
Identification of Pathogenic Leptospira kirschneri Serogroup Grippotyphosa in Water Voles (Arvicola terrestris) from Ruminant Pastures in Puy-de-Dôme, Central France
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement on Vole Sampling
2.2. Provision of Sheep Blood Samples
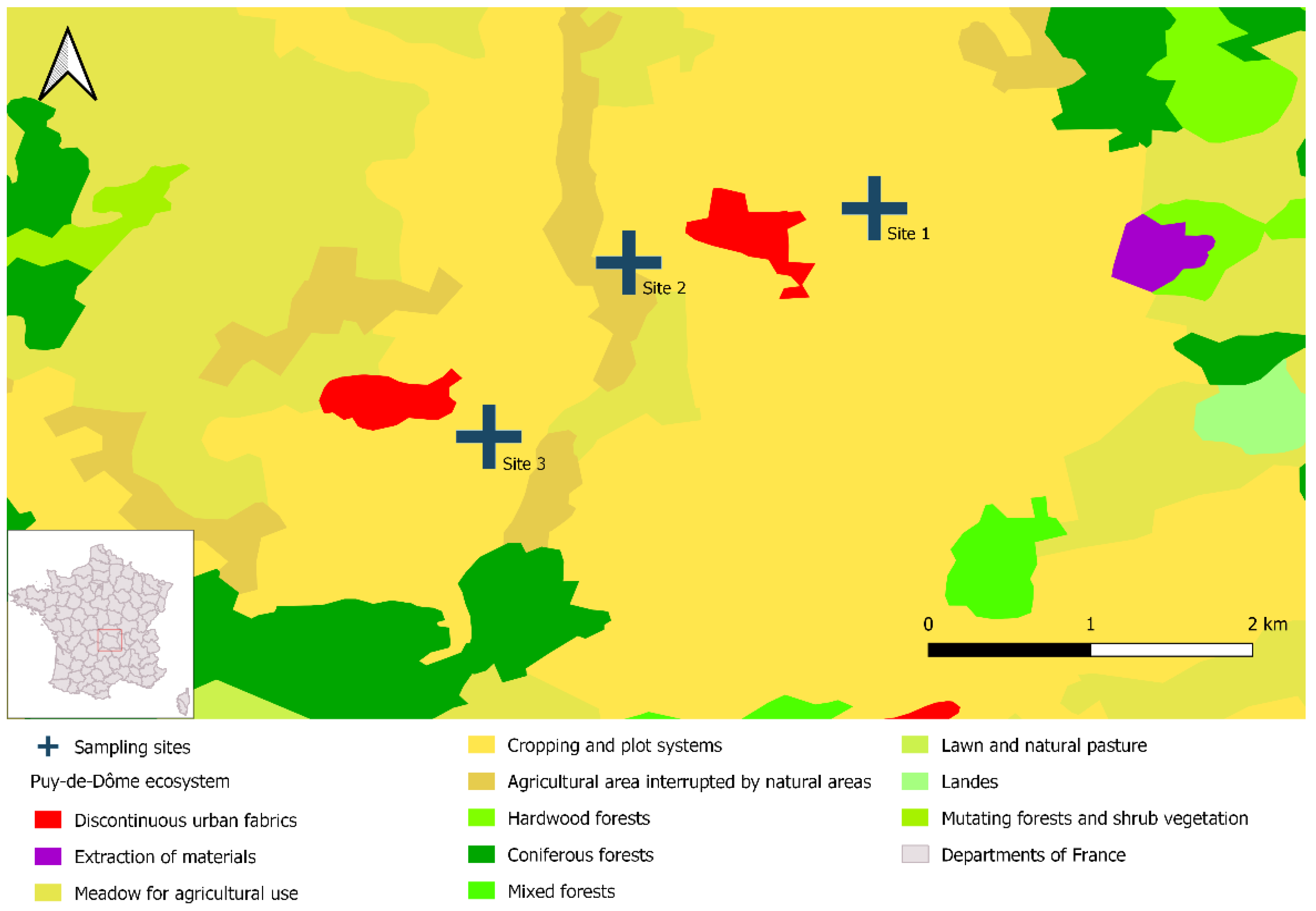
2.3. Sampling Sites
2.4. Vole Trapping and Sampling
2.5. Sheep Sampling
2.6. Culturing Leptospires from Urine and Renal Tissue
2.7. Extraction and Detection of Leptospira DNA
2.8. Genetic Characterization of Leptospira
2.8.1. Conventional PCR Targeting the 16S rDNA Gene
2.8.2. Molecular Typing of Leptospira DNA Based on O-Antigen and Variable Number Tandem Repeat (VNTR) Methods
2.9. Microagglutination Testing
2.10. Data Analysis
3. Results
3.1. Vole Characteristics
3.2. Vole Infection Status
3.2.1. Cultures
3.2.2. Characteristics of Infected Voles
3.2.3. Detecting Leptospira Infection in Voles
3.2.4. Molecular Typing of Leptospira Strains in Voles
3.3. Leptospira Seroprevalence in Sheep
Microagglutination Testing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levett, P.N. Leptospirosis. Clin. Microbiol. Rev. 2001, 14, 296–326. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef]
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E.; et al. Leptospirosis: A Zoonotic Disease of Global Importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Zaki, S.R.; Shieh, W.-J.; Epidemic Working Group at Ministry of Health in Nicaragua; Pan American Health Organisation; US Department of Agriculture; Centers for Disease Control and Prevention. Leptospirosis Associated with Outbreak of Acute Febrile Illness and Pulmonary Haemorrhage, Nicaragua, 1995. Lancet 1996, 347, 535–536. [Google Scholar] [CrossRef] [PubMed]
- Ellis, W.A. Animal Leptospirosis. Curr. Top. Microbiol. Immunol. 2015, 387, 99–137. [Google Scholar]
- Boey, K.; Shiokawa, K.; Rajeev, S. Leptospira Infection in Rats: A Literature Review of Global Prevalence and Distribution. PLoS Negl. Trop. Dis. 2019, 13, e0007499. [Google Scholar] [CrossRef] [PubMed]
- Meerburg, B.G.; Singleton, G.R.; Kijlstra, A. Rodent-Borne Diseases and Their Risks for Public Health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef]
- Yatbantoong, N.; Chaiyarat, R. Factors Associated with Leptospirosis in Domestic Cattle in Salakphra Wildlife Sanctuary, Thailand. Int. J. Environ. Res. Public Health 2019, 16, 1042. [Google Scholar] [CrossRef]
- Joshi, Y.P.; Kim, E.-H.; Cheong, H.-K. The Influence of Climatic Factors on the Development of Hemorrhagic Fever with Renal Syndrome and Leptospirosis during the Peak Season in Korea: An Ecologic Study. BMC Infect. Dis. 2017, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.L.; Smythe, L.D.; Craig, S.B.; Weinstein, P. Climate Change, Flooding, Urbanisation and Leptospirosis: Fuelling the Fire? Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Bourhy, P.; Picardeau, M.; Septfons, A.; Trombert, S.; Cart-tanneur, E. Émergence de La Leptospirose Humaine En France Métropolitaine? Actualités Sur La Surveillance. Médecine Mal. Infect. 2017, 47, S150. [Google Scholar] [CrossRef]
- Perra, A.; Servas, V.; Terrier, G.; Postic, D.; Baranton, G.; André-Fontaine, G.; Vaillant, V.; Capek, I. Clustered Cases of Leptospirosis in Rochefort, France, June 2001. Eurosurveill 2002, 7, 131–136. [Google Scholar] [CrossRef]
- Nardone, A.; Capek, I.; Baranton, G.; Campèse, C.; Postic, D.; Vaillant, V.; Liénard, M.; Desenclos, J.C. Risk Factors for Leptospirosis in Metropolitan France: Results of a National Case-Control Study, 1999-2000. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2004, 39, 423272. [Google Scholar] [CrossRef] [PubMed]
- Guillois, Y.; Bourhy, P.; Ayral, F.; Pivette, M.; Decors, A.; Aranda Grau, J.H.; Champenois, B.; Malhère, C.; Combes, B.; Richomme, C.; et al. An Outbreak of Leptospirosis among Kayakers in Brittany, North-West France, 2016. Eurosurveill 2018, 23, 1700848. [Google Scholar] [CrossRef] [PubMed]
- Arent, Z.; Pardyak, L.; Dubniewicz, K.; Plachno, B.; Kotula-Balak, M. Leptospira Taxonomy: Then and Now. Med. Weter 2022, 78, 489–496. [Google Scholar] [CrossRef]
- Picardeau, M. Virulence of the Zoonotic Agent of Leptospirosis: Still Terra Incognita? Nat. Rev. Microbiol. 2017, 15, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Philip, N.; Affendy, N.B.; Ramli, S.N.A.; Arif, M.; Raja, P.; Nagandran, E.; Renganathan, P.; Taib, N.M.; Masri, S.N.; Yuhana, M.Y.; et al. Leptospira Interrogans and Leptospira Kirschneri Are the Dominant Leptospira Species Causing Human Leptospirosis in Central Malaysia. PLoS Negl. Trop. Dis. 2020, 14, 8197. [Google Scholar] [CrossRef]
- Kmety, E.; Dikken, H. Classification of the Species Leptospira Interrogans and History of Its Serovars; University Press: Groningen, The Nethersland, 1993. [Google Scholar]
- Tsujimoto, M.; Kajikawa, Y.; Tomita, J.; Matsumoto, Y. A Review of the Ecosystem Concept—Towards Coherent Ecosystem Design. Technol. Forecast. Soc. Change 2018, 136, 49–58. [Google Scholar] [CrossRef]
- Roberts, M.G.; Heesterbeek, J.A.P. Characterizing Reservoirs of Infection and the Maintenance of Pathogens in Ecosystems. J. R. Soc. Interface 2020, 17, 20190540. [Google Scholar] [CrossRef] [PubMed]
- Viana, M.; Mancy, R.; Biek, R.; Cleaveland, S.; Cross, P.C.; Lloyd-Smith, J.O.; Haydon, D.T. Assembling Evidence for Identifying Reservoirs of Infection. Trends Ecol. Evol. 2014, 29, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Thiermann, A.B. The Norway Rat as a Selective Chronic Carrier of Leptospira Icterohaemorrhagiae. J. Wildl. Dis. 1981, 17, 39–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haake, D.A.; Levett, P.N. Leptospirosis in Humans. Curr. Top. Microbiol. Immunol. 2015, 387, 65–97. [Google Scholar] [PubMed]
- Thiermann, A.B. Experimental Leptospiral Infections in Pregnant Cattle with Organisms of the Hebdomadis Serogroup. Am. J. Vet. Res. 1982, 43, 780–784. [Google Scholar]
- Schuller, S.; Francey, T.; Hartmann, K.; Hugonnard, M.; Kohn, B.; Nally, J.E.; Sykes, J. European Consensus Statement on Leptospirosis in Dogs and Cats. J. Small Anim. Pract. 2015, 56, 159–179. [Google Scholar] [CrossRef]
- Ellis, W.A.; Straw, B.E.; D’Allaire, S.; Mengeling, W.L.; Taylor, D.J. Diseases of Swine; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Bertasio, C.; Papetti, A.; Scaltriti, E.; Tagliabue, S.; D’Incau, M.; Boniotti, M.B. Serological Survey and Molecular Typing Reveal New Leptospira Serogroup Pomona Strains among Pigs of Northern Italy. Pathogens 2020, 9, 332. [Google Scholar] [CrossRef]
- Cilia, G.; Bertelloni, F.; Piredda, I.; Ponti, M.N.; Turchi, B.; Cantile, C.; Parisi, F.; Pinzauti, P.; Armani, A.; Palmas, B. Presence of Pathogenic Leptospira Spp. in the Reproductive System and Fetuses of Wild Boars (Sus Scrofa) in Italy. PLoS Negl. Trop. Dis. 2020, 14, e0008982. [Google Scholar] [CrossRef]
- Jacquot, M.; Abrial, D.; Gasqui, P.; Bord, S.; Marsot, M.; Masseglia, S.; Pion, A.; Poux, V.; Zilliox, L.; Chapuis, J.-L.; et al. Multiple Independent Transmission Cycles of a Tick-Borne Pathogen within a Local Host Community. Sci. Rep. 2016, 6, 31273. [Google Scholar] [CrossRef] [PubMed]
- Newbold, T.; Hudson, L.N.; Arnell, A.P.; Contu, S.; De Palma, A.; Ferrier, S.; Hill, S.L.; Hoskins, A.J.; Lysenko, I.; Phillips, H.R. Has Land Use Pushed Terrestrial Biodiversity beyond the Planetary Boundary? A Global Assessment. Science 2016, 353, 288–291. [Google Scholar] [CrossRef]
- Fichet-Calvet, E.; Pradier, B.; Quéré, J.-P.; Giraudoux, P.; Delattre, P. Landscape Composition and Vole Outbreaks: Evidence from an Eight Year Study of Arvicola Terrestris. Ecography 2000, 23, 659–668. [Google Scholar] [CrossRef]
- Delattre, P.; Giraudoux, P.; Baudry, J.; Musard, P.; Toussaint, M.; Truchetet, D.; Stahl, P.; Poule, M.L.; Artois, M.; Damange, J.-P.; et al. Land Use Patterns and Types of Common Vole (Microtus Arvalis) Population Kinetics. Agric. Ecosyst. Environ. 1992, 39, 153–168. [Google Scholar] [CrossRef]
- Giraudoux, P.; Delattre, P.; Habert, M.; Quéré, J.P.; Deblay, S.; Defaut, R.; Duhamel, R.; Moissenet, M.F.; Salvi, D.; Truchetet, D. Population Dynamics of Fossorial Water Vole (Arvicola Terrestris Scherman): A Land Use and Landscape Perspective. Agric. Ecosyst. Environ. 1997, 66, 47–60. [Google Scholar] [CrossRef]
- Johnsen, K.; Devineau, O.; Andreassen, H.P. Phase- and Season-Dependent Changes in Social Behaviour in Cyclic Vole Populations. BMC Ecol. 2019, 19, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer-Scholl, A.; Hammerl, J.A.; Schmidt, S.; Ulrich, R.G.; Pfeffer, M.; Woll, D.; Scholz, H.C.; Thomas, A.; Nöckler, K. Leptospira spp. in Rodents and Shrews in Germany. Int. J. Environ. Res. Public Health 2014, 11, 7562–7574. [Google Scholar] [CrossRef]
- Jeske, K.; Emirhar, D.; García, J.T.; González-Barrio, D.; Olea, P.P.; Fons, F.R.; Schulz, J.; Mayer-Scholl, A.; Heckel, G.; Ulrich, R.G. Frequent Leptospira spp. Detection but Absence of Tula Orthohantavirus in Microtus Spp. Voles, Northwestern Spain. J. Wildl. Dis. 2021, 57, 733–742. [Google Scholar] [CrossRef]
- Fischer, S.; Mayer-Scholl, A.; Imholt, C.; Spierling, N.G.; Heuser, E.; Schmidt, S.; Reil, D.; Rosenfeld, U.M.; Jacob, J.; Nöckler, K. Leptospira Genomospecies and Sequence Type Prevalence in Small Mammal Populations in Germany. Vector-Borne Zoonotic Dis. 2018, 18, 188–199. [Google Scholar] [CrossRef]
- Obiegala, A.; Albrecht, C.; Dafalla, M.; Drewes, S.; Oltersdorf, C.; Turni, H.; Imholt, C.; Jacob, J.; Wagner-Wiening, C.; Ulrich, R.G. Leptospira spp. in Small Mammals from Areas with Low and High Human Hantavirus Incidences in South-West Germany. Vector-Borne Zoonotic Dis. 2017, 17, 312–318. [Google Scholar] [CrossRef]
- Villette, P.; Afonso, E.; Couval, G.; Levret, A.; Galan, M.; Tatard, C.; Cosson, J.F.; Giraudoux, P. Consequences of Organ Choice in Describing Bacterial Pathogen Assemblages in a Rodent Population. Epidemiol. Infect. 2017, 145, 3070–3075. [Google Scholar] [CrossRef]
- Marquez, A. Gestion de Populations de Rongeurs Dans Un Contexte Leptospirosique. PhD Thesis, Université de Lyon, Lyon, France, 2019. [Google Scholar]
- Auzias, D.; Labourdette, J.-P. AUVERGNE. Petit Futé: Paris, France; Volume 2017, ISBN 979-10-331-5942-1.
- Meillour, N.-L.; Descamps, A.; Le Danvic, C.; Grandmougin, M.; Saliou, J.-M.; Klopp, C.; Milhes, M.; Bompard, C.; Chesneau, D.; Poissenot, K. Identification of Potential Chemosignals in the European Water Vole Arvicola Terrestris. Sci. Rep. 2019, 9, 18378. [Google Scholar] [CrossRef]
- Truchetet, D.; Couval, G.; Michelin, Y.; Giraudoux, P. Exploring the Factors That Affect Water Vole (Arvicola Terrestris) Outbreaks in Grasslands. Fourrages 2014, 220, 279–284. [Google Scholar]
- Zilber, A.-L.; Belli, P.; Artois, M.; Kodjo, A.; Djelouadji, Z. First Observation of Leptospira Interrogans in the Lungs of Rattus Norvegicus. BioMed Res. Int. 2016, 2016, 9656274. [Google Scholar] [CrossRef] [PubMed]
- World Organization of Animal Health—OIE. Chapter 3.1.12. Leptospirosis. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organization of Animal Health: Paris, France, 2021. [Google Scholar]
- Chakraborty, A.; Miyahara, S.; Villanueva, S.Y.; Saito, M.; Gloriani, N.G.; Yoshida, S. A Novel Combination of Selective Agents for Isolation of Leptospira Species. Microbiol. Immunol. 2011, 55, 494–501. [Google Scholar] [CrossRef]
- Picardeau, M. Diagnosis and Epidemiology of Leptospirosis. Med. Mal. Infect. 2013, 43, 1–9. [Google Scholar] [CrossRef]
- Toussaint, J.F.; Sailleau, C.; Breard, E.; Zientara, S.; De Clercq, K. Bluetongue Virus Detection by Two Real-Time RT-QPCRs Targeting Two Different Genomic Segments. J. Virol. Methods 2007, 140, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Waggoner, J.J.; Balassiano, I.; Abeynayake, J.; Sahoo, M.K.; Mohamed-Hadley, A.; Liu, Y.; Vital-Brazil, J.M.; Pinsky, B.A. Sensitive Real-Time PCR Detection of Pathogenic Leptospira Spp. and a Comparison of Nucleic Acid Amplification Methods for the Diagnosis of Leptospirosis. PLoS ONE 2014, 9, e112356. [Google Scholar] [CrossRef]
- Merien, F.; Amouriaux, P.; Perolat, P.; Baranton, G.; Saint Girons, I. Polymerase Chain Reaction for Detection of Leptospira spp. in Clinical Samples. J. Clin. Microbiol. 1992, 30, 2219–2224. [Google Scholar] [CrossRef]
- Cai, C.-S.; Zhu, Y.-Z.; Zhong, Y.; Xin, X.-F.; Jiang, X.-G.; Lou, X.-L.; He, P.; Qin, J.-H.; Zhao, G.-P.; Wang, S.-Y.; et al. Development of O-Antigen Gene Cluster-Specific PCRs for Rapid Typing Six Epidemic Serogroups of Leptospira in China. BMC Microbiol. 2010, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Salaün, L.; Mérien, F.; Gurianova, S.; Baranton, G.; Picardeau, M. Application of Multilocus Variable-Number Tandem-Repeat Analysis for Molecular Typing of the Agent of Leptospirosis. J. Clin. Microbiol. 2006, 44, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.K.; Gray, M.R.; Mende, K.; Parker, T.M.; Samir, A.; Rahman, B.A.; Habashy, E.E.; Hospenthal, D.R.; Pimentel, G. Use of Patient-Specific Leptospira Isolates in the Diagnosis of Leptospirosis Employing Microscopic Agglutination Testing (MAT). Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 209–213. [Google Scholar] [CrossRef]
- Scarcelli, E.; Piatti, R.M.; Fedullo, J.D.L.; Simon, F.; Cardoso, M.V.; Castro, V.; Miyashiro, S.; Genovez, M.É. Leptospira spp. Detection by Polymerase Chain Reaction (PCR) in Clinical Samples of Captive Black-Capped Capuchin Monkey (Cebus Apella). Braz. J. Microbiol. 2003, 34, 143–146. [Google Scholar] [CrossRef]
- Agudelo-Flórez, P.; Murillo, V.E.; Londoño, A.F.; Rodas, J.D. Histopathological Kidney Alterations in Rats Naturally Infected with Leptospira. Biomédica 2013, 33, 82–88. [Google Scholar]
- Hartskeerl, R.A.; Terpstra, W.J. Leptospirosis in Wild Animals. Vet. Q. 1996, 18 (Suppl. S3), S149–S150. [Google Scholar] [CrossRef]
- AYRAL, F.; ARTOIS, J.; ZILBER, A.-L.; WIDÉN, F.; POUNDER, K.C.; AUBERT, D.; BICOUT, D.J.; ARTOIS, M. The Relationship between Socioeconomic Indices and Potentially Zoonotic Pathogens Carried by Wild Norway Rats: A Survey in Rhône, France (2010–2012). Epidemiol. Infect. 2015, 143, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Romanazzi, V.; Traversi, D.; Lorenzi, E.; Gilli, G. Effects of Freezing Storage on the DNA Extraction and Microbial Evaluation from Anaerobic Digested Sludges. BMC Res. Notes 2015, 8, 420. [Google Scholar] [CrossRef]
- Bahl, M.I.; Bergström, A.; Licht, T.R. Freezing Fecal Samples Prior to DNA Extraction Affects the Firmicutes to Bacteroidetes Ratio Determined by Downstream Quantitative PCR Analysis. FEMS Microbiol. Lett. 2012, 329, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Kwambana, B.A.; Mohammed, N.I.; Jeffries, D.; Barer, M.; Adegbola, R.A.; Antonio, M. Differential Effects of Frozen Storage on the Molecular Detection of Bacterial Taxa That Inhabit the Nasopharynx. BMC Clin. Pathol. 2011, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Lawlor, P.G.; Magowan, E.; Zebeli, Q. Effect of Freezing Conditions on Fecal Bacterial Composition in Pigs. Animals 2016, 6, 18. [Google Scholar] [CrossRef]
- Cameron, C.E.; Zuerner, R.L.; Raverty, S.; Colegrove, K.M.; Norman, S.A.; Lambourn, D.M.; Jeffries, S.J.; Gulland, F.M. Detection of Pathogenic Leptospira Bacteria in Pinniped Populations via PCR and Identification of a Source of Transmission for Zoonotic Leptospirosis in the Marine Environment. J. Clin. Microbiol. 2008, 46, 1728–1733. [Google Scholar] [CrossRef]
- Lucchesi, P.; Arroyo, G.H.; Etcheverría, A.I.; Parma, A.E.; Seijo, A.C. Recommendations for the Detection of Leptospira in Urine by PCR. Rev. Da Soc. Bras. De Med. Trop. 2004, 37, 131–134. [Google Scholar] [CrossRef]
- Shao, W.; Khin, S.; Kopp, W.C. Characterization of Effect of Repeated Freeze and Thaw Cycles on Stability of Genomic DNA Using Pulsed Field Gel Electrophoresis. Biopreserv. Biobank. 2012, 10, 4–11. [Google Scholar] [CrossRef]
- Easterbrook, J.D.; Kaplan, J.B.; Vanasco, N.B.; Reeves, W.K.; Purcell, R.H.; Kosoy, M.Y.; Glass, G.E.; Watson, J.; Klein, S.L. A Survey of Zoonotic Pathogens Carried by Norway Rats in Baltimore, Maryland, USA. Epidemiol. Infect. 2007, 135, 1192–1199. [Google Scholar] [CrossRef]
- Costa, F.; Porter, F.H.; Rodrigues, G.; Farias, H.; de Faria, M.T.; Wunder, E.A.; Osikowicz, L.M.; Kosoy, M.Y.; Reis, M.G.; Ko, A.I.; et al. Infections by Leptospira Interrogans, Seoul Virus, and Bartonella Spp. Among Norway Rats (Rattus Norvegicus) from the Urban Slum Environment in Brazil. Vector Borne Zoonotic Dis. 2014, 14, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Krijger, I.M.; Ahmed, A.A.A.; Goris, M.G.A.; Groot Koerkamp, P.W.G.; Meerburg, B.G. Prevalence of Leptospira Infection in Rodents from Bangladesh. Int. J. Environ. Res. Public Health 2019, 16, 2113. [Google Scholar] [CrossRef] [PubMed]
- Minter, A.; Diggle, P.J.; Costa, F.; Childs, J.; Ko, A.I.; Begon, M. Evidence of Multiple Intraspecific Transmission Routes for Leptospira Acquisition in Norway Rats (Rattus Norvegicus). Epidemiol. Infect. 2017, 145, 3438–3448. [Google Scholar] [CrossRef] [PubMed]
- Tucunduva de Faria, M.; Athanazio, D.A.; Gonçalves Ramos, E.A.; Silva, E.F.; Reis, M.G.; Ko, A.I. Morphological Alterations in the Kidney of Rats with Natural and Experimental Leptospira Infection. J. Comp. Pathol. 2007, 137, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.G.; Allen, J.E.; Wilson, R.B. The Pathogenesis of Hemorrhage in the Lung of the Hamster during Acute Leptospirosis. Med. Microbiol. Immunol. 1974, 160, 269–278. [Google Scholar] [CrossRef]
- Athanazio, D.A.; Silva, E.F.; Santos, C.S.; Rocha, G.M.; Vannier-Santos, M.A.; McBride, A.J.; Ko, A.I.; Reis, M.G. Rattus Norvegicus as a Model for Persistent Renal Colonization by Pathogenic Leptospira Interrogans. Acta Trop. 2008, 105, 176–180. [Google Scholar] [CrossRef]
- Mori, M.; Bourhy, P.; Le Guyader, M.; Van Esbroeck, M.; Djelouadji, Z.; Septfons, A.; Kodjo, A.; Picardeau, M. Pet Rodents as Possible Risk for Leptospirosis, Belgium and France, 2009 to 2016. Eurosurveillance 2017, 22, 16–00792. [Google Scholar] [CrossRef] [PubMed]
- Goy-Thollot, I.; Djelouadji, Z.; Nennig, M.; Hazart, G.; Hugonnard, M. Screening for Leptospira DNA in Blood and Urine from 30 Apparently Healthy Dogs. Rev. Vet. Clin. 2018, 53, 79–86. [Google Scholar] [CrossRef]
- Benacer, D.; Zain, S.N.M.; Amran, F.; Galloway, R.L.; Thong, K.L. Isolation and Molecular Characterization of Leptospira Interrogans and Leptospira Borgpetersenii Isolates from the Urban Rat Populations of Kuala Lumpur, Malaysia. Am. J. Trop. Med. Hyg. 2013, 88, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Public Health Ontario. An Overview of Cycle Threshold Values and Their Role in SARS-CoV-2 Real-Time PCR Test Interpretation; Queen’s Printer for Ontario: Toronto, ON, Canada, 2020; pp. 1–14.
- Waggoner, J.J.; Balassiano, I.; Mohamed-Hadley, A.; Vital-Brazil, J.M.; Sahoo, M.K.; Pinsky, B.A. Reverse-Transcriptase PCR Detection of Leptospira: Absence of Agreement with Single-Specimen Microscopic Agglutination Testing. PLoS ONE 2015, 10, e0132988. [Google Scholar] [CrossRef]
- Wagenaar, J.; Zuerner, R.L.; Alt, D.; Bolin, C.A. Comparison of Polymerase Chain Reaction Assays with Bacteriologic Culture, Immunofluorescence, and Nucleic Acid Hybridization for Detection of Leptospira Borgpetersenii Serovar Hardjo in Urine of Cattle. Am. J. Vet. Res. 2000, 61, 316–320. [Google Scholar] [CrossRef]
- Fornazari, F.; da Silva, R.C.; Richini-Pereira, V.B.; Beserra, H.E.O.; Luvizotto, M.C.R.; Langoni, H. Comparison of Conventional PCR, Quantitative PCR, Bacteriological Culture and the Warthin Starry Technique to Detect Leptospira spp. in Kidney and Liver Samples from Naturally Infected Sheep from Brazil. J. Microbiol. Methods 2012, 90, 321–326. [Google Scholar] [CrossRef]
- Campos, Â.P.; Miranda, D.F.H.; Rodrigues, H.W.S.; da Silva Carneiro Lustosa, M.; Martins, G.H.C.; Mineiro, A.L.B.B.; Castro, V.; Azevedo, S.S.; de Sousa Silva, S.M.M. Seroprevalence and Risk Factors for Leptospirosis in Cattle, Sheep, and Goats at Consorted Rearing from the State of Piauí, Northeastern Brazil. Trop. Anim. Health Prod. 2017, 49, 899–907. [Google Scholar] [CrossRef] [PubMed]
- da Costa, D.F.; da Silva, A.F.; de Brasil, A.W.L.; Loureiro, A.P.P.; dos Santos, F.A.; de Azevedo, S.S.; Lilenbaum, W.; Alves, C.J. Leptospirosis in Native Mixed-Breed Sheep Slaughtered in a Semiarid Region of Brazil. Ciência Rural 2016, 47. [Google Scholar] [CrossRef] [Green Version]
- Amorim, R.M.; Nascimento, E.M.; Santarosa, B.P.; Dantas, G.N.; Ferreira, D.O.L.; Gonçalves, R.C.; Ullmann, L.S.; Langoni, H. Soroprevalência Da Leptospirose Em Ovinos Da Região Centrooeste Do Estado De São Paulo. Veterinária E Zootec. 2016, 23, 297–305. [Google Scholar]
- Bellocchi, G.; Martin, R.; Shtiliyanova, A.; Touhami, H.B.; Carrère, P. Vul’Clim–Climate Change Vulnerability Studies in the Region Auvergne (France). FACCE MACSUR Mid-term Scientific Conference, “Achievements, Activities, Advancement", Sassari, Italy, 1–4 April 2014. [Google Scholar]
- Nau, L.H.; Obiegala, A.; Król, N.; Mayer-Scholl, A.; Pfeffer, M. Survival Time of Leptospira Kirschneri Serovar Grippotyphosa under Different Environmental Conditions. PLoS ONE 2020, 15, e0236007. [Google Scholar] [CrossRef]
- Meylan, A. Fossorial Forms of the Water Vole, Arvicola Terresths (L.), in Europe 1. EPPO Bull. 1977, 7, 209–218. [Google Scholar] [CrossRef]
- Ayral, F.; Djelouadji, Z.; Raton, V.; Zilber, A.-L.; Gasqui, P.; Faure, E.; Baurier, F.; Vourc’h, G.; Kodjo, A.; Combes, B. Hedgehogs and Mustelid Species: Major Carriers of Pathogenic Leptospira, a Survey in 28 Animal Species in France (20122015). PLoS ONE 2016, 11, e0162549. [Google Scholar] [CrossRef] [PubMed]
- Ayral, F.C.; Bicout, D.J.; Pereira, H.; Artois, M.; Kodjo, A. Distribution of Leptospira Serogroups in Cattle Herds and Dogs in France. Am. J. Trop. Med. Hyg. 2014, 91, 756–759. [Google Scholar] [CrossRef] [PubMed]
- Bregoli, M.; Pesaro, S.; Ustulin, M.; Vio, D.; Beraldo, P.; Galeotti, M.; Cocchi, M.; Lucchese, L.; Bertasio, C.; Boniotti, M.B.; et al. Environmental Exposure of Wild Carnivores to Zoonotic Pathogens: Leptospira Infection in the First Free Living Wolf (Canis Lupus Linnaeus, 1758) Found Dead in the Friuli Venezia Giulia Region. Int. J. Environ. Res. Public Health 2021, 18, 2512. [Google Scholar] [CrossRef]
- Baranton, G.; Postic, D. Trends in Leptospirosis Epidemiology in France. Sixty-Six Years of Passive Serological Surveillance from 1920 to 2003. Int. J. Infect. Dis. 2006, 10, 162–170. [Google Scholar] [CrossRef]
- Hamond, C.; Martins, G.; Bremont, S.; Medeiros, M.A.; Bourhy, P.; Lilenbaum, W. Molecular Characterization and Serology of Leptospira Kirschneri (Serogroup Grippotyphosa) Isolated from Urine of a Mare Post-Abortion in Brazil. Zoonoses Public Health 2016, 63, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Soares, P.M.; Gomes, D.O.; Macedo, F.P.; Soares, M.M.; Lemes, K.R.; Jaeger, L.H.; Lilenbaum, W.; Lima, A.M.C. Serological and Molecular Characterization of Leptospira Kirschneri Serogroup Grippotyphosa Isolated from Bovine in Brazil. Microb. Pathog. 2020, 138, 103803. [Google Scholar] [CrossRef] [PubMed]
- Haller, O. Current Topics in Microbiology and Immunology. Curr. Top. Microbiol. Immunol. 1981, 92, 25–52. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Subcategory | Total (%) (n = 46) | Positive (%) (n = 35) | Negative (%) (n = 11) | p-Value * |
|---|---|---|---|---|---|
| Sex | Male | 20 (43.5) | 15 (42.9) | 5 (45.4) | 1 |
| Female | 26 (56.5) | 20 (57.1) | 6 (54.5) | ||
| Sexual maturity | Mature | 26 (56.5) | 22 (62.9) | 4 (36.4) | 0.2 |
| Immature | 20 (43.5) | 13 (37.1) | 7 (63.6) | ||
| Sampling category | Euthanized | 26 (56.5) | 25 (71.4) | 1 (9.1) | 0.001 |
| Cold-stored | 20 (43.5) | 10 (28.6) | 10 (90.9) | ||
| Site | Site 1 | 16 (34.8) | 15 (42.9) | 1 (9.1) | - |
| Site 2 | 9 (19.6) | 8 (22.9) | 1 (9.1) | 1 | |
| Site 3 | 21 (45.6) | 12 (34.2) | 9 (81.8) | 0.3 | |
| Overall length (cm) | Median (Q1–Q3) | 21 (18.25–22.6) | 21 (19.5–22.8) | 17 (16.5–21.8) | 0.03 |
| Voles | RT-PCR Positive | Presence of L. kirschneri | O-Antigen—GRIP | VNTR Profile 1-2-11 | VNTR Profile 1-2-X |
|---|---|---|---|---|---|
| Nb/Nbt | 35/46 | 35/35 | 19/35 | 9/14 † | 5/14 † |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harran, E.; Pinot, A.; Kodjo, A.; Djelouadji, Z.; Le Gudayer, M.; Sionfoungo Daouda, S.; Groud, K.; Lattard, V.; Ayral, F. Identification of Pathogenic Leptospira kirschneri Serogroup Grippotyphosa in Water Voles (Arvicola terrestris) from Ruminant Pastures in Puy-de-Dôme, Central France. Pathogens 2023, 12, 260. https://doi.org/10.3390/pathogens12020260
Harran E, Pinot A, Kodjo A, Djelouadji Z, Le Gudayer M, Sionfoungo Daouda S, Groud K, Lattard V, Ayral F. Identification of Pathogenic Leptospira kirschneri Serogroup Grippotyphosa in Water Voles (Arvicola terrestris) from Ruminant Pastures in Puy-de-Dôme, Central France. Pathogens. 2023; 12(2):260. https://doi.org/10.3390/pathogens12020260
Chicago/Turabian StyleHarran, Elena, Adrien Pinot, Angeli Kodjo, Zouheira Djelouadji, Marine Le Gudayer, Soro Sionfoungo Daouda, Karine Groud, Virginie Lattard, and Florence Ayral. 2023. "Identification of Pathogenic Leptospira kirschneri Serogroup Grippotyphosa in Water Voles (Arvicola terrestris) from Ruminant Pastures in Puy-de-Dôme, Central France" Pathogens 12, no. 2: 260. https://doi.org/10.3390/pathogens12020260