


Emerging and Re-Emerging Diseases Caused by Badnaviruses

Abstract
:1. Introduction
2. Emerging Disease Problems in Plants
2.1. Banana
2.2. Black Pepper
2.3. Citrus
2.4. Cacao
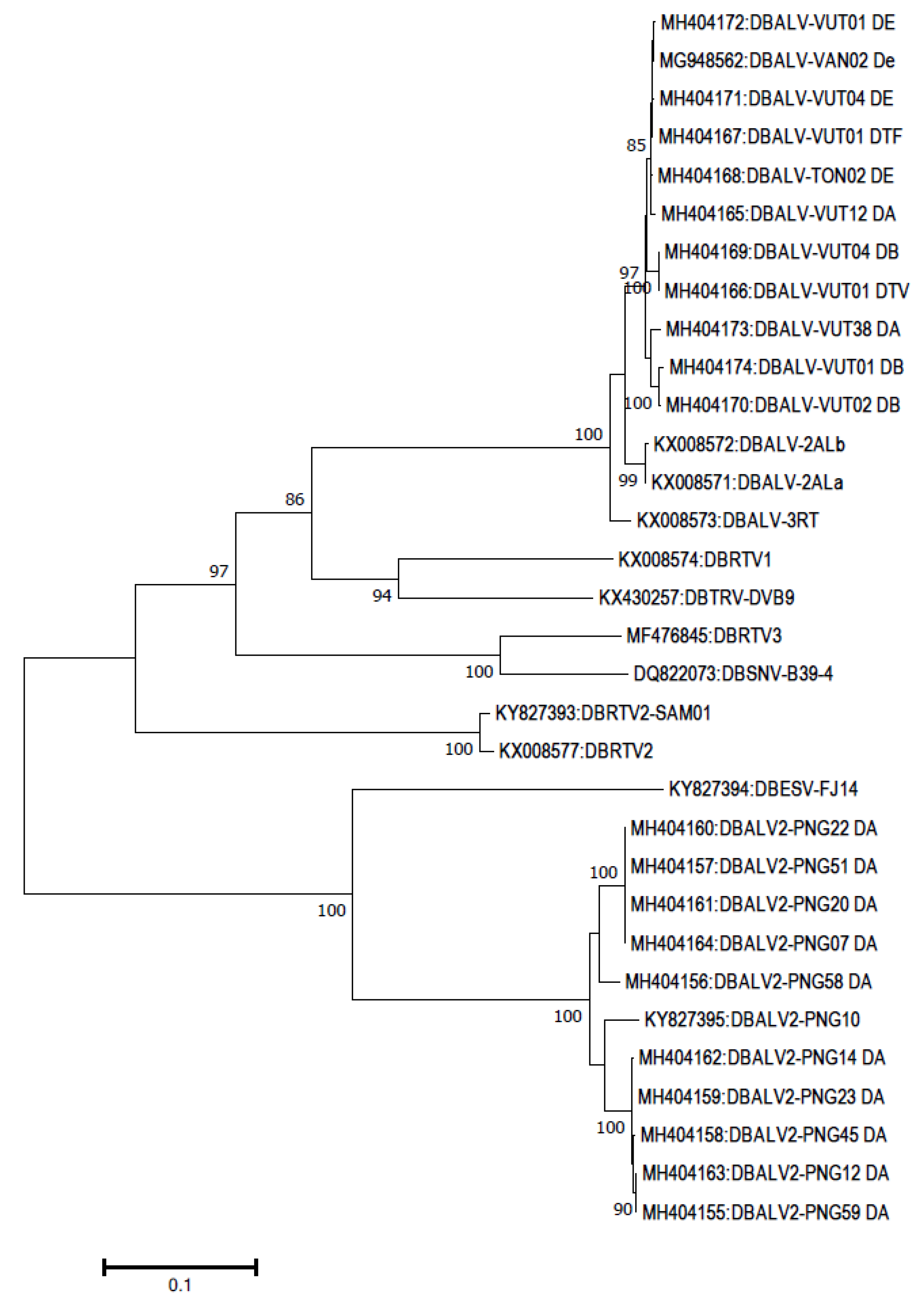
2.5. Dioscorea
2.6. Grapevine
2.7. Pineapple
2.8. Sugarcane
2.9. Sweet Potato
2.10. Taro
2.11. Badnaviruses Infecting Other Plants
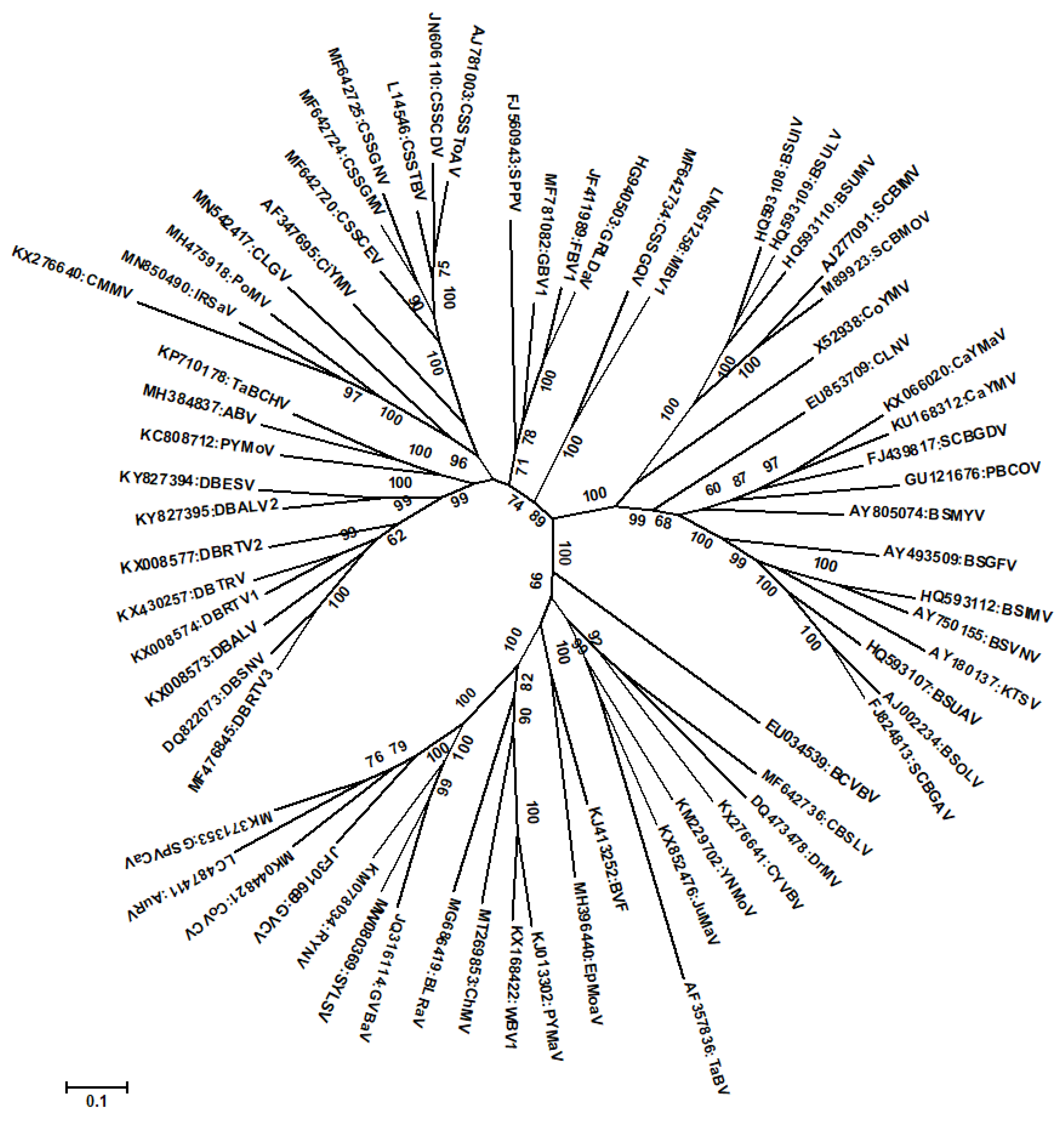
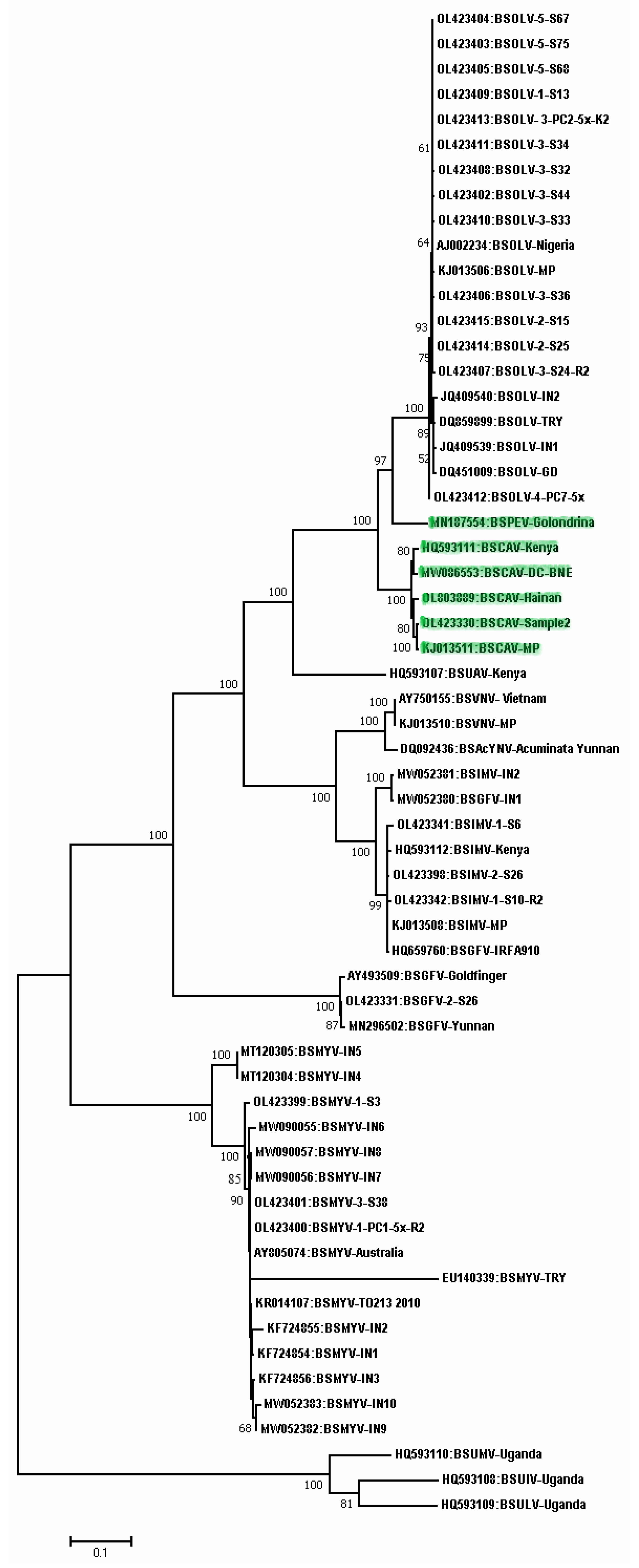
3. Recombination among Badnaviruses
4. Conclusions and Future Research Needs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- King, A.M.Q.; Adams, M.J.; Lefkowitz, E.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Academic Press: San Diego, CA, USA, 2012; 1463p. [Google Scholar]
- International Committee on Taxonomy of Viruses (ICTV). Virus Taxonomy: 2021 Release. Available online: http://www.ictvonline.org/virusTaxonomy.asp (accessed on 28 November 2022).
- Chabannes, M.; Iskra-Caruana, M.-L. Endogenous pararetroviruses—A reservoir of virus infection in plants. Curr. Opin. Virol. 2013, 3, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Geering, A.D.W.; Hull, R. Family Caulimoviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Academic Press: Boston, MA, USA, 2012; pp. 429–443. [Google Scholar]
- Bhat, A.I.; Hohn, T.; Selvarajan, R. Badnaviruses: The current global scenario. Viruses 2016, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Borah, B.K.; Sharma, S.; Kant, R.; Anthony-Johnson, A.M.; Saigopal, D.V.R.; Dasgupta, I. Bacilliform DNA-containing plant viruses in the tropics: Commonalities within a genetically diverse group. Mol. Plant Pathol. 2013, 14, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Hohn, T.; Richert-Poggeler, K.R.; Harper, G.; Schawarzacher, T.; Teo, C.; Teycheney, P.-Y.; Iskra-Caruana, M.-L.; Hull, R. Evolution of integrated plant viruses. In Plant Virus Evolution; Roosinck, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 54–76. [Google Scholar]
- Staginnus, C.; Iskra-Caruana, M.; Lockhart, B.; Hohn, T.; Richert-Pöggeler, K.R. Suggestions for a nomenclature of endogenous pararetroviral sequences in plants. Arch. Virol. 2009, 154, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Dallot, S.; Accuna, P.; Rivera, C.; Ramirez, P.; Cote, F.; Lockhart, B.E.L.; Caruana, M.-L. Evidence that the proliferation stage of micropropagation procedure is determinant in the expression of Banana streak virus integrated into the genome of the FHIA21 hybrid (Musa AAAB). Arch. Virol. 2001, 146, 2179–2190. [Google Scholar] [CrossRef]
- Lheureux, F.; Carreel, F.; Jenny, C.; Lockhart, B.E.L.; Iskra-Caruana, M.-L. Identification of genetic markers linked to banana streak disease expression in inter-specific Musa hybrids. Theor. Appl. Genet. 2003, 106, 594–598. [Google Scholar] [CrossRef]
- Côte, F.X.; Galzi, S.; Folliot, M.; Lamagnère, Y.; Teycheney, P.-Y.; Iskra-Caruana, M.-L. Micropropagation by tissue culture triggers differential expression of infectious endogenous Banana streak virus sequences (eBSV) present in the B genome of natural and synthetic interspecific banana plantains. Mol. Plant Pathol. 2010, 11, 137–144. [Google Scholar] [CrossRef]
- Harper, G.; Osuji, J.O.; Heslop-Harrison, J.S.; Hull, R. Integration of banana streak badnavirus into the Musa genome: Molecular and cytogenetic evidence. Virology 1999, 255, 207–213. [Google Scholar] [CrossRef]
- Harper, G.; Hull, R.; Lockhart, B.E.N.; Olsweski, N. Viral sequences integrated in to plant genomes. Ann. Rev. Phytopathol. 2002, 40, 119–136. [Google Scholar] [CrossRef]
- Staginnus, C.; Richert-Poggeler, K.R. Endogenous pararetroviruses: Two-faced travelers in the plant genome. Trends Plant Sci. 2006, 11, 485–491. [Google Scholar] [CrossRef]
- Mette, M.F.; Kanno, T.; Aufsatz, W.; Jakowitsch, J.; Van der Winden, J.; Matzke, M.A.; Matzke, A.J.M. Endogenous viral sequences and their potential contribution to heritable virus resistance in plants. EMBO J. 2002, 21, 461–469. [Google Scholar] [CrossRef]
- Ndowora, T.; Ganesh, D.; Daryl, L.F.; Harper, G.; Hull, R.; Neil, E.O.; Lockhart, B. Evidence that Badnavirus infection in Musa can originate from integrated Pararetroviral sequences. Virology 1999, 225, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Dahal, G.; Hughes, J.D.A.; Thottappilly, G.; Lockhart, B.E.L. Effect of temperature on symptom expression and reliability of banana streak badnavirus detection in naturally infected plantain and banana (Musa spp.). Plant Dis. 1998, 82, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Dahal, G.; Ortiz, R.; Tenkouano, A.; Hughes, J.D.A.; Thottappilly, G.; Vuylsteke, D.; Lockhart, B.E.L. Relationship between natural occurrence of Banana streak badnavirus and symptom expression, relative concentration of viral antigen, and yield characteristics of some micropropagated Musa spp. Plant Pathol. 2000, 49, 68–79. [Google Scholar] [CrossRef]
- Daniells, J.W.; Geering, A.D.W.; Bryde, N.J.; Thomas, J.E. The effect of Banana streak virus on the growth and yield of dessert bananas in tropical Australia. Ann. Appl. Biol. 2001, 139, 51–60. [Google Scholar] [CrossRef]
- Lockhart, B.E.L. Purification and serology of a bacilliform virus associated with banana streak disease. Phytopathology 1986, 76, 995–999. [Google Scholar] [CrossRef]
- Geering, A.D.W.; McMichael, L.A.; Dietzgen, R.G.; Thomas, J.E. Genetic diversity among banana streak virus isolates from Australia. Phytopathology 2000, 90, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Geering, A.D.W.; Olszewski, N.E.; Harper, G.; Lockhart, B.E.L.; Hull, R.; Thomas, J.E. Banana contains a diverse array of endogenous badnaviruses. J. Gen. Virol. 2005, 86, 511–520. [Google Scholar] [CrossRef]
- Harper, G.; Hart, D.; Moult, S.; Hull, R.; Thresh, J.M.; Jones, R.A.C.; Kuehne, T. Banana streak virus is very diverse in Uganda. Virus Res. 2004, 100, 51–56. [Google Scholar] [CrossRef]
- Harper, G.; Hart, D.; Moult, S.; Hull, R.; Geering, A.; Thomas, J. The diversity of Banana streak virus isolates in Uganda. Arch. Virol. 2005, 150, 2407–2420. [Google Scholar] [CrossRef]
- Geering, A.D.; Parry, J.N.; Thomas, J.E. Complete genome sequence of a novel badnavirus, Banana streak IM virus. Arch. Virol. 2011, 156, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Lheureux, F.; Laboureau, N.; Muller, E.; Lockhart, B.E.; Iskra-Caruana, M.L. Molecular characterization of banana streak acuminata Vietnam virus isolated from Musa acuminata siamea (banana cultivar). Arch. Virol. 2007, 152, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Iskra-Caruana, M.L.; Duroy, P.O.; Chabannes, M.; Muller, E. The common evolutionary history of badnaviruses and banana. Infect. Genet. Evol. 2014, 21, 83–89. [Google Scholar] [CrossRef]
- Chabannes, M.; Gabriel, M.; Aksa, A.; Galzi, S.; Dufayard, J.F.; Iskra-Caruana, M.L.; Muller, E. Badnaviruses and banana genomes: A long association sheds light on Musa phylogeny and origin. Mol. Plant Pathol. 2021, 22, 216–230. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Vignesh Kumar, P.; Geetanjali, A.S.; Pun, K.B.; Baranwal, V.K. Subpopulation level variation of banana streak viruses in India and common evolution of banana and sugarcane badnaviruses. Virus Genes 2015, 50, 450–465. [Google Scholar] [CrossRef]
- Delanoy, M.; Salmon, M.; Kummert, J.; Frison, E.; Lepoivre, P. Development of real-time PCR for the rapid detection of episomal Banana streak virus (BSV). Plant Dis. 2003, 87, 33–38. [Google Scholar] [CrossRef]
- Le Provost, G.; Iskra-Caruana, M.L.; Acina, I.; Teycheney, P.Y. Improved detection of episomal banana streak viruses by multiplex immunocapture PCR. J. Virol. Methods 2006, 137, 7–13. [Google Scholar] [CrossRef]
- James, A.P.; Geijskes, R.J.; Dale, J.L.; Harding, R.M. Development of a novel rolling-circle amplification technique to detect Banana streak virus which also discriminates between integrated and episomal virus sequences. Plant Dis. 2011, 95, 57–62. [Google Scholar] [CrossRef]
- Javer-Higginson, E.; Acina-Mambole, I.; González, J.E.; Font, C.; González, G.; Echemendía, A.L.; Muller, E.; Teycheney, P.-Y. Occurrence, prevalence and molecular diversity of banana streak viruses in Cuba. Eur. J. Plant Pathol. 2013, 138, 157–166. [Google Scholar] [CrossRef]
- Sharma, S.K.; Vignesh Kumar, P.; Baranwal, V.K. Immunodiagnosis of episomal Banana streak MY virus using polyclonal antibodies to an expressed putative coat protein. J. Virol. Methods 2014, 207, 86–94. [Google Scholar] [CrossRef]
- Selvarajan, R.; Sheeba, M.M.; Balasubramanian, V. Simultaneous detection of episomal Banana streak Mysore virus and Banana bunchy top virus using multiplex RT-PCR. Curr. Sci. 2011, 100, 31–34. [Google Scholar]
- Liu, F.X.; Feng, L.X.; Chen, X.; Han, Y.C.; Li, W.D.; Xu, W.; Cai, B.; Lin, M.G. Simultaneous detection of four banana viruses by multiplex PCR. J. Phytopathol. 2012, 160, 622–627. [Google Scholar] [CrossRef]
- Selvarajan, R.; Balasubramanian, V.; Gayathrie, T. Highly efficient immunodiagnosis of episomal Banana streak MY virus (BSMYV) using polyclonal antibodies raised against recombinant viral associated protein. J. Phytopathol. 2016, 207, 86–94. [Google Scholar]
- Zhang, J.; Borth, W.; Lin, B.; Melzer, M.; Shen, H.; Pu, X.; Sun, D.; Nelson, S.; Hu, J. Multiplex detection of three banana viruses by reverse transcription loop-mediated isothermal amplification (RT-LAMP). Trop. Plant Pathol. 2018, 43, 543–551. [Google Scholar] [CrossRef]
- Kapoor, R.; Srivastava, N.; Kumar, R.; Sharma, S.K.; Rai, R.; Kumar, S.; Baranwal, V.K. Detection of episomal banana streak Mysore virus by reverse transcription-recombinase polymerase amplification assay. J. Plant Pathol. 2020, 102, 499–503. [Google Scholar] [CrossRef]
- Kuan, C.P.; Tsai, C.H.; Tseng, C.S.; Yang, T.C. Development of a bead-based assay for detection of three banana-infecting viruses. Peer J. 2022, 26, e13409. [Google Scholar] [CrossRef]
- Helliot, B.; Panis, B.; Poumay, Y.; Swennen, R.; Lepoivre, P.; Frison, E. Cryopreservation for the elimination of cucumber mosaic and banana streak viruses from banana (Musa spp.). Plant Cell Rep. 2002, 20, 1117–1122. [Google Scholar] [CrossRef]
- Helliot, B.; Panis, B.; Frison, E.; De Clercq, E.; Swennen, R.; Lepoivre, P.; Neyts, J. The acyclic nucleoside phosphonate analogues, adefovir, tenofovir and PMEDAP, efficiently eliminate banana streak virus from banana (Musa) sp. Antivir. Res. 2003, 59, 121–126. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Ntui, V.O.; Ron, M.; Muiruri, S.K.; Britt, A.; Tripathi, L. CRISPR/Cas9 editing of endogenous banana streak virus in the B genome of Musa spp. overcomes a major challenge in banana breeding. Commun. Biol. 2019, 2, 46. [Google Scholar] [CrossRef]
- Lockhart, B.E.L.; Kirtisak, K.A.; Jones, P.; Padmini, D.D.; Olszieewski, N.E.; Lockhart, N.; Nuarchan, D.; Sangalang, J. Identification of Piper yellow mottle virus, a mealy bug transmited badnavirus infecting Piper spp. in south East Asia. Eur. J. Plant Pathol. 1997, 103, 303–311. [Google Scholar] [CrossRef]
- De Silva, D.P.; Jones, P.P.; Shaw, M.W. Identification and transmission of Piper yellow mottle virus and Cucumber mosaic virus infecting black pepper (Piper nigrum) in Sri Lanka. Plant Pathol. 2002, 51, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Bhat, A.I.; Devasahayam, S.; Sarma, Y.R.; Pant, R.P. Association of a Badnavirus in black pepper (Piper nigrum L.) transmitted by mealybug (Ferrisia virgata) in India. Curr. Sci. 2003, 84, 1547–1550. [Google Scholar]
- Che, H.; Cao, X.; Liu, P. Occurrence and characterization of virus species associated with black pepper (piper nigrum l.) virus diseases in Hainan province, China. J. Phytopathol. 2020, 169, 247–252. [Google Scholar] [CrossRef]
- Bhat, A.I.; Biju, C.N.; Srinivasan, V.; Ankegowda, S.J.; Krishnamurthy, K.S. Current status of viral diseases affecting black pepper and cardamom. J. Spices Aromat. Crops 2018, 27, 1–16. [Google Scholar]
- Ahamedemujtaba, V.; Atheena, P.V.; Bhat, A.I.; Krishnamurthy, K.S.; Srinivasan, V. Symptoms of piper yellow mottle virus in black pepper as influenced by temperature and relative humidity. Virus Dis. 2021, 32, 305–313. [Google Scholar] [CrossRef]
- Duarte, M.L.R.; Albuquerque, F.C.; Poltranieri, L.S.; Trindade, D.R.; Kitajima, E.W.; Brioso, P.S.T. Mosqueado Amarelo da Pimenta-do-Reino; Embrapa Amazônia Oriental: Belém, Brazil, 2001; p. 30. [Google Scholar]
- Siju, S.; Bhat, A.I.; Hareesh, P.S. Identification and characterization of a Badnavirus infecting betel vine and Indian long pepper. J. Plant Biochem. Biotechnol. 2008, 17, 73–76. [Google Scholar] [CrossRef]
- Bhat, A.I.; Sasi, S.; Revathy, K.A.; Deeshma, K.P.; Saji, K.V. Sequence diversity among badnavirus isolates infecting black pepper and related species in India. Virus Dis. 2014, 25, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Hany, U.; Adams, I.P.; Glover, R.; Bhat, A.I.; Boonham, N. The complete genome sequence of Piper yellow mottle virus (PYMoV). Arch. Virol. 2014, 159, 385–388. [Google Scholar] [CrossRef]
- Deeshma, K.P.; Bhat, A.I. Complete genome sequencing of Piper yellow mottle virus infecting black pepper, betelvine and Indian long pepper. Virus Genes 2015, 50, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.W.T.; Liew, J.X.; Chung, H.H. Piper yellow mottle virus: A deep dive into the genome. Gene Rep. 2022, 29, 101680. [Google Scholar] [CrossRef]
- Hareesh, P.S.; Bhat, A.I. Detection and partial nucleotide sequence analysis of Piper yellow mottle virus infecting black pepper in India. Indian J. Virol. 2008, 19, 160–167. [Google Scholar]
- Bhat, A.I.; Siljo, A.; Deeshma, K.P. Rapid detection of Piper yellow mottle virus and Cucumber mosaic virus infecting black pepper (Piper nigrum) by loop-mediated isothermal amplification (LAMP). J. Virol. Methods 2013, 193, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.I.; Siljo, A. Detection of viruses infecting black pepper by SYBR Green-based real-time PCR assay. J. Plant Pathol. 2014, 96, 105–109. [Google Scholar]
- Mohandas, A.; Bhat, A.I. Recombinase polymerase amplification assay for the detection of piper yellow mottle virus infecting black pepper. Virus Dis. 2020, 31, 38–44. [Google Scholar] [CrossRef]
- Sasi, S.; Bhat, A.I. In vitro elimination of Piper yellow mottle virus from infected black pepper through somatic embryogenesis and meristem-tip culture. Crop Prot. 2018, 103, 39–45. [Google Scholar] [CrossRef]
- Srinivasan, V.; Bhat, A.I.; Dinesh, R.; Ankegowda, S.J.; Hamza, S.; Biju, C.N.; Krishnamurthy, K.S. Rejuvenation of virus affected black pepper plantations through soil and plant health management. Indian J. Arecanut Spices Med. Plants 2017, 19, 24–27. [Google Scholar]
- Ahlawat, Y.S.; Pant, R.P.; Lockhart, B.E.L.; Srivastava, M.; Chakraborty, N.K.; Varma, A. Association of a badnavirus with citrus mosaic disease in India. Plant Dis. 1996, 80, 590–592. [Google Scholar] [CrossRef]
- Huang, Q.; Hartung, J.S. Cloning and sequence analysis of an infectious clone of Citrus yellow mosaic virus that can infect sweet orange via Agrobacterium-mediated inoculation. J. Gen. Virol. 2001, 82, 2549–2558. [Google Scholar] [CrossRef]
- Vadlamudi, T.; Kaldis, A.; Divi, V.S.G.; Patil, B.L.; Voloudakis, A.E. The Citrus yellow mosaic badnavirus ORFI functions as a RNA-silencing suppressor. Virus Genes 2021, 57, 469–473. [Google Scholar] [CrossRef]
- Borah, B.K.; Johnson, A.M.A.; Sai-Gopal, D.V.R.; Dasgupta, I. Sequencing and computational analysis of complete genome sequences of Citrus yellow mosaic badnavirus from acid lime and pummelo. Virus Genes 2009, 39, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Anthony-Johnson, A.M.; Borah, B.K.; Sai Gopal, D.V.; Dasgupta, I. Analysis of full-length sequences of two Citrus yellow mosaic badnavirus isolates infecting Citrus jambhiri (Rough Lemon) and Citrus sinensis L. Osbeck (Sweet Orange) from a nursery in India. Virus Genes 2012, 45, 600–605. [Google Scholar] [CrossRef]
- Geering, A.D.W.; Maumus, F.; Copetti, D.; Choisne, N.; Zwickl, D.J.; Zytnicki, M.; McTaggart, A.R.; Scalabrin, S.; Vezzulli, S.; Wing, R.A.; et al. Endogenous florendoviruses are major components of plant genomes and hallmarks of virus evolution. Nat. Commun. 2014, 5, 5269. [Google Scholar] [CrossRef]
- Kumar, P.V.; Sharma, S.K.; Rishi, N.; Ghosh, D.K.; Baranwal, V.K. An isothermal based recombinase polymerase amplification assay for rapid, sensitive, and robust indexing of citrus yellow mosaic virus. Acta Virol. 2018, 62, 104–108. [Google Scholar] [CrossRef]
- Ghosh, D.K.; Aglave, B.; Bhanare, K.; Baranwal, V.K. PCR based detection of Citrus yellow mosaic disease from Vidarbha region of Maharastra. Indian Phytopathol. 2007, 60, 520–526. [Google Scholar]
- Gopi, V.; Gopal, K.; Gourisankar, T.; Palanivel, S. Detection of citrus yellow mosaic virus by PCR and nucleic acid spot hybridisation using non-radioactive probes in commercial citrus species. Arch. Phytopathol. Plant Prot. 2010, 43, 892–899. [Google Scholar] [CrossRef]
- Motghare, M.; Dhar, A.K.; Kokane, A.; Warghane, A.; Kokane, S.; Sharma, A.K.; Reddy, M.K.; Ghosh, D.K. Quantitative Distribution of Citrus yellow mosaic badnavirus in Sweet orange (Citrus sinensis) and its Implication in Developing Disease Diagnostics. J. Virol. Methods 2018, 259, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Thresh, M.J. The origin and epidemiology of some important plant virus diseases. Appl. Biol. 1980, 5, 1–65. [Google Scholar]
- Ramos-Sobrinho, R.; Kouakou, K.; Bi, A.B.; Keith, C.V.; Diby, L.; Kouame, C.; Aka, R.A.; Marelli, J.-P.; Brown, J.K. Molecular detection of cacao swollen shoot badnavirus species by amplification with four PCR primer pairs, and evidence that Cacao swollen shoot Togo B virus-like isolates are highly prevalent in Côte d’Ivoire. Eur. J. Plant Pathol. 2021, 159, 941–947. [Google Scholar] [CrossRef]
- Kandito, A.; Hartono, S.; Trisyono, Y.A.; Somowiyarjo, S. First report of Cacao mild mosaic virus associated with cacao mosaic disease in Indonesia. New Dis. Rep. 2022, 45, e12071. [Google Scholar] [CrossRef]
- Muller, E.; Ravel, S.; Agret, C.; Abrokwah, F.; Dzahini-Obiatey, H.; Galyuon, I.; Kouakou, K.; Jeyaseelan, E.C.; Allainguillaume, J.; Wetten, A. Next generation sequencing elucidates cacao badnavirus diversity and reveals the existence of more than ten viral species. Virus Res. 2018, 244, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Chingandu, N.; Zia-ur-rehman, M.; Sreenivasan, T.N.; Surujdeo-Maharaj, S.; Umaharan, P.; Gutierrez, O.A.; Brown, J.K. Molecular characterization of previously elusive badnaviruses associated with symptomatic cacao in the New World. Arch. Virol. 2017, 162, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Daymond, A.J.; Hadley, P.; End, M.J.; Umaharan, P.; Dunwell, J.M. Identification of Cacao Mild Mosaic Virus (CaMMV) and Cacao Yellow Vein-Banding Virus (CYVBV) in Cocoa (Theobroma cacao) Germplasm. Viruses 2021, 13, 2152. [Google Scholar] [CrossRef] [PubMed]
- Posnette, A.F. Virus diseases of cacao in West Africa VII: Virus transmission by different vector species. Ann. Appl. Biol. 1950, 37, 378–384. [Google Scholar] [CrossRef]
- Marelli, J.P.; Guest, D.I.; Bailey, B.A.; Evans, H.C.; Brown, J.K.; Junaid, M.; Barreto, R.W.; Lisboa, D.O.; Puig, A.S. Chocolate under threat from old and new cacao diseases. Phytopathology 2019, 109, 1331–1343. [Google Scholar] [CrossRef]
- Posnette, A.F. Virus diseases of cacao in West Africa. 1. Cacao viruses 1A, 1B, 1C and 1D. Ann. Appl. Biol. 1947, 34, 388–402. [Google Scholar] [CrossRef]
- Jacquot, E.; Hagen, L.S.; Michler, P.; Rohfritsch, O.; Stussi-Garaud, C.; Keller, M.; Jacquemond, M.; Yot, P. In situ localisation of cacao swollen shoot virus in agroinoculated Theobroma cacao. Arch. Virol. 1999, 144, 259–271. [Google Scholar] [CrossRef]
- Roivainen, O. Transmission of cocoa viruses by mealy bugs (Homoptera: Pseudococcidae). J. Sci. Agric. Soc. 1976, 48, 433–453. [Google Scholar]
- Quainoo, A.K.; Wetten, A.C.; Allainguillaume, J. Transmission of cocoa swollen shoot virus by seeds. J. Virol. Methods 2008, 150, 45–49. [Google Scholar] [CrossRef]
- Ameyaw, G.A.; Wetten, A.; Dzahini-Obiatey, H.; Domfeh, O.; Allainguillaume, J. Investigation on Cacao swollen shoot virus (CSSV) pollen transmission through cross-pollination. Plant Pathol. 2013, 62, 421–427. [Google Scholar] [CrossRef]
- Abrokwah, F.K.; Dabie, K.; Adulley, F. Genome variability, species diversity, phylogenetic relationships, origin, and geographical distribution of badnaviruses involved in the cacao swollen shoot disease: The case of West Africa. Trop. Plant Pathol. 2022, 47, 201–213. [Google Scholar] [CrossRef]
- Hagen, L.S.; Jacquemond, M.; Lepingle, A.; Lot, H.; Tepfer, M. Nucleotide sequence and genomic organization of Cacao swollen shoot virus. Virology 1993, 196, 619–628. [Google Scholar] [CrossRef]
- Jacquot, E.; Hagen, L.S.; Jacquemond, M.; Yot, P. The open reading frame 2 product of cacao swollen shoot badnavirus is a nucleic acid-binding protein. Virology 1996, 225, 191–195. [Google Scholar] [CrossRef]
- Chingandu, N.; Kouakou, K.; Aka, R.; Ameyaw, G.; Gutierrez, O.A.; Herrmann, H.; Brown, J.K. The proposed new species, Cacao red vein virus, and three previously recognized badnavirus species are associated with cacao swollen shoot disease. Virol. J. 2017, 14, 199. [Google Scholar] [CrossRef]
- Chingandu, N.; Kouakou, K.; Aka, R.; Gutierrez, O.A.; Brown, J.K. Unexpected genome variability at multiple loci suggests Cacao swollen shoot virus comprises multiple, divergent molecular variants. J. Emerg. Dis. Virol. 2017, 3, 2473-1846. [Google Scholar]
- Chingandu, N.; Dongo, L.; Gutierrez, O.A.; Brown, J.K. The previously unidentified, divergent badnavirus species Cacao red vein-banding virus is associated with cacao swollen shoot disease in Nigeria. Plant Dis. 2019, 103, 1302–1308. [Google Scholar] [CrossRef]
- Ramos-Sobrinho, R.; Chingandu, N.; Gutierrez, O.A.; Marelli, J.-P.; Brown, J.K. A Complex of Badnavirus Species Infecting Cacao Reveals Mixed Infections, Extensive Genomic Variability, and Interspecific Recombination. Viruses 2020, 12, 443. [Google Scholar] [CrossRef] [PubMed]
- Muller, E.; Ullah, I.; Dunwell, J.M.; Richardson, M.; Allainguillaume, J.; Wetten, A. Identification and distribution of novel badnaviral sequences integrated in the genome of cacao (Theobroma cacao). Sci. Rep. 2021, 11, 8270. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Sobrinho, R.; Ferro, M.M.M.; Nagata, T.; Puig, A.S.; Keith, C.V.; Britto, D.S.; Gutierrez, O.A.; Mareli, J.P.; Brown, J.K. Complete genome sequences of three newly discovered cacao mild mosaic virus isolates from Theobroma cacao L. in Brazil and Puerto Rico and evidence for recombination. Arch. Virol. 2021, 166, 2027–2031. [Google Scholar] [CrossRef] [PubMed]
- Ameyaw, G.A.; Domfeh, O.; Armooh, B.; Boakye, A.Y.; Arjarquah, A. Inconsistent PCR detection of Cacao swollen shoot virus (CSSV) is linked to the occurrence of different variants across the cocoa regions of Ghana. J. Virol. Methods 2022, 300, 114400. [Google Scholar] [CrossRef] [PubMed]
- Domfeh, O.; Ameyaw, G.A.; Gyamera, E.A. Mild strain cross-protection in cacao: Evaluation of symptom suppression in three progenies exposed to natural spread of the New Juabeng strain of the Cacao swollen shoot Togo B virus (CSSTBV). Trop. Plant Pathol. 2022, 47, 562–568. [Google Scholar] [CrossRef]
- Andres, C.; Gattinger, A.; Dzahini-Obiatey, H.K.; Blaser, W.J.; Offei, S.K.; Six, J. Combatting Cocoa Swollen Shoot Virus Disease: What do we know? Crop Prot. 2017, 98, 76–84. [Google Scholar] [CrossRef]
- Quainoo, A.K.; Wetten, A.C.; Allainguillaume, J. The effectiveness of somatic embryogenesis in eliminating the cocoa swollen shoot virus from infected cocoa trees. J. Virol. Methods 2008, 149, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Turaki, A.A.; Bömer, M.; Silva, G.; Lava Kumar, P.; Seal, S.E. PCR-DGGE analysis: Unravelling complex mixtures of badnavirus sequences present in yam germplasm. Viruses 2017, 9, 181. [Google Scholar] [CrossRef]
- Bömer, M.; Rathnayake, A.I.; Visendi, P.; Silva, G.; Seal, S.E. Complete genome sequence of a new member of the genus Badnavirus, Dioscorea bacilliform RT virus 3, reveals the first evidence of recombination in yam badnaviruses. Arch. Virol. 2018, 163, 533–538. [Google Scholar] [CrossRef]
- Bömer, M.; Turaki, A.; Silva, G.; Kumar, P.; Seal, S. A sequence-independent strategy for amplification and characterization of episomal badnavirus sequences reveals three previously uncharacterized yam badnaviruses. Viruses 2016, 8, 188. [Google Scholar] [CrossRef]
- Umber, M.; Gomez, R.; Gelabale, S.; Bonheur, L.; Pavis, C.; Teycheney, P. The genome sequence of Dioscorea bacilliform TR virus, a member of the genus Badnavirus infecting Dioscorea spp., sheds light on the possible function of endogenous Dioscorea bacilliform viruses. Arch. Virol. 2016, 162, 517–521. [Google Scholar] [CrossRef]
- Sukal, A.; Kidanemariam, D.; Dale, J.; James, A.; Harding, R. Characterization of badnaviruses infecting Dioscorea spp. in the Pacific reveals two putative novel species and the first report of Dioscorea bacilliform RT virus 2. Virus Res. 2017, 238, 29–34. [Google Scholar] [CrossRef]
- Diouf, M.B.; Festus, R.; Silva, G.; Guyader, S.; Umber, M.; Seal, S.; Teycheney, P.Y. Viruses of Yams (Dioscorea spp.): Current Gaps in Knowledge and Future Research Directions to Improve Disease Management. Viruses 2022, 14, 1884. [Google Scholar] [CrossRef]
- Umber, M.; Filloux, D.; Muller, E.; Laboureau, N.; Galzi, S.; Roumagnac, P.; Iskra-Caruana, M.-L.; Pavis, C.; Teycheney, P.-Y.; Seal, S. The genome of African yam (Dioscorea cayenensis-rotundata complex) hosts endogenous sequences from four distinct badnavirus species. Mol. Plant Pathol. 2014, 15, 790–801. [Google Scholar] [CrossRef]
- Filloux, D.; Fernandez, E.; Loire, E.; Claude, L.; Galzi, S.; Candresse, T.; Winter, S.; Jeeva, M.L.; Makeshkumar, T.; Martin. D.P.; et al. Nanopore-Based Detection and Characterization of Yam Viruses. Sci. Rep. 2018, 148, 17879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Singh, K.; Kaur, R.; Qiu, W. Association of a novel DNA virus with the grapevine vein-clearing and vine decline syndrome. Phytopathology 2011, 101, 1081–1090. [Google Scholar] [CrossRef]
- Cieniewicz, E.J.; Qiu, W.; Saldarelli, P.; Fuchs, M. Believing is seeing: Lessons from emerging viruses in grapevine. J. Plant Pathol. 2020, 102, 619–632. [Google Scholar] [CrossRef]
- Rumbos, I.C.; Avgelis, A.D. Roditis leaf discoloration—A new virus disease of grapevine: Symptomatology and transmission to indicators plants. Phytopathol. Z. 1989, 152, 274–278. [Google Scholar] [CrossRef]
- Maliogka, V.I.; Olmos, A.; Pappi, P.G.; Lotos, L.; Efthimiou, K.; Grammatikaki, G.; Candresse, T.; Katis, N.I.; Avgeli, A.D. A novel grapevine badnavirus is associated with the Roditis leaf discoloration disease. Virus Res. 2015, 203, 47–55. [Google Scholar] [CrossRef]
- Chiumenti, M.; Morelli, M.; Giampetruzzi, A.; Palmisano, F.; Savino, V.N.; La Notte, P.; Martelli, G.P.; Saldarelli, P. First report of grapevine Roditis leaf discoloration-associated virus in Italy. J. Plant Pathol. 2015, 97, 551. [Google Scholar]
- Uluba, S.; Serçe, Ç.; Altan, B.; Bolat, V.; Ayyaz, M.; Çifçi, O.; Önder, S.; Öztürk Gökçe, Z.N.; Maliogka, V.I. First Report of grapevine Roditis leaf discoloration-associated virus infecting grapevine (Vitis vinifera) in Turkey. Plant Dis. 2018, 2, 256. [Google Scholar]
- Bester, R.; Lotos, L.; Vermeulen, A.; Pietersen, G.; Maliogka, V.I.; Maree, H.J. Complete genome sequence of a grapevine Roditis leaf discoloration-associated virus (GRLDaV) variant from South Africa. Arch. Virol. 2021, 166, 2041–2044. [Google Scholar] [CrossRef]
- Ekemen, M. Investigation of Etiology on Grapevine Roditis Leaf Discoloration-Associated Virus. Master’s Thesis, Nigde Omer Halisdemir University, Niğde, Turkey, 2021. Available online: http://acikerisim.ohu.edu.tr/xmlui/handle/11480/8548 (accessed on 12 January 2023).
- Jagunic, M.; De Stradis, A.; Preiner, D.; La Notte, P.; Al Rwahnih, M.; Almeida, R.P.P.; Voncina, D. Biology and ultrastructural characterization of Grapevine Badnavirus 1 and Grapevine Virus G. Viruses 2022, 14, 2695. [Google Scholar] [CrossRef] [PubMed]
- Vončina, D.; Almeida, R.P.P. Screening of some Croatian autochthonous grapevine varieties reveals a multitude of viruses, including novel ones. Arch. Virol. 2018, 163, 2239–2243. [Google Scholar] [CrossRef]
- Wakman, W.D.; Teakle, D.S.; Thomas, J.E.; Dietzgen, R.G. Presence of clostero-like virus and a bacilliform virus in pineapple plants in Queensland. Aust. J. Agric. Res. 1995, 46, 947–958. [Google Scholar] [CrossRef]
- Thomson, K.G.; Dietzgen, R.G.; Thomas, J.E.; Teakle, D.S. Detection of pineapple bacilliform virus using the polymerase chain reaction. Ann. Appl. Biol. 1996, 129, 57–69. [Google Scholar] [CrossRef]
- Gambley, C.F.; Geering, A.D.; Steele, V.; Thomas, J.E. Identification of viral and non-viral reverse transcribing elements in pineapple (Ananas comosus), including members of two new badnavirus species. Arch. Virol. 2008, 153, 1599–1604. [Google Scholar] [CrossRef] [PubMed]
- Sether, D.M.; Melzer, M.J.; Borth, W.B.; Hu, J.S. Pineapple bacilliform CO virus: Diversity, detection, distribution, and transmission. Plant Dis. 2012, 96, 1798–1804. [Google Scholar] [CrossRef]
- Liting, W.; Xiaolei, R.; Wenjin, S. Sequencing and analysis of the complete genomic sequence of pineapple bacilliform comosus virus. China Agric. Sci. 2010, 43, 1969–1976. [Google Scholar]
- Autrey, L.J.C.; Boolell, S.; Jones, P. Distribution of sugarcane bacilliform virus in various geographical regions. In Proceedings of the XXI Congress of the International Society of Sugar Cane Technologists, Bangkok, Thailand, 5–14 March 1992; Kasetsart University: Bangkok, Thailand, 1995. [Google Scholar]
- Rao, G.P.; Sharma, S.K.; Kumar, P.V. Sugarcane Bacilliform Viruses: Present Status. In Plant Viruses; CRC Press: Boca Raton, FL, USA, 2018; pp. 117–130. [Google Scholar]
- Ahmad, K.; Sun, S.R.; Chen, J.L.; Huang, M.T.; Fu, H.Y.; Gao, S.J. Presence of diverse sugarcane bacilliform viruses infecting sugarcane in China revealed by pairwise sequence comparisons and phylogenetic analysis. Plant Pathol. J. 2019, 35, 41–50. [Google Scholar] [CrossRef]
- Lockhart, B.E.L.; Autrey, L.J.C. Occurrence in sugarcane of a bacilliform virus related serologically to banana streak virus. Plant Dis. 1988, 72, 230–233. [Google Scholar] [CrossRef]
- Lockhart, B.E.L.; Irey, M.J.; Comstock, J.C. Sugarcane bacilliform virus, Sugarcane mild mosaic virus, and sugarcane yellow leaf syndrome. In Sugarcane Germplasm Conservation and Exchange; Croft, B.J., Piggin, C.T., Wallis, E.S., Hogarth, D.M., Eds.; Australian Centre for International Agricultural Research (ACIAR): Brisbane, Australia, 1996; pp. 108–112. [Google Scholar]
- Bouhida, M.; Lockhart, B.E.; Olszewski, N.E. An analysis of the complete sequence of a sugarcane bacilliform virus genome infectious to banana and rice. J. Gen. Virol. 1993, 74, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Balan, S.; Nithya, K.; Cherian, K.A.; Viswanathan, R. True seed transmission of Sugarcane bacilliform virus (SCBV) in sugarcane. Sugar Tech 2022, 24, 513–521. [Google Scholar] [CrossRef]
- Braithwaite, K.S.; Egeskov, N.M.; Smith, G.R. Detection of Sugarcane bacilliform virus using the polymerase chain reaction. Plant Dis. 1995, 79, 792–796. [Google Scholar] [CrossRef]
- Geijskes, R.J.; Braithwaite, K.S.; Dale, J.L. Sequence analysis of an Australian isolate of sugarcane bacilliform badnavirus. Arch. Virol. 2002, 147, 2393–2404. [Google Scholar] [CrossRef]
- Muller, E.; Dupuy, V.; Blondin, L.; Bauffe, F.; Daugrois, J.H.; Nathalie, L.; Iskra-Caruana, M.L. High molecular variability of sugarcane bacilliform viruses in Guadeloupe implying the existence of at least three new species. Virus Res. 2011, 160, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Karuppaiah, R.; Viswanathan, R.; Kumar, V.G. Genetic diversity of Sugarcane bacilliform virus isolates infecting Saccharum spp. in India. Virus Genes 2013, 46, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.R.; Damaj, M.B.; Alabi, O.J.; Wu, X.B.; Erik Mirkov, T.; Fu, H.Y.; Chen, R.K.; Gao, S.J. Molecular characterization of two divergent variants of sugarcane bacilliform viruses infecting sugarcane in China. Eur. J. Plant Pathol. 2016, 145, 375–384. [Google Scholar] [CrossRef]
- Janiga, P.K.; Nithya, K.; Viswanathan, R. Dynamics of Genetic Diversity Among Indian Sugarcane Bacilliform Virus Species and Implications of Associated Recombination Events in the Virus. Sugar Tech 2022. [Google Scholar] [CrossRef]
- Haregu, S.; Kidanemariam, D.; Abraham, A. Molecular identification and characterization of badnaviruses infecting sugarcane in Ethiopia. Acta Virol. 2022, 66, 3–10. [Google Scholar] [CrossRef]
- Rao, G.P.; Sharma, S.K.; Singh, D.; Arya, M.; Singh, P.; Baranwal, V.K. Genetically diverse variants of Sugarcane bacilliform virus infecting sugarcane in India and evidence of a novel recombinant Badnavirus variant. J. Phytopathol. 2014, 162, 779–787. [Google Scholar] [CrossRef]
- Geijskes, R.J.; Braithwaite, K.S.; Smith, G.R.; Dale, J.L.; Harding, R.M. Sugarcane bacilliform virus encapsidates genome concatamers and does not appear to integrate into the Saccharum officinarum genome. Arch. Virol. 2004, 149, 791–798. [Google Scholar] [CrossRef]
- Cai, Y.; Xu, D.; Zhou, G. Evidence of sugarcane bacilliform virus DNA fragment integrated into the saccharum inter-specific hybrids genome. J. South China Agric. Univ. 2009, 30, 19–23. [Google Scholar]
- Schenk, P.M.; Sagi, L.; Remans, T.; Dietzgen, R.G.; Bernard, M.J.; Graham, M.W.; Manners, J.M. A promoter from sugarcane bacilliform badnavirus drives transgene expression in banana and other monocot and dicot plants. Plant Mol. Biol. 1999, 39, 1221–1230. [Google Scholar] [CrossRef]
- Braithwaite, K.S.; Geijskes, R.J.; Smith, G.R. A variable region of the sugarcane bacilliform virus (SCBV) genome can be used to generate promoters for transgene expression in sugarcane. Plant Cell Rep. 2004, 23, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A.; Feng, X.; Hu, X.; Ashraf, F.; Shen, L.; Iqbal, M.S.; Ashraf, F.; Shen, L.; Iqbal, M.S.; Zhang, S. In silico identification of sugarcane (Saccharum officinarum L.) genome encoded microRNAs targeting sugarcane bacilliform virus. PLoS ONE 2022, 17, e0261807. [Google Scholar] [CrossRef] [PubMed]
- Kreuze, J.F.; Perez, A.; Untiveros, M.; Quispe, D.; Fuentes, S.; Barker, I.; Simon, R. Complete viral genome sequence and discovery of novel viruses by deep sequencing of small RNAs: A generic method for diagnosis, discovery and sequencing of viruses. Virology 2009, 388, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kashif, M.; Pietilä, S.; Artola, K.; Jones, R.A.C.; Tugume, A.K.; Mäkinen, V.; Valkonen, J.P.T. Detection of viruses in sweetpotato from Honduras and Guatemala augmented by deep-sequencing of small-RNAs. Plant Dis. 2012, 96, 1430–1437. [Google Scholar] [CrossRef]
- Mbanzibwa, D.R.; Tugume, A.K.; Chiunga, E.; Mark, D.; Tairo, F.D. Small RNA deep sequencing-based detection and further evidence of DNA viruses infecting sweetpotato plants in Tanzania. Ann. Appl. Biol. 2014, 165, 329–339. [Google Scholar] [CrossRef]
- Buko, D.; Spetz, C.; Hvoslef-Eide, T. Next generation sequencing as a method to verify virus elimination using heat treatment and meristem tip culture in the five most widely used sweet potato varieties in Ethiopia. Afr. J. Biotechnol. 2020, 19, 458–463. [Google Scholar]
- Kreuze, J.F.; Perez, A.; Gargurevich, M.G.; Cuellar, W.J. Badnaviruses of sweet potato: Symptomless coinhabitants on a global scale. Front Plant Sci. 2020, 11, 313. [Google Scholar] [CrossRef] [PubMed]
- Devitt, L.; Ebenebe, A.; Gregory, H.; Harding, R.; Hunter, D.; Macanawai, A. Investigations into the seed and mealybug transmission of Taro bacilliform virus. Aust. Plant Pathol. 2005, 34, 73–76. [Google Scholar]
- Yang, I.C.; Hafner, G.J.; Dale, J.L.; Harding, R.M. Genomic characterisation of taro bacilliform virus. Arch. Virol. 2003, 148, 937–949. [Google Scholar] [CrossRef]
- Yang, I.C.; Hafner, G.J.; Revill, P.A.; Dale, J.L.; Harding, R.M. Sequence diversity of South Pacific isolates of Taro bacilliform virus and the development of a PCR-based diagnostic test. Arch. Virol. 2003, 148, 1957–1966. [Google Scholar] [CrossRef]
- Kazmi, S.A.; Yang, Z.; Hong, N.; Wang, G.; Wang, Y. Characterization by small RNA sequencing of Taro Bacilliform CH Virus (TaBCHV), a novel Badnavirus. PLoS ONE 2015, 10, e0134147. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.C.; Iommarini, J.P.; Becker, D.K.; Hafner, G.J.; Dale, J.L.; Harding, R.M. A promoter derived from taro bacilliform badnavirus drives strong expression in transgenic banana and tobacco plants. Plant Cell Rep. 2003, 21, 1199–1206. [Google Scholar] [CrossRef]
- Li, Y.; Xu, C.; Chen, J. Establishment of virus-free Taro (Colocasia esulenta cv. Fenghuayunaitou) by meristem-tip culture combined with thermotherapy. Pak. J. Plant Pathol. 2002, 1, 40–43. [Google Scholar] [CrossRef]
- Zhao, J.-P.; Jiang, X.-M.; Bai, X.-F.; Zhang, P.; Yong, J.-P. Several Physiological Factors Influencing Virus-free and Rapid Propagation in Shoot-tip Culture of Colocasia esculenta L. Schott. Plant Phisiol. J. 2009, 45, 133–136. [Google Scholar]
- Stace-Smith, R.; Jones, A.T. Virus Diseases of Small Fruits; Converse, R.H., Ed.; United States Department of Agriculture: Washington, DC, USA, 1987; Section 4; p. 175. [Google Scholar]
- Marais, A.; Murolo, S.; Faure, C.; Brans, Y.; Larue, C.; Maclot, F.; Massart, S.; Chiumenti, M.; Minafra, A.; Romanazzi, G.; et al. Sixty Years from the First Disease Description, a Novel Badnavirus Associated with Chestnut Mosaic Disease. Phytopathology 2021, 111, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Rumbou, A.; Candresse, T.; Marais, A.; Theil, S.; Langer, J.; Jalkanen, R.; Büttner, C. A novel badnavirus discovered from Betula sp. affected by birch leaf-roll disease. PLoS ONE 2018, 13, e0193888. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cheng, X.; Wu, X.; Wang, A.; Wu, X. Characterization of complete genome and small RNA profile of Pagoda yellow mosaic associated virus, a novel Badnavirus in China. Virus Res. 2014, 188, 103–108. [Google Scholar] [CrossRef]
- Chiumenti, M.; Morelli, M.; Elbeaino, T.; Stavolone, L.; De Stradis, A.; Digiaro, M.; Minafra, A.; Martelli, G.P. Sequencing an unconventional virus genome: The mulberry badnavirus 1 case. J. Plant Pathol. 2014, 96, 36–37. [Google Scholar]
- Alvarez-Quinto, R.A.; Lockhart, B.E.L.; Moreno-Martinez, J.M.; Olszewski, N.E. Complete genome sequence of aglaonema bacilliform virus (ABV). Arch. Virol. 2020, 165, 237–239. [Google Scholar] [CrossRef]
- Uke, A.; Pinili, M.S.; Natsuaki, K.T.; Geering, A.D.W. Complete genome sequence of aucuba ringspot virus. Arch. Virol. 2021, 166, 1227–1230. [Google Scholar] [CrossRef]
- Tsai, C.H.; Su, H.J.; Wu, M.-L.; Feng, Y.C.; Hung, T.H. Identification and detection of Bougainvillea spectabilis chlorotic vein-banding virus in different bougainvillea cultivars in Taiwan. Ann. Appl. Biol. 2008, 53, 187–193. [Google Scholar] [CrossRef]
- Baranwal, V.K.; Meenakshi, A.; Singh, J. First report of two distinct badnaviruses associated with Bougainvillea spectabilis in India. J. Gen. Plant Pathol. 2010, 76, 236–239. [Google Scholar] [CrossRef]
- Yusop, M.S.M.; Ersoy, R.; Akbar, M.A.; Saad, M.F.M.; Goh, H.H.; Baharum, S.N.; Bunawan, H. First report of Bougainvillea chlorotic vein-banding virus causing Chlorosis and vein banding of Bougainvillea spectabilis in Malaysia. Plant Dis. 2019, 103, 2974. [Google Scholar] [CrossRef]
- Naito, F.Y.B.; De Nazaré Almeida dos Reis, L.; Batista, J.G.; Nery, F.M.B.; Rossato, M.; Melo, F.L.; De Cássia Pereira-Carvalho, R. Complete genome sequence of Bougainvillea chlorotic vein banding virus in Bougainvillea spectabilis from Brazil. Trop. Plant Pathol. 2020, 45, 159–162. [Google Scholar] [CrossRef]
- Li, R.; Zheng, L.; Cao, M.; Wu, L.; Normandy, P.; Liu, H. First identification and molecular characterization of a new badnavirus infecting camellia. Arch. Virol. 2020, 165, 2115–2118. [Google Scholar] [CrossRef] [PubMed]
- Wijayasekara, D.; Hoyt, P.; Gimondo, A.; Dunn, B.; Thapa, A.; Jones, H.; Verchot, J. Molecular characterization of two badnavirus genomes associated with Canna yellow mottle disease. Virus Res. 2018, 2, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Dey, K.K.; Lin, B.; Borth, W.B.; Melzer, M.J.; Sether, D.; Wang, Y.; Wang, I.C.; Shen, H.; Pu, X.; et al. Characterization of Canna yellow mottle virus in a New Host, Alpinia purpurata, in Hawaii. Phytopathology 2017, 107, 791–799. [Google Scholar] [CrossRef]
- Lim, S.; Park, J.M.; Kwon, S.Y.; Cho, H.S.; Kim, H.S.; Lee, S.H.; Moon, J.S. Complete genome sequence of a tentative new member of the genus Badnavirus identified in Codonopsis lanceolata. Arch. Virol. 2019, 164, 1733–1737. [Google Scholar] [CrossRef]
- Su, L.; Gao, S.; Huang, Y.; Ji, C.; Wang, D.; Ma, Y.; Fang, R.; Chen, X. Complete genomic sequence of Dracaena mottle virus, a distinct badnavirus. Virus Genes 2007, 35, 423–430. [Google Scholar] [CrossRef]
- Lan, P.X.; Tian, T.Y.; Pu, L.L.; Rao, W.L.; Li, F.; Li, R.H. Characterization and detection of a new badnavirus infecting Epiphyllum spp. Arch. Virol. 2019, 164, 1837–1841. [Google Scholar] [CrossRef]
- Bester, R.; Burger, J.T.; Maree, H.J. Genomic characterisation of a newly identified badnavirus infecting ivy (Hedera helix). Arch. Virol. 2020, 165, 1511–1514. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Nicotaisen, M.; Olszewski, N.E.; Lockhart, B.E. Sequencing, improved detection, and a novel form of Kalanchoë top-spotting virus. Plant Dis. 2005, 89, 298–302. [Google Scholar] [CrossRef]
- Diaz-Lara, A.; Mosier, N.J.; Keller, K.E.; Martin, R.R. A variant of Rubus yellow net virus with altered genomic organization. Virus Genes 2015, 50, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Quinto, R.A.; Serrano, J.; Olszewski, N.; Grinstead, S.; Mollov, D.; Lockhart, B.E.L. Complete genome sequences of two isolates of spiraea yellow leafspot virus (genus Badnavirus) from Spiraea x bumalda ‘Anthony Waterer’. Arch. Virol. 2022, 167, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Deng, C.; Qiao, Y.; Zhao, X.; Zhou, Q. Characterization of a new badnavirus from Wisteria sinensis. Arch. Virol. 2017, 162, 2125–2129. [Google Scholar] [CrossRef]
- Mehrvar, M.; Moradi, Z.; Al-Jaberi, M.S. First report of Wisteria badnavirus 1 infecting Wisteria sinensisin Iran. New Dis. Rep. 2022, 46, e12133. [Google Scholar] [CrossRef]
- Alvarez-Quinto, R.A.; Lockhart, B.E.L.; Fetzer, J.L.; Olszewski, N.E. Genomic characterization of cycad leaf necrosis virus, the first badnavirus identified in a gymnosperm. Arch Virol. 2020, 165, 1671–1673. [Google Scholar] [CrossRef]
- Shahid, M.S.; Aboughanem-Sabanadzovic, N.; Sabanadzovic, S.; Tzanetakis, I.E. Genomic Characterization and Population Structure of a Badnavirus Infecting Blackberry. Plant Dis. 2017, 101, 110–115. [Google Scholar] [CrossRef]
- Laney, A.G.; Hassan, M.; Tzanetakis, I.E. An integrated badnavirus is prevalent in fig germplasm. Phytopathology 2012, 102, 1182–1189. [Google Scholar] [CrossRef] [Green Version]
- Alishiri, A.; Rakhshandehroo, F.; Jouzani, G.S.; Shams-bakhsh, M. Exploring the genetic diversity and molecular evolution of fig badnavirus-1 from Iran. J. Plant Pathol. 2018, 100, 287–299. [Google Scholar] [CrossRef]
- Xu, D.; Mock, R.; Kinard, G.; Li, R. Molecular analysis of the complete genomic sequences of four isolates of Gooseberry vein banding associated virus. Virus Genes 2011, 43, 130–137. [Google Scholar] [CrossRef]
- Petrzik, K.; Přibylová, J.; Spak, J. Molecular analysis of gooseberry vein banding associated virus. Acta Virol. 2012, 56, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Zuļģe, N.; Stalažs, A.; Moročko-Bičevska, I.; Namniece, S.; Drevinska, K.; Konavko, D. Gooseberry vein banding associated virus on Ribes in Latvia: Occurrence, host plants and vectors. Plant Pathol. 2022, 71, 1910–1920. [Google Scholar] [CrossRef]
- Du, K.; Liu, S.; Chen, Z.; Fan, Z.; Wang, H.; Tian, G.; Zhou, T. Full genome sequence of jujube mosaic-associated virus, a new member of the family Caulimoviridae. Arch. Virol. 2017, 162, 3221–3224. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, G.; Song, D.; Wang, Q.; Li, H.; Gu, A.; Bai, J. Complete genome sequence of a novel virus belonging to the genus Badnavirus in jujube (Ziziphus jujuba Mill.) in China. Arch. Virol. 2022, 167, 1885–1888. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Quinto, R.A.; Lockhart, B.E.L.; Olszewski, N. Complete genome sequence of a previously undescribed badnavirus occurring in Polyscias fruticosa L. (Ming aralia). Arch Virol. 2019, 164, 2371–2374. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, S.; Xuan, Z.; Wu, J.; Dong, P.; Zhou, Y.; Li, R.; Cao, M. Molecular characterization of a new badnavirus infecting green Sichuan pepper (Zanthoxylum schinifolium). Arch. Virol. 2019, 164, 2613–2616. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Kwak, H.R.; Lee, Y.K.; Kim, M.K.; Choi, H.S.; Seo, J.K. Complete genome sequence of yacon necrotic mottle virus, a novel putative member of the genus Badnavirus. Arch. Virol. 2015, 160, 1139–1142. [Google Scholar] [CrossRef]
- Medberry, S.L.; Lockhart, B.E.L.; Olszewski, N.E. The Commelinayellow m ottlevirus promoter is a strong promoter in vascular and reproductive tissues. Plant Cell 1992, 4, 185–192. [Google Scholar]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No | Species | Host | Acronym | GenBank Accession Number | No. of ORFs | Genome Length (Nucleotide) |
|---|---|---|---|---|---|---|
| 1. | aglaonema bacilliform virus | Aglaonema modestum | ABV | MH384837 | 3 | 7178 |
| 2. | Badnavirus aucubae (aucuba ringspot virus) | Aucuba japonica | AuRV | LC487411 | 3 | 9092 |
| 3. | Badnavirus castaneae (chestnut mosaic virus) | Castanea sativa | ChMD | MT269853 | 3 | 7160 |
| 4. | banana streak GF virus | Plantain | BSGFV | AY493509 | 3 | 7263 |
| 5. | banana streak IM virus | Musa sp. cv. Mshule | BSIMV | HQ593112 | 3 | 7769 |
| 6. | banana streak MY virus | Musa sp. | BSMYV | AY805074 | 3 | 7650 |
| 7. | banana streak OL virus | Musa sp. | BSOLV | AJ002234 | 3 | 7389 |
| 8. | banana streak UA virus | Musa sp. cv. Likhako | BSUAV | HQ593107 | 3 | 7519 |
| 9. | banana streak UI virus | Musa sp. cv. Kisansa | BAUIV | HQ593108 | 3 | 7458 |
| 10. | banana streak UL virus | Musa sp. cv. Kibuzi | BSULV | HQ593109 | 3 | 7401 |
| 11. | banana streak UM virus | Musa sp. cv. Mbwazirume | BSUMV | HQ593110 | 3 | 7532 |
| 12. | banana streak VN virus | Plantain | BAVNV | AY750155 | 3 | 7801 |
| 13. | birch leaf roll-associated virus | Betula pubescens | BLRaV | MG686419 | 4 | 7864 |
| 14. | blackberry virus F | Rubus sp. | BVF | KJ413252 | 3 | 7663 |
| 15. | bougainvillea chlorotic vein banding virus | Bougainvillea sp. | BCVBV | EU034539 | 4 | 8759 |
| 16. | cacao bacilliform Sri Lanka virus | Theobroma cacao | CBSLV | MF642736 | 3 | 1772 |
| 17. | cacao mild mosaic virus | Theobroma cacao | CaMMV | KX276640 | 4 | 7533 |
| 18. | cacao swollen shoot CD virus | Theobroma cacao | CSSCDV | JN606110 | 4 | 7203 |
| 19. | cacao swollen shoot CE virus | Theobroma cacao | CSSCEV | MF642720 | 4 | 7131 |
| 20. | cacao swollen shoot Ghana M virus | Theobroma cacao | CSSGMV | MF642724 | 4 | 7009 |
| 21. | cacao swollen shoot Ghana N virus | Theobroma cacao | CSSGNV | MF642725 | 4 | 7173 |
| 22. | cacao swollen shoot Ghana Q virus | Theobroma cacao | CSSGQV | MF642734 | 4 | 7102 |
| 23. | cacao swollen shoot Togo A virus | Theobroma cacao | CSSTAV | AJ781003 | 9 | 7297 |
| 24. | cacao swollen shoot Togo B virus | Theobroma cacao | CSSTBV | L14546 | 3 | 7161 |
| 25. | cacao yellow vein banding virus | Theobroma cacao | CYVBV | KX276641 | 3 | 1958 |
| 26. | camellia lemon glow virus | Camellia japonica cv. Lemon Glow | CLGV | MN542417 | 3 | 8203 |
| 27. | canna yellow mottle associated virus | Canna indica cv. Striped Beauty leaf | CaYMAV | KX066020 | 3 | 6966 |
| 28. | canna yellow mottle virus | Alpinia purpurata | CaYMV | KU168312 | 3 | 7120 |
| 29. | citrus yellow mosaic virus | Citrussinensis | CYMV | AF347695 | 6 | 7559 |
| 30. | codonopsis vein clearing virus | Codonopsis lanceolata | CoVCV | MK044821 | 3 | 8112 |
| 31. | commelina yellow mottle virus | Commelina sp. | ComYMV | X52938 | 3 | 7489 |
| 32. | cycad leaf necrosis | Zamia fischeri | CLNV | EU853709 | 3 | 9205 |
| 33. | dioscorea bacilliform AL virus | Dioscorea rotundata | DBALV | KX008573 | 3 | 7609 |
| 34. | dioscorea bacilliform AL virus 2 | Dioscorea alata | DBALV2 | KY827395 | 3 | 7871 |
| 35. | dioscorea bacilliform ES virus | Dioscorea esculenta | DBESV | KY827394 | 3 | 8106 |
| 36. | dioscorea bacilliform RT virus 1 | Dioscorea rotundata | DBRTV1 | KX008574 | 3 | 7702 |
| 37. | dioscorea bacilliform RT virus 2 | Dioscorea rotundata | DBRTV2 | KX008577 | 3 | 7438 |
| 38. | dioscorea bacilliform RT virus 3 | Dioscorea rotundata | DBRTV3 | MF476845 | 3 | 7506 |
| 39. | dioscorea bacilliform SN virus | Dioscorea sansibarensis | DBSNV | DQ822073 | 3 | 7261 |
| 40. | dioscorea bacilliform TR virus | Dioscorea trifida var. Borelli voucher | DBTRV | KX430257 | 3 | 7333 |
| 41. | dracaena mottle virus | Dracaena sanderiana | DrMV | DQ473478 | 7 | 7531 |
| 42. | epiphyllum mottle-associated virus | Hylocereus polyrhizus | EpMoaV | MH396440 | 4 | 7837 |
| 43. | fig badnavirus 1 | Ficus carica | FBV -1 | JF411989 | 3 | 7140 |
| 44. | gooseberry vein banding associated virus | Ribes rubrum cv. Holandsky cerveny | GVBAV | JQ316114 | 3 | 7659 |
| 45. | grapevine badnavirus 1 | Vitis vinifera var. Ljutun | GBV-1 | MF781082 | 3 | 7145 |
| 46. | grapevine Roditis leaf discoloration-associated virus | Vitis vinifera | GRLDaV | HG940503 | 4 | 6988 |
| 47. | grapevine vein clearing virus | Vitis vinifera | GVCV | JF301669 | 3 | 7753 |
| 48. | green Sichuan pepper vein clearing-associated virus | Zanthoxylum | GSPVCaV | MK371353 | 3 | 8070 |
| 49. | ivy ringspot-associated virus | Hedera helix | IRSaV | MN850490 | 3 | 8885 |
| 50. | jujube mosaic-associated virus | Ziziphus jujube | JuMaV | KX852476 | 4 | 7194 |
| 51. | kalanchoe top-spotting virus | Kalanchoe blossfeldiana | KTSV | AY180137 | 3 | 7591 |
| 52. | mulberry badnavirus 1 | Morus alba | MBV-1 | LN651258 | 2 | 6945 |
| 53. | pagoda yellow mosaic associated virus | Styphnolobium japonicum | PYMaV | KJ013302 | 5 | 7424 |
| 54. | pineapple bacilliform CO virus | Ananas comosus | PBCOV | GU121676 | 3 | 7543 |
| 55. | pineapple bacilliform ER virus | Ananas comosus var. erectifolius | PBERV | EU377673 | 3 | 1510 |
| 56. | piper yellow mottle virus | Piper nigrum | PYMoV | KC808712 | 4 | 7562 |
| 57. | polyscias mosaic virus | Polyscias fruticosa cv. Ming aralia | PoMV | MH475918 | 3 | 7592 |
| 58. | rubus yellow net virus | Rubus sp. | RYNV | KM078034 | 6 | 7836 |
| 59. | spiraea yellow leafspot virus | Spiraea (Spiraea × bumalda) | SYLSV | MW080369 | 3 | 8017 |
| 60. | sugarcane bacilliform Guadeloupe A virus | Saccharum officinarum var. R570 | SCBGAV | FJ824813 | 3 | 7444 |
| 61. | sugarcane bacilliform Guadeloupe D virus | Saccharum officinarum cv.Batavia | SCBGDV | FJ439817 | 3 | 7317 |
| 62. | sugarcane bacilliform IM virus | Saccharum officinarum | SCBIMV | AJ277091 | 3 | 7687 |
| 63. | sugarcane bacilliform MO virus | Saccharum officinarum | SCBMOV | M89923 | 3 | 7568 |
| 64. | sweet potato pakakuy virus | Ipomoea batatas | SPPV | FJ560943 | 4 | 8082 |
| 65. | taro bacilliform CH virus | Colocasia esculenta | TaBCHV | KP710178 | 6 | 7641 |
| 66. | taro bacilliform virus | Colocasia esculenta | TaBV | AF357836 | 3 | 7458 |
| 67. | wisteria badnavirus 1 | Wisteria sinensis | WBV1 | KX168422 | 4 | 7326 |
| 68. | yacon necrotic mottle virus | Smallanthus sonchifolius | YNMoV | KM229702 | 4 | 7661 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishwara Bhat, A.; Selvarajan, R.; Balasubramanian, V. Emerging and Re-Emerging Diseases Caused by Badnaviruses. Pathogens 2023, 12, 245. https://doi.org/10.3390/pathogens12020245
Ishwara Bhat A, Selvarajan R, Balasubramanian V. Emerging and Re-Emerging Diseases Caused by Badnaviruses. Pathogens. 2023; 12(2):245. https://doi.org/10.3390/pathogens12020245
Chicago/Turabian StyleIshwara Bhat, Alangar, Ramasamy Selvarajan, and Velusamy Balasubramanian. 2023. "Emerging and Re-Emerging Diseases Caused by Badnaviruses" Pathogens 12, no. 2: 245. https://doi.org/10.3390/pathogens12020245