phenotype. Am. J. Transpl. 2010, 10, 2410–2420. [Google Scholar] [CrossRef] [Green Version]
- Schulze zur Wiesch, J.; Thomssen, A.; Hartjen, P.; Toth, I.; Lehmann, C.; Meyer-Olson, D.; Colberg, K.; Frerk, S.; Babikir, D.; Schmiedel, S.; et al. Comprehensive Analysis of Frequency and Phenotype of T Regulatory Cells in HIV Infection: CD39 Expression of FoxP3+ T Regulatory Cells Correlates with Progressive Disease. J. Virol. 2010, 85, 1287–1297. [Google Scholar] [CrossRef]
- Ouyang, W.; Kolls, J.K.; Zheng, Y. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity 2008, 28, 454–467. [Google Scholar] [CrossRef]
- Otsuka, A.; Hanakawa, S.; Miyachi, Y.; Kabashima, K. CD39: A new surface marker of mouse regulatory γδ T cells. J. Allergy Clin. Immunol. 2013, 132, 1448–1451. [Google Scholar] [CrossRef]
- Kolbe, K.; Wittner, M.; Hartjen, P.; Hüfner, A.D.; Degen, O.; Ackermann, C.; Cords, L.; Stellbrink, H.J.; Haag, F.; Schulze Zur Wiesch, J. Inversed Ratio of CD39/CD73 Expression on γδ T Cells in HIV Versus Healthy Controls Correlates With Immune Activation and Disease Progression. Front. Immunol. 2022, 13, 867167. [Google Scholar] [CrossRef]
- Mizumoto, N.; Kumamoto, T.; Robson, S.C.; Sévigny, J.; Matsue, H.; Enjyoji, K.; Takashima, A. CD39 is the dominant Langerhans cell–associated ecto-NTPDase: Modulatory roles in inflammation and immune responsiveness. Nat. Med. 2002, 8, 358–365. [Google Scholar] [CrossRef]
- Kaczmarek, E.; Koziak, K.; Sévigny, J.; Siegel, J.B.; Anrather, J.; Beaudoin, A.R.; Bach, F.H.; Robson, S.C. Identification and characterization of CD39/vascular ATP diphosphohydrolase. J. Biol. Chem. 1996, 271, 33116–33122. [Google Scholar] [CrossRef]
- Dierks, P.; Wroblewski, R.; Eberhard, J.M.; Martrus, G.; Degen, O.; Hertling, S.; Schmiedel, S.; Lunemann, S.; Hüfner, A.; Lohse, A.W.; et al. Brief Report: Increased Frequency of CD39+ CD56bright Natural Killer Cells in HIV-1 Infection Correlates With Immune Activation and Disease Progression. J. Acquir. Immune. Defic. Syndr. 2017, 74, 467–472. [Google Scholar] [CrossRef]
- Li, J.; Huang, H.H.; Tu, B.; Zhou, M.J.; Hu, W.; Fu, Y.L.; Li, X.Y.; Yang, T.; Song, J.W.; Fan, X.; et al. Reversal of the CD8(+) T-Cell Exhaustion Induced by Chronic HIV-1 Infection Through Combined Blockade of the Adenosine and PD-1 Pathways. Front. Immunol. 2021, 12, 687296. [Google Scholar] [CrossRef]
- Jenabian, M.-A.; Seddiki, N.; Yatim, A.; Carriere, M.; Hulin, A.; Younas, M.; Ghadimi, E.; Kök, A.; Routy, J.-P.; Tremblay, A. Regulatory T cells negatively affect IL-2 production of effector T cells through CD39/adenosine pathway in HIV infection. PLoS Pathog. 2013, 9, e1003319. [Google Scholar] [CrossRef]
- Fenoglio, D.; Dentone, C.; Signori, A.; Di Biagio, A.; Parodi, A.; Kalli, F.; Nasi, G.; Curto, M.; Cenderello, G.; De Leo, P.; et al. CD8(+)CD28(−)CD127(lo)CD39(+) regulatory T-cell expansion: A new possible pathogenic mechanism for HIV infection? J. Allergy Clin. Immunol. 2018, 141, 2220–2233. [Google Scholar] [CrossRef] [Green Version]
- Seddiki, N.; Cook, L.; Hsu, D.C.; Phetsouphanh, C.; Brown, K.; Xu, Y.; Kerr, S.J.; Cooper, D.A.; Munier, C.M.; Pett, S.; et al. Human antigen-specific CD4⁺ CD25⁺ CD134⁺ CD39⁺ T cells are enriched for regulatory T cells and comprise a substantial proportion of recall responses. Eur. J. Immunol. 2014, 44, 1644–1661. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Ackermann, C.; Tóth, I.; Dierks, P.; Eberhard, J.M.; Wroblewski, R.; Scherg, F.; Geyer, M.; Schmidt, R.E.; Beisel, C.; et al. Down-regulation of CD73 on B cells of patients with viremic HIV correlates with B cell activation and disease progression. J. Leukoc Biol. 2017, 101, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Cachem, F.; Dias, A.S.; Monteiro, C.; Fernandes, G.; Delphim, L.; Tavares, F.; Maciel, A.M.A.; Amendola-Pires, M.M.; Brandão-Mello, C.E.; Andrade, R.M.; et al. Different core-specific T cell subsets are expanded in chronic hepatitis C with advanced liver disease. Cytokine 2019, 124, 154456. [Google Scholar] [CrossRef] [PubMed]
- Keoshkerian, E.; Hunter, M.; Cameron, B.; Nguyen, N.; Sugden, P.; Bull, R.; Zekry, A.; Maher, L.; Seddiki, N.; Zaunders, J. Hepatitis C-specific effector and regulatory CD 4 T-cell responses are associated with the outcomes of primary infection. J. Viral Hepat. 2016, 23, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Wildner, N.H.; Walker, A.; Brauneck, F.; Ditt, V.; Peine, S.; Huber, S.; Haag, F.; Beisel, C.; Timm, J.; Schulze Zur Wiesch, J. Transcriptional Pattern Analysis of Virus-Specific CD8+ T Cells in Hepatitis C Infection: Increased Expression of TOX and Eomesodermin During and After Persistent Antigen Recognition. Front. Immunol. 2022, 13, 886646. [Google Scholar] [CrossRef]
- Rathod, S.B.; Das, R.; Thanapati, S.; Arankalle, V.A.; Tripathy, A.S. Suppressive activity and altered conventional phenotype markers/mediators of regulatory T cells in patients with self-limiting hepatitis E. J. Viral Hepat. 2014, 21, 141–151. [Google Scholar] [CrossRef]
- Taylor, J.M.; Han, Z. Purinergic receptor functionality is necessary for infection of human hepatocytes by hepatitis delta virus and hepatitis B virus. PLoS ONE 2010, 5, e15784. [Google Scholar] [CrossRef]
- Lu, C.; Zanker, D.; Lock, P.; Jiang, X.; Deng, J.; Duan, M.; Liu, C.; Faou, P.; Hickey, M.J.; Chen, W. Memory regulatory T cells home to the lung and control influenza A virus infection. Immunol. Cell Biol. 2019, 97, 774–786. [Google Scholar] [CrossRef]
- Hall, O.J.; Limjunyawong, N.; Vermillion, M.S.; Robinson, D.P.; Wohlgemuth, N.; Pekosz, A.; Mitzner, W.; Klein, S.L. Progesterone-Based Therapy Protects Against Influenza by Promoting Lung Repair and Recovery in Females. PLoS Pathog. 2016, 12, e1005840. [Google Scholar] [CrossRef]
- Tsai, C.-Y.; Liong, K.H.; Gunalan, M.G.; Li, N.; Lim, D.S.L.; Fisher, D.A.; MacAry, P.A.; Leo, Y.S.; Wong, S.-C.; Puan, K.J. Type I IFNs and IL-18 regulate the antiviral response of primary human γδ T cells against dendritic cells infected with Dengue virus. J. Immunol. 2015, 194, 3890–3900. [Google Scholar] [CrossRef] [Green Version]
- Guerra-Gomes, I.C.; Gois, B.M.; Peixoto, R.F.; Palmeira, P.H.S.; Dias, C.N.S.; Csordas, B.G.; Araújo, J.M.G.; Veras, R.C.; de Medeiros, I.A.; de Azevedo, F.; et al. Phenotypical characterization of regulatory T cells in acute Zika infection. Cytokine 2021, 146, 155651. [Google Scholar] [CrossRef]
- Pietrobon, A.J.; Andrejew, R.; Custódio, R.W.A.; Oliveira, L.M.; Scholl, J.N.; Teixeira, F.M.E.; de Brito, C.A.; Glaser, T.; Kazmierski, J.; Goffinet, C.; et al. Dysfunctional purinergic signaling correlates with disease severity in COVID-19 patients. Front. Immunol. 2022, 13, 1012027. [Google Scholar] [CrossRef]
- Cérbulo-Vázquez, A.; García-Espinosa, M.; Briones-Garduño, J.C.; Arriaga-Pizano, L.; Ferat-Osorio, E.; Zavala-Barrios, B.; Cabrera-Rivera, G.L.; Miranda-Cruz, P.; García de la Rosa, M.T.; Prieto-Chávez, J.L.; et al. The percentage of CD39+ monocytes is higher in pregnant COVID-19+ patients than in nonpregnant COVID-19+ patients. PLoS ONE 2022, 17, e0264566. [Google Scholar] [CrossRef]
- Dorneles, G.P.; Teixeira, P.C.; da Silva, I.M.; Schipper, L.L.; Santana Filho, P.C.; Rodrigues Junior, L.C.; Bonorino, C.; Peres, A.; Fonseca, S.G.; Monteiro, M.C.; et al. Alterations in CD39/CD73 axis of T cells associated with COVID-19 severity. J. Cell Physiol. 2022, 237, 3394–3407. [Google Scholar] [CrossRef]
- Díaz-García, E.; García-Tovar, S.; Alfaro, E.; Zamarrón, E.; Mangas, A.; Galera, R.; Ruíz-Hernández, J.J.; Solé-Violán, J.; Rodríguez-Gallego, C.; Van-Den-Rym, A.; et al. Role of CD39 in COVID-19 Severity: Dysregulation of Purinergic Signaling and Thromboinflammation. Front. Immunol. 2022, 13, 847894. [Google Scholar] [CrossRef]
- Luciw, P.A. Human immunodeficiency viruses and their replication. In Fields Virology, 3rd ed.; Fields, B.N., Knipe, D.M., Eds.; Lippincott-Raven Publishers: Philadelphia, PA, USA, 1996; pp. 1881–1952. [Google Scholar]
- Shaw, G.M.; Hunter, E. HIV transmission. Cold Spring Harb. Perspect. Med. 2012, 2, a006965. [Google Scholar] [CrossRef]
- Okoye, A.A.; Picker, L.J. CD4(+) T-cell depletion in HIV infection: Mechanisms of immunological failure. Immunol Rev. 2013, 254, 54–64. [Google Scholar] [CrossRef]
- Leal, D.B.; Streher, C.A.; Bertoncheli, C.d.M.; Carli, L.F.; Leal, C.A.; da Silva, J.E.; Morsch, V.M.; Schetinger, M.R. HIV infection is associated with increased NTPDase activity that correlates with CD39-positive lymphocytes. Biochim. Et Biophys. Acta (BBA) Mol. Cell Res. 2005, 1746, 129–134. [Google Scholar] [CrossRef]
- Leal, D.B.; Schetinger, M.R.; Leal, C.A.; Bertoncheli Cde, M.; Morsch, V.M. NTPDase activity in human lymphocytes is not affected by therapeutic doses of anti-HIV drugs. Biomed. Pharm. 2011, 65, 594–596. [Google Scholar] [CrossRef]
- Barat, C.; Martin, G.; Beaudoin, A.R.; Sévigny, J.; Tremblay, M.J. The nucleoside triphosphate diphosphohydrolase-1/CD39 is incorporated into human immunodeficiency type 1 particles, where it remains biologically active. J. Mol. Biol. 2007, 371, 269–282. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Fernandez, M.E.; Rueda, C.M.; Rusie, L.K.; Chougnet, C.A. Regulatory T cells control HIV replication in activated T cells through a cAMP-dependent mechanism. Blood 2011, 117, 5372–5380. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, M.; Carriere, M.; Jenabian, M.-A.; Limou, S.; Younas, M.; Kök, A.; Huë, S.; Seddiki, N.; Hulin, A.; Delaneau, O. CD39/adenosine pathway is involved in AIDS progression. PLoS Pathog. 2011, 7, e1002110. [Google Scholar] [CrossRef] [PubMed]
- Yero, A.; Shi, T.; Farnos, O.; Routy, J.P.; Tremblay, C.; Durand, M.; Tsoukas, C.; Costiniuk, C.T.; Jenabian, M.A. Dynamics and epigenetic signature of regulatory T-cells following antiretroviral therapy initiation in acute HIV infection. EBioMedicine 2021, 71, 103570. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-W.; Huang, H.-H.; Zhang, C.; Yang, H.-G.; Zhang, J.-Y.; Xu, R.-N.; Jin, L.; Shi, M.; Wang, F.-S.; Jiao, Y.-M. Expression of CD39 Is Correlated With HIV DNA Levels in Naïve Tregs in Chronically Infected ART Naïve Patients. Front. Immunol. 2019, 10, 02465. [Google Scholar] [CrossRef]
- Hoffmann, M.; Pantazis, N.; Martin, G.E.; Hickling, S.; Hurst, J.; Meyerowitz, J.; Willberg, C.B.; Robinson, N.; Brown, H.; Fisher, M.; et al. Exhaustion of Activated CD8 T Cells Predicts Disease Progression in Primary HIV-1 Infection. PLoS Pathog. 2016, 12, e1005661. [Google Scholar] [CrossRef]
- Schachter, J.; Delgado, K.V.; Barreto-de-Souza, V.; Bou-Habib, D.C.; Persechini, P.M.; Meyer-Fernandes, J.R. Inhibition of ecto-ATPase activities impairs HIV-1 infection of macrophages. Immunobiology 2015, 220, 589–596. [Google Scholar] [CrossRef]
- Cao, W.J.; Zhang, X.C.; Wan, L.Y.; Li, Q.Y.; Mu, X.Y.; Guo, A.L.; Zhou, M.J.; Shen, L.L.; Zhang, C.; Fan, X.; et al. Immune Dysfunctions of CD56(neg) NK Cells Are Associated With HIV-1 Disease Progression. Front. Immunol. 2021, 12, 811091. [Google Scholar] [CrossRef]
- Qian, S.; Xiong, C.; Wang, M.; Zhang, Z.; Fu, Y.; Hu, Q.; Ding, H.; Han, X.; Shang, H.; Jiang, Y. CD38(+)CD39(+) NK cells associate with HIV disease progression and negatively regulate T cell proliferation. Front. Immunol. 2022, 13, 946871. [Google Scholar] [CrossRef]
- Catanese, M.T.; Uryu, K.; Kopp, M.; Edwards, T.J.; Andrus, L.; Rice, W.J.; Silvestry, M.; Kuhn, R.J.; Rice, C.M. Ultrastructural analysis of hepatitis C virus particles. Proc. Natl. Acad. Sci. USA 2013, 110, 9505–9510. [Google Scholar] [CrossRef]
- Alter, H.J.; Seeff, L.B. Recovery, persistence, and sequelae in hepatitis C virus infection: A perspective on long-term outcome. Semin. Liver Dis. 2000, 20, 17–35. [Google Scholar] [CrossRef] [Green Version]
- Abdelwahab, S.F. Cellular immune response to hepatitis-C-virus in subjects without viremia or seroconversion: Is it important? Infect. Agents Cancer 2016, 11, 23. [Google Scholar] [CrossRef]
- Li, Z.; Ping, Y.; Yu, Z.; Wang, M.; Yue, D.; Zhang, Z.; Li, J.; Zhang, B.; Shi, X.; Zhang, Y. Dynamic changes in CD45RA(-)Foxp3(high) regulatory T-cells in chronic hepatitis C patients during antiviral therapy. Int. J. Infect. Dis. 2016, 45, 5–12. [Google Scholar] [CrossRef]
- Sayed, I.M.; Vercouter, A.S.; Abdelwahab, S.F.; Vercauteren, K.; Meuleman, P. Is hepatitis E virus an emerging problem in industrialized countries? Hepatology 2015, 62, 1883–1892. [Google Scholar] [CrossRef]
- Sayed, I.M.; Vercauteren, K.; Abdelwahab, S.F.; Meuleman, P. The emergence of hepatitis E virus in Europe. Future Virol. 2015, 10, 763–778. [Google Scholar] [CrossRef]
- Sayed, I.M.; Vercouter, A.S.; Meuleman, P. Hepatitis E virus in acute liver failure: An unusual suspect? Hepatology 2016, 64, 1837–1839. [Google Scholar] [CrossRef]
- Kamar, N.; Selves, J.; Mansuy, J.M.; Ouezzani, L.; Péron, J.M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E virus and chronic hepatitis in organ-transplant recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef]
- Pischke, S.; Hartl, J.; Pas, S.D.; Lohse, A.W.; Jacobs, B.C.; Van der Eijk, A.A. Hepatitis E virus: Infection beyond the liver? J. Hepatol. 2017, 66, 1082–1095. [Google Scholar] [CrossRef]
- Tripathy, A.S.; Das, R.; Rathod, S.B.; Gurav, Y.K.; Arankalle, V.A. Peripheral T regulatory cells and cytokines in hepatitis E infection. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 179–184. [Google Scholar] [CrossRef]
- Deaglio, S.; Dwyer, K.M.; Gao, W.; Friedman, D.; Usheva, A.; Erat, A.; Chen, J.-F.; Enjyoji, K.; Linden, J.; Oukka, M. Adenosine generation catalyzed by CD39 and CD73 expressed on regulatory T cells mediates immune suppression. J. Exp. Med. 2007, 204, 1257–1265. [Google Scholar] [CrossRef]
- Rathod, S.B.; Tripathy, A.S. TGF-β(1) and contact mediated suppression by CD4+CD25+CD127− T regulatory cells of patients with self-limiting hepatitis E. Hum. Immunol. 2016, 77, 1254–1263. [Google Scholar] [CrossRef]
- Koytak, E.S.; Yurdaydin, C.; Glenn, J.S. Hepatitis D. Curr. Treat. Options Gastroenterol. 2007, 10, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Seeger, C.; Zoulim, F.; Mason, W. Hepadnaviruses. Fields Virol. 2007, 2, 2977–3029. [Google Scholar]
- Negro, F. Hepatitis D virus coinfection and superinfection. Cold Spring Harb. Perspect. Med. 2014, 4, a021550. [Google Scholar] [CrossRef] [PubMed]
- Rizzetto, M.; Canese, M.G.; Arico, S.; Crivelli, O.; Trepo, C.; Bonino, F.; Verme, G. Immunofluorescence detection of new antigen-antibody system (delta/anti-delta) associated to hepatitis B virus in liver and in serum of HBsAg carriers. Gut 1977, 18, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Corriden, R.; Insel, P.A. Basal release of ATP: An autocrine-paracrine mechanism for cell regulation. Sci. Signal. 2010, 3, re1. [Google Scholar] [CrossRef]
- Faas, M.; Sáez, T.; De Vos, P. Extracellular ATP and adenosine: The Yin and Yang in immune responses? Mol. Asp. Med. 2017, 55, 9–19. [Google Scholar] [CrossRef]
- Arbeitskreis, B. Untergruppe «Bewertung Blutassoziierter Krankheitserreger». Influenza Virus. Transfus. Med. Hemother. 2009, 36, 32–39. [Google Scholar]
- Zambon, M.C. The pathogenesis of influenza in humans. Rev. Med. Virol. 2001, 11, 227–241. [Google Scholar] [CrossRef]
- Saito, T.; Tanikawa, T.; Uchida, Y.; Takemae, N.; Kanehira, K.; Tsunekuni, R. Intracontinental and intercontinental dissemination of Asian H5 highly pathogenic avian influenza virus (clade 2.3.4.4) in the winter of 2014–2015. Rev. Med. Virol. 2015, 25, 388–405. [Google Scholar] [CrossRef]
- Aeffner, F.; Woods, P.S.; Davis, I.C. Ecto-5’-nucleotidase CD73 modulates the innate immune response to influenza infection but is not required for development of influenza-induced acute lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 309, L1313–L1322. [Google Scholar] [CrossRef]
- Zimmermann, H.; Zebisch, M.; Sträter, N. Cellular function and molecular structure of ecto-nucleotidases. Purinergic Signal. 2012, 8, 437–502. [Google Scholar] [CrossRef]
- Woods, P.S.; Doolittle, L.M.; Hickman-Davis, J.M.; Davis, I.C. ATP catabolism by tissue nonspecific alkaline phosphatase contributes to development of ARDS in influenza-infected mice. Am. J. Physiol -Lung Cell. Mol. Physiol. 2018, 314, L83–L92. [Google Scholar] [CrossRef]
- Leyva-Grado, V.H.; Ermler, M.E.; Schotsaert, M.; Gonzalez, M.G.; Gillespie, V.; Lim, J.K.; García-Sastre, A. Contribution of the Purinergic Receptor P2X7 to Development of Lung Immunopathology during Influenza Virus Infection. mBio 2017, 8, e00229-17. [Google Scholar] [CrossRef]
- Weinberg, A.; Muresan, P.; Richardson, K.M.; Fenton, T.; Dominguez, T.; Bloom, A.; Watts, D.H.; Abzug, M.J.; Nachman, S.A.; Levin, M.J. Determinants of vaccine immunogenicity in HIV-infected pregnant women: Analysis of B and T cell responses to pandemic H1N1 monovalent vaccine. PLoS ONE 2015, 10, e0122431. [Google Scholar] [CrossRef]
- Weinberg, A.; Muresan, P.; Richardson, K.; Fenton, T.; Dominguez, T.; Bloom, A.; Watts, D.H.; Abzug, M.J.; Nachman, S.A.; Levin, M.J. Heterogeneity of T Cell Responses to Pandemic pH1N1 Monovalent Vaccine in HIV-Infected Pregnant Women. AIDS Res. Hum. Retrovir. 2015, 31, 1170–1177. [Google Scholar] [CrossRef]
- Mathew, A.; Rothman, A.L. Understanding the contribution of cellular immunity to dengue disease pathogenesis. Immunol. Rev. 2008, 225, 300–313. [Google Scholar] [CrossRef]
- Gubler, D.J. Dengue and dengue hemorrhagic fever. Clin. Microbiol. Rev. 1998, 11, 480–496. [Google Scholar] [CrossRef]
- Malhão, T.A.; Resende, C.M.; Gamerman, D.; Medronho Rde, A. [A Bayesian model to investigate excess mortality during the dengue epidemic in Greater Metropolitan Rio de Janeiro, Brazil, in 2007–2008]. Cad. Saude Publica 2013, 29, 2057–2070. [Google Scholar] [CrossRef]
- Corrêa, G.; Lindenberg, C.d.A.; Fernandes-Santos, C.; Gandini, M.; Paiva, F.P.; Coutinho-Silva, R.; Kubelka, C.F. The purinergic receptor P2X7 role in control of Dengue virus-2 infection and cytokine/chemokine production in infected human monocytes. Immunobiology 2016, 221, 794–802. [Google Scholar] [CrossRef]
- Patkar, C.; Giaya, K.; Libraty, D.H. Dengue virus type 2 modulates endothelial barrier function through CD73. Am. J. Trop. Med. Hyg. 2013, 88, 89. [Google Scholar] [CrossRef]
- Zhang, X.; Li, G.; Chen, G.; Zhu, N.; Wu, D.; Wu, Y.; James, T.D. Recent progresses and remaining challenges for the detection of Zika virus. Med. Res. Rev. 2021, 41, 2039–2108. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Novella, I.S.; Teng, M.N.; Oldstone, M.B.; de La Torre, J.C. NP and L proteins of lymphocytic choriomeningitis virus (LCMV) are sufficient for efficient transcription and replication of LCMV genomic RNA analogs. J. Virol. 2000, 74, 3470–3477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldstone, M.B.A.; Ahmed, R.; Byrne, J.; Buchmeier, M.J.; Riviere, Y.; Southern, P. Virus and immune responses: Lymphocytic choriomeningitis virus as a prototype model of viral pathogenesis. Br. Med. Bull. 1985, 41, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Bonthius, D.J. Lymphocytic choriomeningitis virus: An underrecognized cause of neurologic disease in the fetus, child, and adult. Semin. Pediatr. Neurol. 2012, 19, 89–95. [Google Scholar] [CrossRef]
- Schøller, A.S.; Nazerai, L.; Christensen, J.P.; Thomsen, A.R. Functionally Competent, PD-1(+) CD8(+) Trm Cells Populate the Brain Following Local Antigen Encounter. Front. Immunol. 2020, 11, 595707. [Google Scholar] [CrossRef]
- Rothan, H.A.; Byrareddy, S.N. The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J. Autoimmun. 2020, 109, 102433. [Google Scholar] [CrossRef]
- Hatmal, M.m.M.; Alshaer, W.; Al-Hatamleh, M.A.; Hatmal, M.; Smadi, O.; Taha, M.O.; Oweida, A.J.; Boer, J.C.; Mohamud, R.; Plebanski, M. Comprehensive structural and molecular comparison of spike proteins of SARS-CoV-2, SARS-CoV and MERS-CoV, and their interactions with ACE2. Cells 2020, 9, 2638. [Google Scholar] [CrossRef]
- Masters, P.S. Coronavirus genomic RNA packaging. Virology 2019, 537, 198–207. [Google Scholar] [CrossRef]
- Berlin, D.A.; Gulick, R.M.; Martinez, F.J. Severe covid-19. N. Engl. J. Med. 2020, 383, 2451–2460. [Google Scholar] [CrossRef]
- Merad, M.; Martin, J.C. Pathological inflammation in patients with COVID-19: A key role for monocytes and macrophages. Nat. Rev. Immunol. 2020, 20, 355–362. [Google Scholar] [CrossRef]
- Machhi, J.; Herskovitz, J.; Senan, A.M.; Dutta, D.; Nath, B.; Oleynikov, M.D.; Blomberg, W.R.; Meigs, D.D.; Hasan, M.; Patel, M. The natural history, pathobiology, and clinical manifestations of SARS-CoV-2 infections. J. Neuroimmune Pharmacol. 2020, 15, 359–386. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020, 181, 281–292.e286. [Google Scholar] [CrossRef]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef]
- Wang, K.; Chen, W.; Zhang, Z.; Deng, Y.; Lian, J.Q.; Du, P.; Wei, D.; Zhang, Y.; Sun, X.X.; Gong, L.; et al. CD147-spike protein is a novel route for SARS-CoV-2 infection to host cells. Signal Transduct. Target. Ther. 2020, 5, 283. [Google Scholar] [CrossRef]
- Ulrich, H.; Pillat, M.M. CD147 as a target for COVID-19 treatment: Suggested effects of azithromycin and stem cell engagement. Stem Cell Rev. Rep. 2020, 16, 434–440. [Google Scholar] [CrossRef]
- da Silva, G.B.; Manica, D.; da Silva, A.P.; Kosvoski, G.C.; Hanauer, M.; Assmann, C.E.; Simões, J.L.B.; Pillat, M.M.; de Lara, J.D.; Marafon, F.; et al. High levels of extracellular ATP lead to different inflammatory responses in COVID-19 patients according to the severity. J. Mol. Med. 2022, 100, 645–663. [Google Scholar] [CrossRef]
- Romão, P.R.; Teixeira, P.C.; Schipper, L.; da Silva, I.; Santana Filho, P.; Júnior, L.C.R.; Peres, A.; Gonçalves da Fonseca, S.; Chagas Monteiro, M.; Lira, F.S.; et al. Viral load is associated with mitochondrial dysfunction and altered monocyte phenotype in acute severe SARS-CoV-2 infection. Int. Immunopharmacol. 2022, 108, 108697. [Google Scholar] [CrossRef]
- Kas-Deelen, A.; Bakker, W.; Olinga, P.; Visser, J.; De Maar, E.; Van Son, W.; The, T.; Harmsen, M.J. Cytomegalovirus infection increases the expression and activity of ecto-ATPase (CD39) and ecto-5′ nucleotidase (CD73) on endothelial cells. FEBS Lett. 2001, 491, 21–25. [Google Scholar] [CrossRef]
- Schwele, S.; Fischer, A.M.; Brestrich, G.; Wlodarski, M.W.; Wagner, L.; Schmueck, M.; Roemhild, A.; Thomas, S.; Hammer, M.H.; Babel, N.; et al. Cytomegalovirus-specific regulatory and effector T cells share TCR clonality--possible relation to repetitive CMV infections. Am. J. Transpl. 2012, 12, 669–681. [Google Scholar] [CrossRef]
- De Rossi, A.; Calabro, M.L.; D’Andrea, E.; Panozzo, M.; Saggiori, D.; Mammano, F.; Roncella, S.; Ferrarini, M.; Chieco-Bianchi, L. Morphological and phenotypical changes in EBV positive lymphoblastoid cells infected by HIV-1. Leukemia 1992, 6 (Suppl. S3), 38s–40s. [Google Scholar]
- Falk, M.H.; Trauth, B.C.; Debatin, K.M.; Klas, C.; Gregory, C.D.; Rickinson, A.B.; Calender, A.; Lenoir, G.M.; Ellwart, J.W.; Krammer, P.H.; et al. Expression of the APO-1 antigen in Burkitt lymphoma cell lines correlates with a shift towards a lymphoblastoid phenotype. Blood 1992, 79, 3300–3306. [Google Scholar] [CrossRef] [PubMed]
- Auclair, H.; Ouk-Martin, C.; Roland, L.; Santa, P.; Al Mohamad, H.; Faumont, N.; Feuillard, J.; Jayat-Vignoles, C. EBV Latency III-Transformed B Cells Are Inducers of Conventional and Unconventional Regulatory T Cells in a PD-L1-Dependent Manner. J. Immunol. 2019, 203, 1665–1674. [Google Scholar] [CrossRef] [PubMed]
- Tang, A.; Dadaglio, G.; Oberkampf, M.; Di Carlo, S.; Peduto, L.; Laubreton, D.; Desrues, B.; Sun, C.M.; Montagutelli, X.; Leclerc, C. B cells promote tumor progression in a mouse model of HPV-mediated cervical cancer. Int. J. Cancer 2016, 139, 1358–1371. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Hoya, A.; Zerecero-Carreón, O.; Valle-Mendiola, A.; Moreno-Lafont, M.; López-Santiago, R.; Weiss-Steider, B.; Soto-Cruz, I. Cervical Cancer Cells Express Markers Associated with Immunosurveillance. J. Immunol. Res. 2019, 2019, 1242979. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.N.; Zhang, N.; Liu, H.H.; Xia, P.; Zhang, C.; Song, J.W.; Fan, X.; Shi, M.; Jin, L.; Zhang, J.Y.; et al. Skewed CD39/CD73/adenosine pathway contributes to B-cell hyperactivation and disease progression in patients with chronic hepatitis B. Gastroenterol. Rep. 2021, 9, 49–58. [Google Scholar] [CrossRef]
- Schottstedt, V.; Blümel, J.; Burger, R.; Drosten, C.; Gröner, A.; Gürtler, L.; Heiden, M.; Hildebrandt, M.; Jansen, B.; Montag-Lessing, T.; et al. Human Cytomegalovirus (HCMV)—Revised. Transfus. Med. Hemother. 2010, 37, 365–375. [Google Scholar] [CrossRef]
- Hosie, L.; Pachnio, A.; Zuo, J.; Pearce, H.; Riddell, S.; Moss, P. Cytomegalovirus-Specific T Cells Restricted by HLA-Cw*0702 Increase Markedly with Age and Dominate the CD8(+) T-Cell Repertoire in Older People. Front. Immunol. 2017, 8, 1776. [Google Scholar] [CrossRef]
- Soldan, S.S.; Lieberman, P.M. Epstein–Barr virus and multiple sclerosis. Nat. Rev. Microbiol. 2022. [Google Scholar] [CrossRef]
- Thorley-Lawson, D.A. EBV Persistence--Introducing the Virus. Curr. Top. Microbiol. Immunol. 2015, 390, 151–209. [Google Scholar] [CrossRef]
- Young, L.S.; Yap, L.F.; Murray, P.G. Epstein-Barr virus: More than 50 years old and still providing surprises. Nat. Rev. Cancer 2016, 16, 789–802. [Google Scholar] [CrossRef]
- Nie, K.; Zheng, G.G.; Zhang, X.J.; Lin, Y.M.; Wang, L.; Li, G.; Song, Y.H.; Wu, K.F. CD 39-associated high ATPase activity contribute to the loss of P 2 X 7-mediated calcium response in LCL cells. Leuk. Res. 2005, 29, 1325–1333. [Google Scholar] [CrossRef]
- Simoni, Y.; Becht, E.; Li, S.; Loh, C.Y.; Yeong, J.P.S.; Lim, T.K.H.; Takano, A.; Tan, D.S.W.; Newell, E.W. Partial absence of PD-1 expression by tumor-infiltrating EBV-specific CD8(+) T cells in EBV-driven lymphoepithelioma-like carcinoma. Clin. Transl. Immunol. 2020, 9, e1175. [Google Scholar] [CrossRef]
- Brianti, P.; De Flammineis, E.; Mercuri, S.R. Review of HPV-related diseases and cancers. New Microbiol. 2017, 40, 80–85. [Google Scholar]
- zur Hausen, H. Papillomaviruses in the causation of human cancers—A brief historical account. Virology 2009, 384, 260–265. [Google Scholar] [CrossRef] [Green Version]
- Stoler, M.H.; Rhodes, C.R.; Whitbeck, A.; Wolinsky, S.M.; Chow, L.T.; Broker, T.R. Human papillomavirus type 16 and 18 gene expression in cervical neoplasias. Hum. Pathol. 1992, 23, 117–128. [Google Scholar] [CrossRef]
- de Lourdes Mora-García, M.; López-Cisneros, S.; Gutiérrez-Serrano, V.; García-Rocha, R.; Weiss-Steider, B.; Hernández-Montes, J.; Sánchez-Peña, H.I.; Ávila-Ibarra, L.R.; Don-López, C.A.; Muñóz-Godínez, R.; et al. HPV-16 Infection Is Associated with a High Content of CD39 and CD73 Ectonucleotidases in Cervical Samples from Patients with CIN-1. Mediat. Inflamm. 2019, 2019, 4651627. [Google Scholar] [CrossRef]
- Duurland, C.L.; Santegoets, S.J.; Abdulrahman, Z.; Loof, N.M.; Sturm, G.; Wesselink, T.H.; Arens, R.; Boekestijn, S.; Ehsan, I.; van Poelgeest, M.I.E.; et al. CD161 expression and regulation defines rapidly responding effector CD4+ T cells associated with improved survival in HPV16-associated tumors. J. Immunother. Cancer 2022, 10, e003995. [Google Scholar] [CrossRef]
- Schaefer, S. Hepatitis B virus taxonomy and hepatitis B virus genotypes. World J. Gastroenterol. 2007, 13, 14–21. [Google Scholar] [CrossRef]
- Liang, T.J. Hepatitis B: The virus and disease. Hepatology 2009, 49 (Suppl. S5), S13–S21. [Google Scholar] [CrossRef]
- Ravimohan, S.; Tamuhla, N.; Nfanyana, K.; Ni, H.; Steenhoff, A.P.; Gross, R.; Weissman, D.; Bisson, G.P. Elevated Pre-Antiretroviral Therapy CD39+CD8+ T Cell Frequency Is Associated with Early Mortality in Advanced Human Immunodeficiency Virus/Tuberculosis Co-infection. Clin. Infect. Dis. 2017, 64, 1453–1456. [Google Scholar] [CrossRef]
- Angerami, M.T.; Suarez, G.V.; Vecchione, M.B.; Laufer, N.; Ameri, D.; Ben, G.; Perez, H.; Sued, O.; Salomón, H.; Quiroga, M.F. Expansion of CD25-Negative Forkhead Box P3-Positive T Cells during HIV and Mycobacterium tuberculosis Infection. Front. Immunol. 2017, 8, 528. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Virus | Family | Virology | Main Findings Related to CD39+ Cells | References |
|---|---|---|---|---|
| HIV | Retroviridae |
|
| [21,38] |
| [39] | |||
| [40] | |||
| [41] | |||
| [42] | |||
| HCV | Flaviviridae |
|
| [43,44] |
| [21] | |||
| [44] | |||
| [45] | |||
| HEV | Hepeviridae |
|
| [46] |
| [46] | |||
| HDV | The Deltaviridae genus does not belong to a known family |
|
| [47] |
| Influenza | Orthomyxoviridae |
|
| [48,49] |
| Dengue virus | Flaviviridae |
|
| [50] |
| Zika virus | Flaviviridae |
|
| [51] |
| Severe Acute Respiratory Syndrome Corona Virus-2 (SARS-CoV-2) | Coronaviridae |
|
| [52] |
| [53] | |||
| [54] | |||
| [55] |
| Virus | Family | Virology | Main Findings Related to CD39+ Cells | References |
|---|---|---|---|---|
| CMV | Herpesviridae |
| ↑ expression of CD39 on endothelial cells → regulates the platelet function and inflammation | [119] |
| ↑ expression on CD25+ Treg cells → recurrence and/or latent infection | [120] | |||
| EBV | Herpesviridae |
| Expressed on EBV-infected cancer B cells, Burkitt lymphoma cells, and LCL | [3,121,122] |
| EBV ↑ CD39+ Treg cells | [123] | |||
| HPV | Papillomaviridae |
| ↑ expression of CD39 on the B cells inside the tumor | [124] |
| ↑ expression of CD39 and CD73 on cervical cancer cells infected with HPV → immune escape | [125] | |||
| HBV | Hepadnaviridae |
| A lower percentage of CD39+ Treg cells in HBV-infected patients than in healthy controls | [29] |
| The expression CD39/CD73 in B cells is negatively associated with liver inflammation | [126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsaghir, A.; El-Sabaa, E.M.W.; Ahmed, A.K.; Abdelwahab, S.F.; Sayed, I.M.; El-Mokhtar, M.A. The Role of Cluster of Differentiation 39 (CD39) and Purinergic Signaling Pathway in Viral Infections. Pathogens 2023, 12, 279. https://doi.org/10.3390/pathogens12020279
Elsaghir A, El-Sabaa EMW, Ahmed AK, Abdelwahab SF, Sayed IM, El-Mokhtar MA. The Role of Cluster of Differentiation 39 (CD39) and Purinergic Signaling Pathway in Viral Infections. Pathogens. 2023; 12(2):279. https://doi.org/10.3390/pathogens12020279
Chicago/Turabian StyleElsaghir, Alaa, Ehsan M. W. El-Sabaa, Abdulrahman K. Ahmed, Sayed F. Abdelwahab, Ibrahim M. Sayed, and Mohamed A. El-Mokhtar. 2023. "The Role of Cluster of Differentiation 39 (CD39) and Purinergic Signaling Pathway in Viral Infections" Pathogens 12, no. 2: 279. https://doi.org/10.3390/pathogens12020279