
Establishment of a Real-Time Recombinase Polymerase Amplification for Rapid Detection of Pathogenic Yersinia enterocolitica

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Condition
2.2. Extraction of Genomic DNA
2.3. RPA Primer and Probe Designs
2.4. Real-Time RPA Assay
2.5. Analytical Sensitivity and Specificity
2.6. Repeatability Testing
2.7. Validation with Artificially Contaminated Samples
2.8. Data Analysis
3. Results
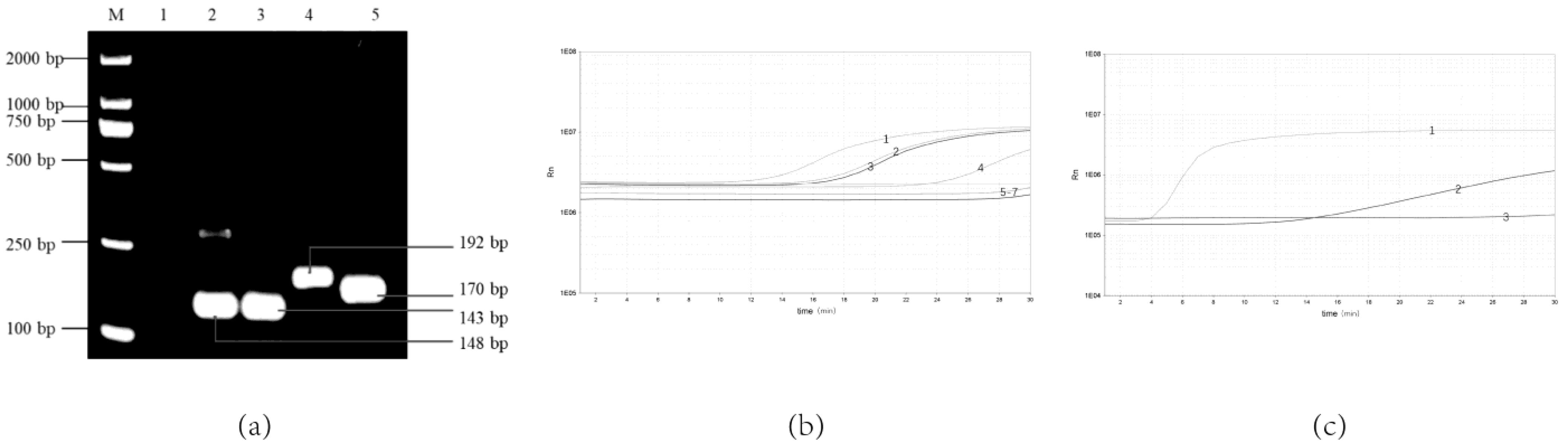
3.1. Optimal Combination Primers and Probes for Real-Time RPA Assay
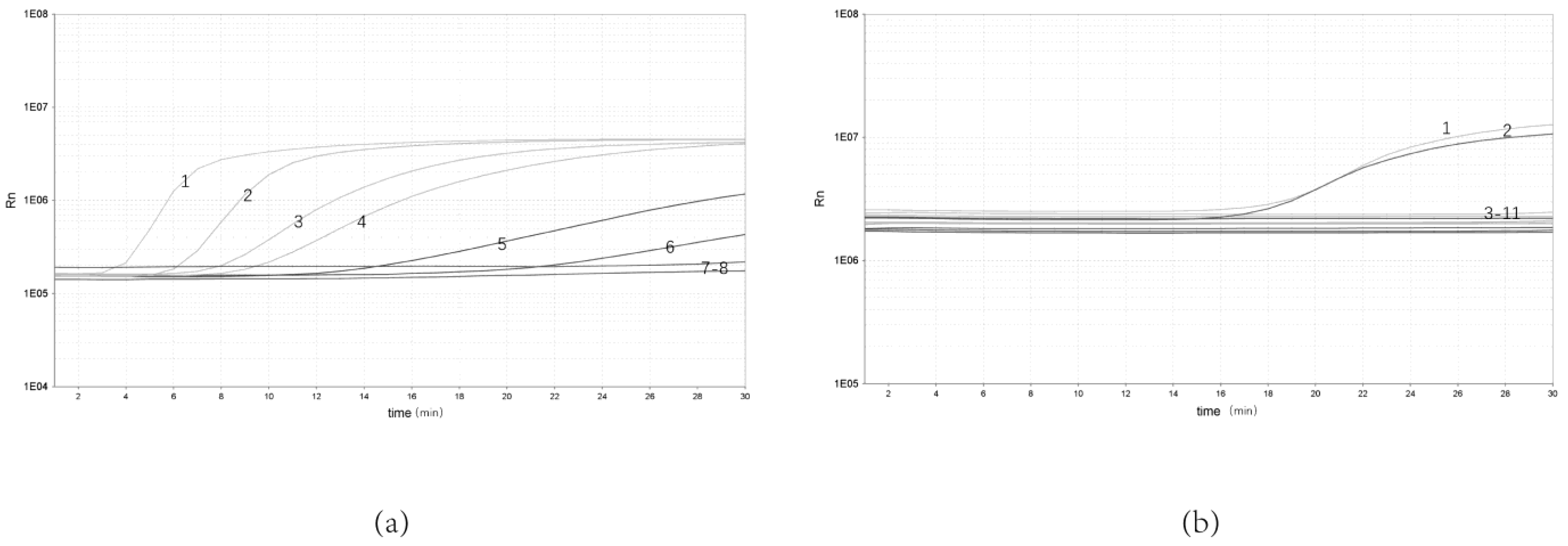
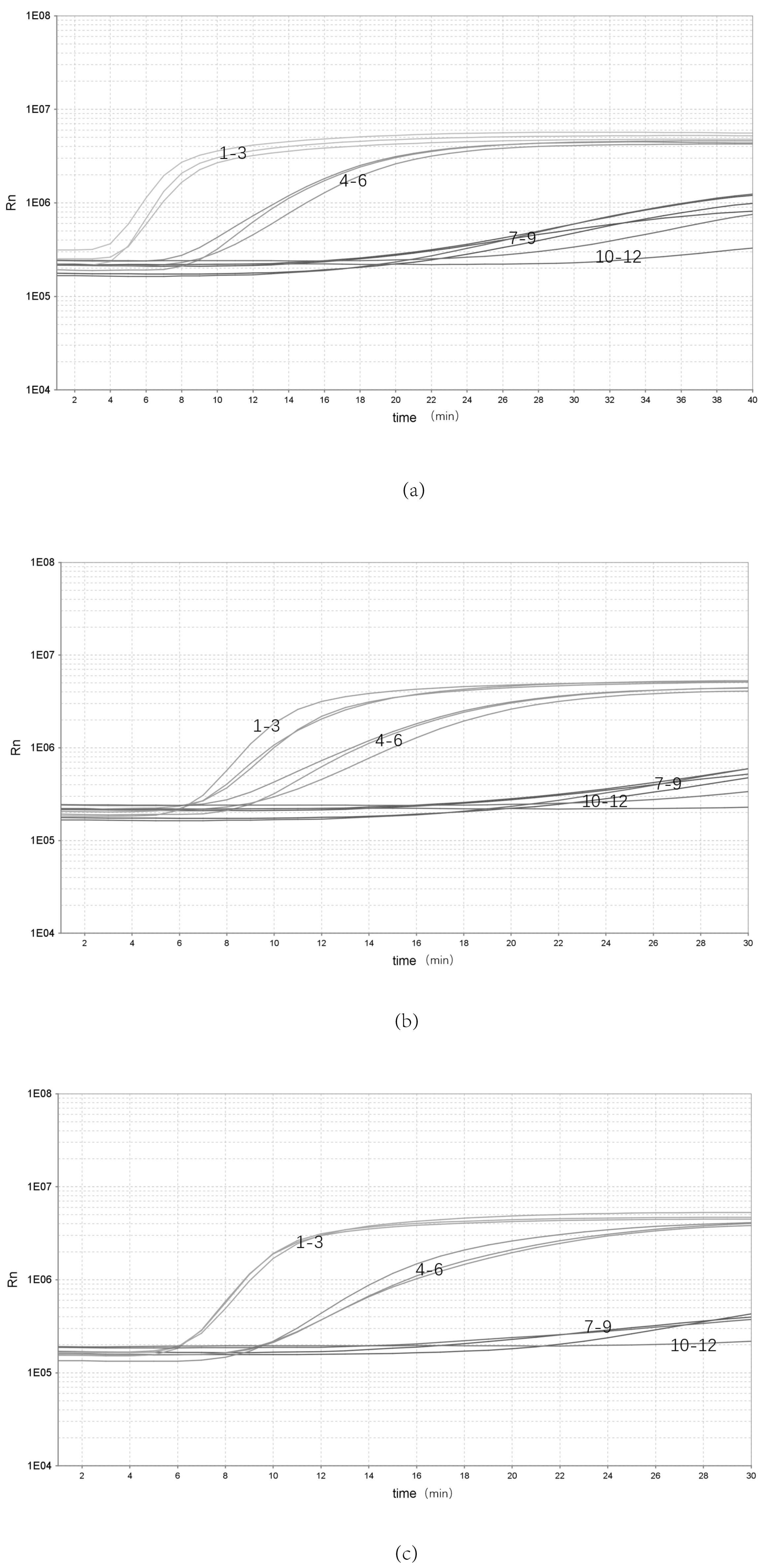
3.2. Analytical Sensitivity and Specificity of the Real-Time RPA Assay
3.3. Analytical Repeatability of the Real-Time RPA Assay
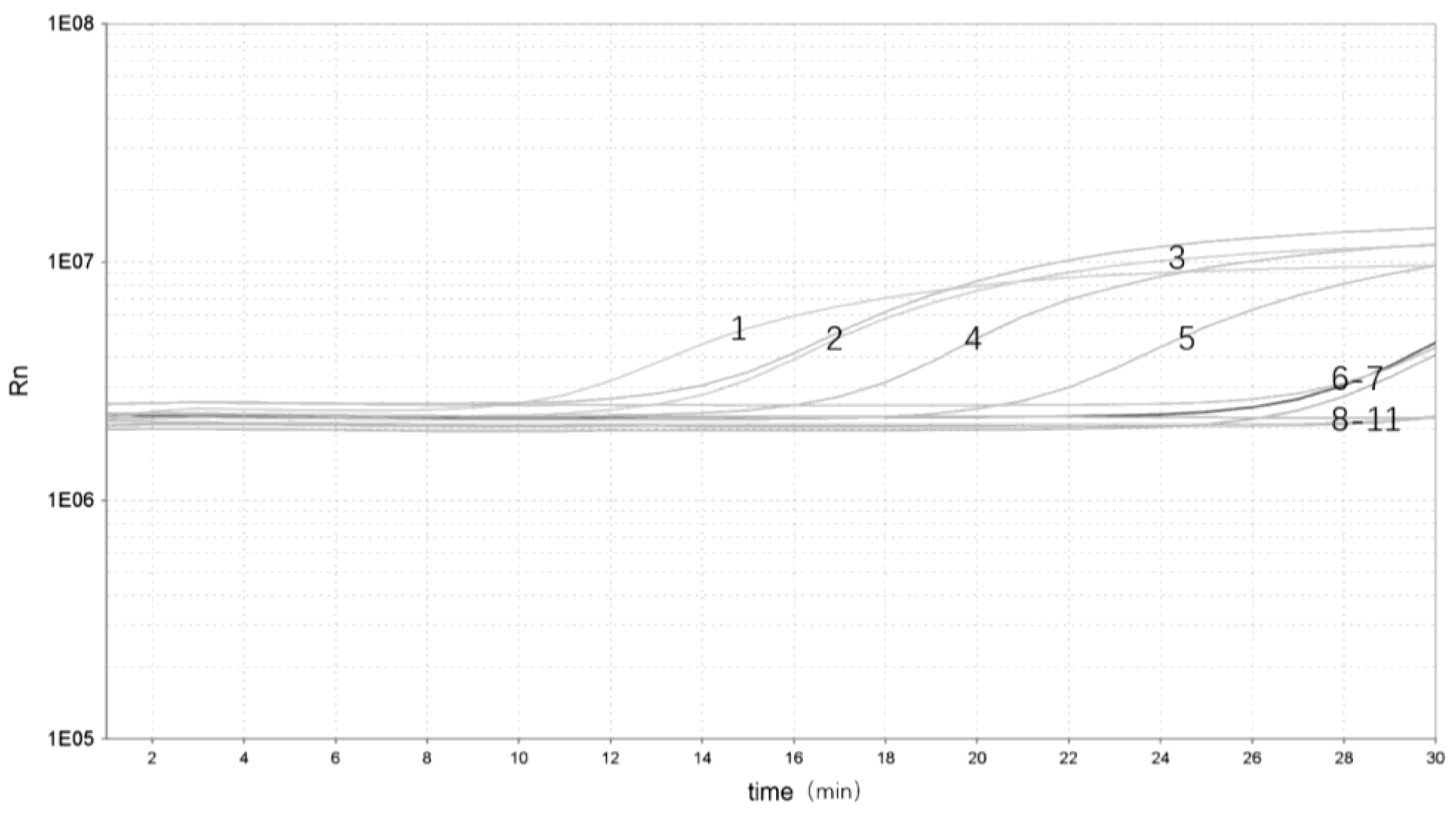
3.4. Validation of the Real-Time RPA Assay on Artificially Contaminated Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bottone, E.J. Yersinia enterocolitica: Revisitation of an Enduring Human Pathogen. Clin. Microbiol. Newsl. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- Rusak, L.A.; Pereira, R.D.C.L.; Freitag, I.G.; Hofer, C.B.; Hofer, E.; Asensi, M.D.; Vallim, D.C. Rapid detection of Yersinia enterocolitica serotype O:3 using a duplex PCR assay. J. Microbiol. Methods 2018, 154, 107–111. [Google Scholar] [CrossRef]
- Galindo, C.L.; Rosenzweig, J.A.; Kirtley, M.L.; Chopra, A.K. Pathogenesis of Y. enterocolitica and Y. pseudotuberculosis in Human Yersiniosis. J. Pathog. 2011, 2011, 182051. [Google Scholar]
- Stachelska, M. Detection of Yersinia Enterocolitica Species in Pig Tonsils and Raw Pork Meat by the Real-Time Pcr and Culture Methods. Pol. J. Veter. Sci. 2017, 20, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Gulati, P.; Bhagat, N.; Dhar, M.S.; Virdi, J.S. Detection of Yersinia enterocolitica in food: An overview. Eur. J. Clin. Microbiol. 2015, 34, 641. [Google Scholar] [CrossRef]
- Platt-Samoraj, A.; Syczyło, K.; Szczerba-Turek, A.; Bancerz-Kisiel, A.; Jabłoński, A.; Łabuć, S.; Pajdak, J.; Oshakbaeva, N.; Szweda, W. Presence of ail and ystB genes in Yersinia enterocolitica biotype 1A isolates from game animals in Poland. Veter. J. 2017, 221, 11–13. [Google Scholar] [CrossRef]
- Kraushaar, B.; Dieckmann, R.; Wittwer, M.; Knabner, D.; Konietzny, A.; Mäde, D.; Strauch, E. Characterization of a Yersinia enterocolitica biotype 1A strain harbouring an ail gene. J. Appl. Microbiol. 2011, 111, 997–1005. [Google Scholar] [CrossRef]
- Bui, T.H.; Ikeuchi, S.; O’Brien, Y.S.; Niwa, T.; Hara-Kudo, Y.; Taniguchi, T.; Hayashidani, H. Multiplex PCR method for differentiating highly pathogenic Yersinia enterocolitica and low pathogenic Yersinia enterocolitica, and Yersinia pseudotuberculosis. J. Veter. Med. Sci. 2021, 83, 1982–1987. [Google Scholar] [CrossRef]
- Rohde, A.; Hammerl, J.A.; Appel, B.; Dieckmann, R.; Al Dahouk, S. Differential detection of pathogenic Yersinia spp. by fluorescence in situ hybridization. Food Microbiol. 2017, 62, 39–45. [Google Scholar] [CrossRef]
- Tack, D.M.; Marder, E.; Griffin, P.; Cieslak, P.; Dunn, J.; Hurd, S.; Scallan, E.; Lathrop, S.; Muse, A.; Ryan, P.; et al. Preliminary Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food-Foodborne Diseases Active Surveillance Network, 10 US Sites, 2015–2018. Mmwr-Morb. Mortal. Wkly. Rep. 2019, 68, 369–373. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, M.; Wang, H.; Wu, Q.; Ding, Y.; Xu, T.; Ma, G.; Zhong, Y.; Zhang, J.; Chen, M.; et al. Occurrence, molecular characterization, and antimicrobial susceptibility of Yersinia enterocolitica isolated from retail food samples in China. LWT 2021, 150, 111876. [Google Scholar] [CrossRef]
- Aldová, E.; Švandová, E.; Votýpka, J.; Šourek, J. Comparative Study of Culture Methods to Detect Yersinia enterocolitica Serogroup O3 on Swine Tongues. Zent. Bakteriol. Int. J. Med. Microbiol. 1990, 272, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Kwaga, J.; Iversen, J.O.; Misra, V. Detection of pathogenic Yersinia enterocolitica by polymerase chain reaction and digoxigenin-labeled polynucleotide probes. J. Clin. Microbiol. 1992, 30, 2668–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambertz, S.T.; Nilsson, C.; Hallanvuo, S.; Lindblad, M. Real-Time PCR Method for Detection of Pathogenic Yersinia enterocolitica in Food. Appl. Environ. Microbiol. 2008, 74, 6060–6067. [Google Scholar] [CrossRef] [Green Version]
- Ranjbar, R.; Afshar, D. Development of a loop-mediated isothermal amplification assay for rapid detection of Yersinia enterocolitica via targeting a conserved locus. Iran. J. Microbiol. 2015, 7, 185–190. [Google Scholar] [PubMed]
- Luciani, M.; Schirone, M.; Portanti, O.; Visciano, P.; Armillotta, G.; Tofalo, R.; Suzzi, G.; Sonsini, L.; Di Febo, T. Development of a rapid method for the detection of Yersinia enterocolitica serotype O:8 from food. Food Microbiol. 2018, 73, 85–92. [Google Scholar] [CrossRef]
- Mullis, K.; Faloona, F.; Scharf, S.; Saiki, R.; Horn, G.; Erlich, H. Specific enzymatic amplification of DNA in vitro: The polymerase chain reaction. Cold Spring Harb. Symp. Quant. Biol. 1986, 51 Pt 1, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.; Ma, J.; Li, X.; Wang, R.; Chen, S. A real-time recombinase polymerase amplification assay for fast and accurate detection of Ditylenchus destructor. Mol. Cell. Probes 2021, 61, 101788. [Google Scholar] [CrossRef]
- Moore, M.D.; Jaykus, L.-A. Development of a Recombinase Polymerase Amplification Assay for Detection of Epidemic Human Noroviruses. Sci. Rep. 2017, 7, 40244. [Google Scholar] [CrossRef]
- Prescott, M.A.; Reed, A.N.; Jin, L.; Pastey, M.K. Rapid Detection of Cyprinid Herpesvirus 3 in Latently Infected Koi by Recombinase Polymerase Amplification. J. Aquat. Anim. Health 2016, 28, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase polymerase amplification: Basics, applications and recent advances. TrAC Trends Anal. Chem. 2017, 98, 19–35. [Google Scholar] [CrossRef]
- Frimpong, M.; Kyei-Tuffuor, L.; Fondjo, L.A.; Ahor, H.S.; Adjei-Kusi, P.; Maiga-Ascofare, O.; Phillips, R.O. Evaluation of a real-time recombinase polymerase amplification assay for rapid detection of Schistosoma haematobium infection in resource-limited setting. Acta Trop. 2021, 216, 105847. [Google Scholar] [CrossRef]
- Wang, H.; Hou, P.; Zhao, G.; Yu, L.; Gao, Y.-W.; He, H. Development and evaluation of serotype-specific recombinase polymerase amplification combined with lateral flow dipstick assays for the diagnosis of foot-and-mouth disease virus serotype A, O and Asia1. BMC Veter. Res. 2018, 14, 359. [Google Scholar] [CrossRef]
- Zhao, G.; Hou, P.; Huan, Y.; He, C.; Wang, H.; He, H. Development of a recombinase polymerase amplification combined with a lateral flow dipstick assay for rapid detection of the Mycoplasma bovis. BMC Veter. Res. 2018, 14, 412. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Liu, G.; Liu, L.; Deng, Q.; Zhao, L.; Sun, X.X.; Wang, J.; Zhao, B.; Wang, J. Real-time recombinase polymerase amplification assay for the rapid and sensitive detection of Campylobacter jejuni in food samples. J. Microbiol. Methods 2018, 157, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, X.; Wang, Y.; Shen, H.; Jiang, G.; Dong, J.; Zhao, P.; Gao, S. A Real-Time Recombinase Polymerase Amplification Method for Rapid Detection of Vibrio vulnificus in Seafood. Front. Microbiol. 2020, 11, 586981. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Shi, H.; Zhang, M.; Song, Y.; Zhang, K.; Cong, F. Establishment of a Real-Time Recombinase Polymerase Amplification Assay for the Detection of Avian Reovirus. Front. Veter. Sci. 2020, 7, 551350. [Google Scholar] [CrossRef]
- Laporte, J.; Savin, C.; Lamourette, P.; Devilliers, K.; Volland, H.; Carniel, E.; Créminon, C.; Simon, S. Fast and Sensitive Detection of Enteropathogenic Yersinia by Immunoassays. J. Clin. Microbiol. 2015, 53, 146–159. [Google Scholar] [CrossRef] [Green Version]
- Van Noyen, R.; Vandepitte, J.; Wauters, G. Nonvalue of cold enrichment of stools for isolation of Yersinia enterocolitica serotypes 3 and 9 from patients. J. Clin. Microbiol. 1980, 11, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Blom, M.; Meyer, A.; Gerner-Smidt, P.; Gaarslev, K.; Espersen, F. Evaluation of Statens Serum Institut enteric medium for detection of enteric pathogens. J. Clin. Microbiol. 1999, 37, 2312–2316. [Google Scholar] [CrossRef] [Green Version]
- Petsios, S.; Fredriksson-Ahomaa, M.; Sakkas, H.; Papadopoulou, C. Conventional and molecular methods used in the detection and subtyping of Yersinia enterocolitica in food. Int. J. Food Microbiol. 2016, 237, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Hu, P.; Ren, H.; Wang, H.; Cao, Q.; Zhao, Q.; Li, H.; Zhang, H.; Liu, Z.; Li, Y.; et al. RPA-SYBR Green I based instrument-free visual detection for pathogenic Yersinia enterocolitica in meat. Anal. Biochem. 2021, 621, 114157. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Species | Source | ail Gene |
|---|---|---|---|
| ZDN 6 | Y. enterocolitica | isolated from Yak | + |
| ATCC 23715 | Y. enterocolitica | CIVDC | + |
| 08-01 | Y. enterocolitica | isolated from Landrace | + |
| GN 22 | Y. enterocolitica | isolated from Yak | + |
| ZDN 22 | Y. enterocolitica | isolated from Yak | + |
| GLN 1 | Y. enterocolitica | isolated from Yak | − |
| HNN 18 | Y. enterocolitica | isolated from Yak | − |
| HNN 47 | Y. enterocolitica | isolated from Yak | − |
| ZDN 37 | Y. enterocolitica | isolated from Yak | − |
| HNN 63 | Y. enterocolitica | isolated from Yak | − |
| 06-01-03 | Shigella Castellani | isolated from Yak | − |
| 06-03-01 | Salmonella | isolated from Feedstuff | − |
| 03-03 | Escherichia coli | isolated from Feedstuff | − |
| AH 1302 | Aeromonas hydrophila | isolated from Gymnocypris przewalskii | − |
| 06-05-02 | Bacillus cereus | isolated from Feedstuff | − |
| P0810 | Pasteurella | isolated from Yak | − |
| 04-011 | Staphylococcus aureus | isolated from Yak | − |
| Name | Sequence (5′-3′) |
|---|---|
| RPA-F-1 | TCATGGAAAGGTTAAGGCATCTGTATTTGA |
| RPA-F-2 | ATAGGTTCGTTTGCTTATACTCATCAGGGA |
| RPA-F-3 | AAAGGTTTTAACCTGAAGTACCGTTATGAA |
| RPA-R-1 | TTTTATGCTATCGAGTTTGGAGTATTCATA |
| RPA-R-2 | AGTAATCCATAAAGGCTAACATATTCGTTG |
| RPA-R-3 | TAATCCATAAAGGCTAACATATTCGTTGAT |
| RPA-Probe-5 | AGGTTCGTTTGCTTATACTCATCAGGGATA(FAM-dT) (THF)A(BHQ1-dT) TTCTTCTATGGCAGTA(3′-block) |
| RPA-Probe-6 | GAATAGTAATCAACATCACCATGACCAAACT(FAM-dT) (THF) (BHQ1-dT) TACTGCCATAGAAGA(3′-block) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zhao, M.; Hu, S.; Ma, K.; Li, J.; Zhao, J.; Wei, X.; Tong, L.; Li, S. Establishment of a Real-Time Recombinase Polymerase Amplification for Rapid Detection of Pathogenic Yersinia enterocolitica. Pathogens 2023, 12, 226. https://doi.org/10.3390/pathogens12020226
Zhang H, Zhao M, Hu S, Ma K, Li J, Zhao J, Wei X, Tong L, Li S. Establishment of a Real-Time Recombinase Polymerase Amplification for Rapid Detection of Pathogenic Yersinia enterocolitica. Pathogens. 2023; 12(2):226. https://doi.org/10.3390/pathogens12020226
Chicago/Turabian StyleZhang, Hongjian, Meng Zhao, Siyun Hu, Kairu Ma, Jixu Li, Jing Zhao, Xin Wei, Lina Tong, and Shengqiang Li. 2023. "Establishment of a Real-Time Recombinase Polymerase Amplification for Rapid Detection of Pathogenic Yersinia enterocolitica" Pathogens 12, no. 2: 226. https://doi.org/10.3390/pathogens12020226