
Evaluation of the Biocontrol Efficiency of Bacillus subtilis Wettable Powder on Pepper Root Rot Caused by Fusarium solani

Abstract
:1. Introduction
2. Results
2.1. Antifungal Activity of B. subtilis PTS-394 and Its Wettable Powder
2.2. Identification of Lipopeptide from B. subtilis PTS-394 and Its Wettable Powder
2.3. Growth-Promoting Effect of B. subtilis PTS-394 Wettable Powder on Pepper
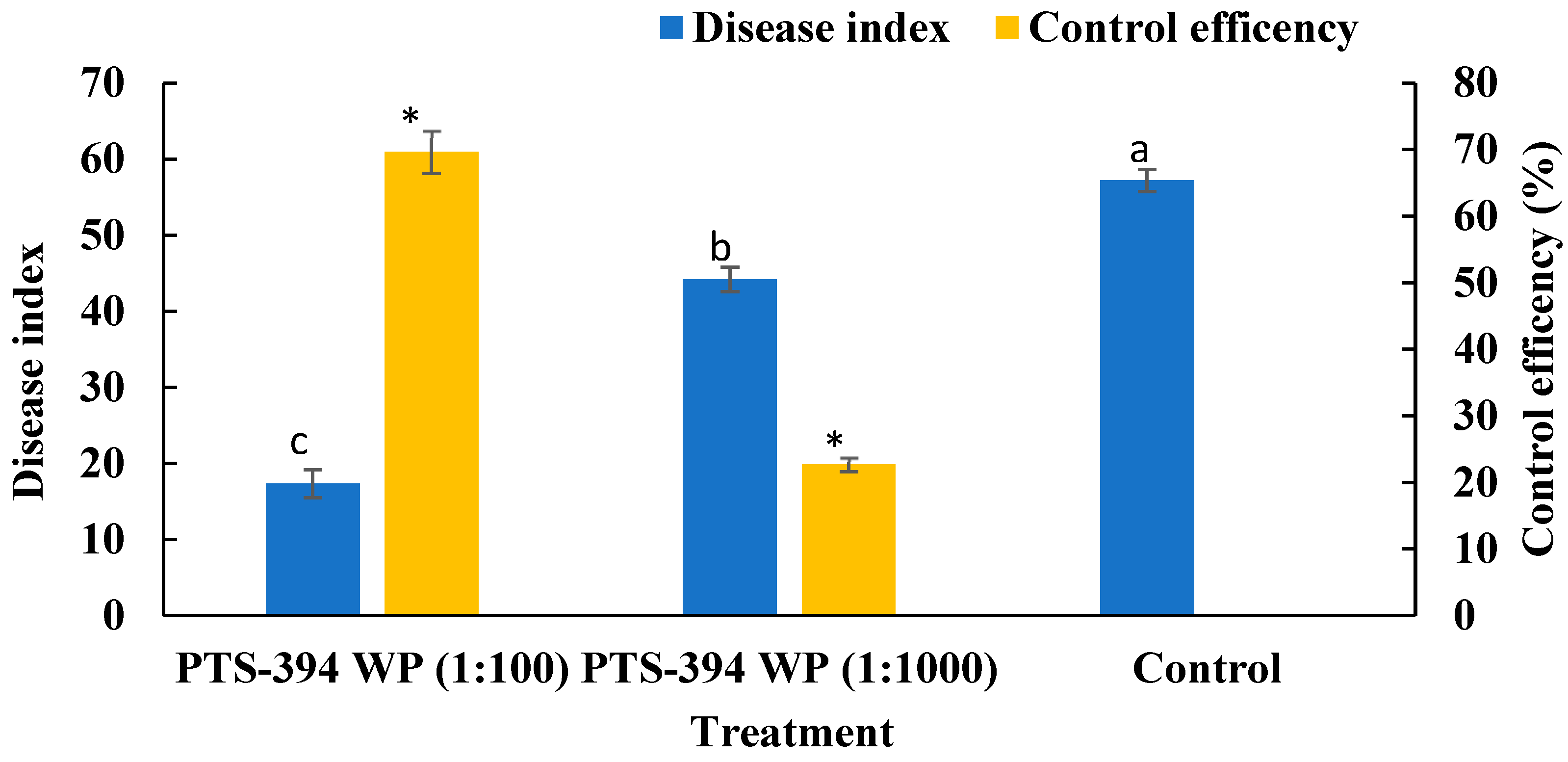
2.4. B. subtilis PTS-394 Wettable Powder Biocontrol Effect on Pepper Root Rot in Pot Experiments
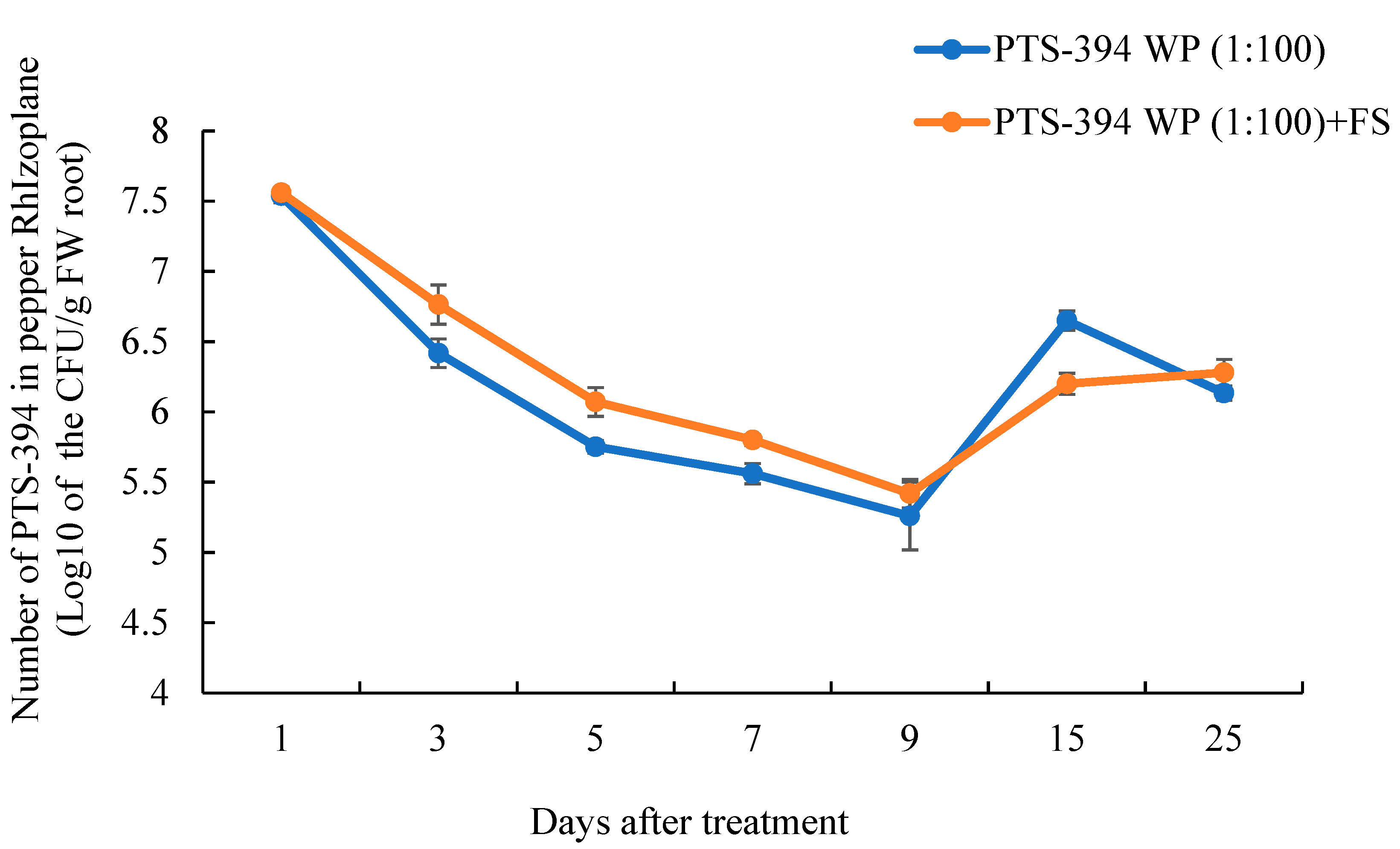
2.5. B. subtilis PTS-394 Dynamic Colonization on the Pepper Rhizoplane
2.6. B. subtilis PTS-394 Wettable Powder Bicontrol of Pepper Root Rot in Field
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bacterial Strains and Growth Conditions
5.2. Preparation of the Lipopeptides Crude Extract
5.3. Antifungal Activity Assay
5.4. Identification of Lipopeptides with MALDI-TOF-MS
5.5. Growth-Promoting Effects of B. subtilis PTS-394 Wettable Powder on Pepper
5.6. The Colonization of B. subtilis PTS-394 Wettable Powder on the Pepper Rhizoplane
5.7. The Biocontrol Effects of B. subtilis PTS-394 Wettable Powder in Pots
5.8. The Biocontrol Effects of B. subtilis PTS-394 Wettable Powder in Field
5.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Food and Agriculture Organization of the United Nations FAOSTAT Database. 2021. Available online: https://www.fao.org/faostat/zh/#data/QCL (accessed on 21 December 2022).
- Junqing, Q.; Dexu, L.; Yudong, S.; Zhichun, Z.; Yongfeng, L.; Youzhou, L. Investigation on the occurrence and control status of pests and diseases in pepper in Huai’an City, Jiangsu Province. Jiangsu Agric. Sci. 2019, 47, 127–132. [Google Scholar] [CrossRef]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borriss, R. Use of Plant-Associated Bacillus Strains as Biofertilizers and Biocontrol Agents in Agriculture. In Bacteria in Agrobiology: Plant Growth Responses; Maheshwari, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 41–76. [Google Scholar] [CrossRef]
- Weller, D.M. Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology 2007, 97, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Qiao, J.; Yu, X.; Liang, X.; Liu, Y.; Borriss, R.; Liu, Y. Addition of plant-growth-promoting Bacillus subtilis PTS-394 on tomato rhizosphere has no durable impact on composition of root microbiome. BMC Microbiol. 2017, 17, 131. [Google Scholar] [CrossRef]
- Bais, H.P.; Vivanco, F.J.M. Biocontrol of Bacillus subtilis against Infection of Arabidopsis Roots by Pseudomonas syringae Is Facilitated by Biofilm Formation and Surfactin Production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Gu, Q.; Qiao, J.; Wang, R.; Lu, J.; Wang, Z.; Li, P.; Zhang, L.; Ali, Q.; Khan, A.R.; Gao, X.; et al. The Role of Pyoluteorin from Pseudomonas protegens Pf-5 in Suppressing the Growth and Pathogenicity of Pantoea ananatis on Maize. Int. J. Mol. Sci. 2022, 23, 6431. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clement, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Syed-Ab-Rahman, S.F.; Arkhipov, A.; Wass, T.J.; Xiao, Y.; Carvalhais, L.C.; Schenk, P.M. Rhizosphere bacteria induce programmed cell death defence genes and signalling in chilli pepper. J. Appl. Microbiol. 2022, 132, 3111–3124. [Google Scholar] [CrossRef]
- Martinez-Raudales, I.; De La Cruz-Rodriguez, Y.; Alvarado-Gutierrez, A.; Vega-Arreguin, J.; Fraire-Mayorga, A.; Alvarado-Rodriguez, M.; Balderas-Hernandez, V.; Fraire-Velazquez, S. Draft genome sequence of Bacillus velezensis 2A-2B strain: A rhizospheric inhabitant of Sporobolus airoides (Torr.) Torr., with antifungal activity against root rot causing phytopathogens. Stand. Genom. Sci. 2017, 12, 73. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Huang, Y.; Li, Y.; Dong, J.; Liu, X.; Li, C. Biocontrol of Rhizoctonia solani via Induction of the Defense Mechanism and Antimicrobial Compounds Produced by Bacillus subtilis SL-44 on Pepper (Capsicum annuum L.). Front. Microbiol. 2019, 10, 2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Wu, Z.; He, Y.; Ye, B.C.; Li, C. Rhizospheric Bacillus subtilis Exhibits Biocontrol Effect against Rhizoctonia solani in Pepper (Capsicum annuum). Biomed. Res. Int. 2017, 2017, 9397619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cisternas-Jamet, J.; Salvatierra-Martinez, R.; Vega-Galvez, A.; Stoll, A.; Uribe, E.; Goni, M.G. Biochemical composition as a function of fruit maturity stage of bell pepper (Capsicum annum) inoculated with Bacillus amyloliquefaciens. Sci. Hortic-Amst. 2020, 263, 109107. [Google Scholar] [CrossRef]
- Liang, Y.R.; Liao, F.C.; Huang, T.P. Deciphering the influence of Bacillus subtilis strain Ydj3 colonization on the vitamin C contents and rhizosphere microbiomes of sweet peppers. PLoS ONE 2022, 17, e0264276. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 2022, 11, 386. [Google Scholar] [CrossRef]
- Cawoy, H.; Debois, D.; Franzil, L.; De Pauw, E.; Thonart, P.; Ongena, M. Lipopeptides as main ingredients for inhibition of fungal phytopathogens by Bacillus subtilis/amyloliquefaciens. Microb. Biotechnol. 2015, 8, 281–295. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microb. Biot. 2017, 33, 197. [Google Scholar] [CrossRef] [Green Version]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Qiao, J.; Liu, Y.; Liang, X.; Hu, Y.; Du, Y. Draft Genome Sequence of Root-Colonizing Bacterium Bacillus sp. Strain PTS-394. Genome. Announc. 2014, 2, e00038-14. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, Z.; Liang, X.; Zhu, J. Screening, Evaluation and Identification of Antagonistic Bacteria against Fusarium oxysporum f. sp. lycopersici and Ralstonia solanacearum. Chin. J. Biol. Control 2012, 28, 101–108. [Google Scholar]
- Junqing, Q.; Xinning, Z.; Xuejie, L.; Yongfeng, L.; Youzhou, L. Plant System Resistance Triggered by Root-colonizing Bacillus subtilis PTS−394 and Its Control Effect on Tomato Gray Mold. Chin. J. Biol. Control 2017, 33, 219–225. [Google Scholar] [CrossRef]
- Liang, X.; Liu, Y.; Zhang, R.; Qiao, J.; Chen, Z.; Lu, F. Optimization of fementation medium and conditions of Bacillus subtilis PTS-394. J. Yangzhou Univ. 2013, 34, 75–82. [Google Scholar]
- Adesemoye, A.O.; Torbert, H.A.; Kloepper, J.W. Plant growth-promoting rhizobacteria allow reduced application rates of chemical fertilizers. Microb. Ecol. 2009, 58, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Qiang, R.; Zhou, Z.J.; Pan, Y.; Yu, S.Q.; Yuan, W.; Cheng, J.N.; Wang, J.H.; Zhao, D.M.; Zhu, J.H.; et al. Biocontrol and Action Mechanism of Bacillus subtilis Lipopeptides’ Fengycins Against Alternaria solani in Potato as Assessed by a Transcriptome Analysis. Front. Microbiol. 2022, 13, 861113. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Selvaraj, J.N.; Xing, F.G.; Zhou, L.; Wang, Y.; Song, H.M.; Tan, X.X.; Sun, L.C.; Sangare, L.; Folly, Y.M.E.; et al. Antagonistic Action of Bacillus subtilis Strain SG6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar] [CrossRef]
- Ongena, M.; Jourdan, E.; Adam, A.; Paquot, M.; Brans, A.; Joris, B.; Arpigny, J.L.; Thonart, P. Surfactin and fengycin lipopeptides of Bacillus subtilis as elicitors of induced systemic resistance in plants. Env. Microbiol. 2007, 9, 1084–1090. [Google Scholar] [CrossRef]
- Li, B.; Li, Q.; Xu, Z.; Zhang, N.; Shen, Q.; Zhang, R. Responses of beneficial Bacillus amyloliquefaciens SQR9 to different soilborne fungal pathogens through the alteration of antifungal compounds production. Front. Microbiol. 2014, 5, 636. [Google Scholar] [CrossRef] [Green Version]
- Fan, B.; Chen, X.H.; Budiharjo, A.; Bleiss, W.; Vater, J.; Borriss, R. Efficient colonization of plant roots by the plant growth promoting bacterium Bacillus amyloliquefaciens FZB42, engineered to express green fluorescent protein. J. Biotechnol. 2011, 151, 303–311. [Google Scholar] [CrossRef]
- Junqing, Q.; Dexu, L.; Biao, P.; Yudong, S.; Yongfeng, L.; Youzhou, L. The Evaluation of Impact on Pepper Root and Rhizosphere Microbial Community Exerted by Bacillus subtilis PTS-394 Water Powder. Chin. J. Biol. Control 2018, 34, 287–293. [Google Scholar] [CrossRef]
- Ma, X.; Wang, X.B.; Cheng, J.; Nie, X.; Yu, X.X.; Zhao, Y.T.; Wang, W. Microencapsulation of Bacillus subtilis B99-2 and its biocontrol efficiency against Rhizoctonia solani in tomato. Biol. Control 2015, 90, 34–41. [Google Scholar] [CrossRef]
- Panpan, J. Study on the Control Efficiency and Induced Resistance of Bacillus Amyloliquefaciens B1619 against Root Knot Nematode Disease; Nanjing Agricultural University: Nanjing, China, 2016. [Google Scholar]
- Xiang, Y.P.; Zhou, H.F.; Liu, Y.F.; Chen, Z.Y. Isolation and Identification of Lipopeptide Antibiotics Produced by Bacillus amyloliquefaciens B1619 and the Inhibition of the Lipopeptide Antibiotics to Fusarium oxysporum f. sp. lycopersici. entia. Agric. Sin. 2016, 49, 2935–2944. [Google Scholar]
- Qiao, J.; Chen, Z.; Liang, X.; Liu, Y.; Liu, Y. Biocontrol Efficacy on Tomato Bacterial Wilt by Bacillus subtilis Bs916. Chin. J. Biol. Control 2016, 32, 229–234. [Google Scholar]
- Chen, Z.Y.; Liu, Y.F.; Lu, F. Study on key technology in the industrialized production of Bacillus subtilis Bs-916,the rice sheath blight control agent. J. Plant Prot. 2004, 31, 230–234. [Google Scholar]
- Zhang, L.N.; Wang, D.C.; Hu, Q.; Dai, X.Q.; Xie, Y.S.; Li, Q.; Liu, H.M.; Guo, J.H. Consortium of Plant Growth-Promoting Rhizobacteria Strains Suppresses Sweet Pepper Disease by Altering the Rhizosphere Microbiota. Front. Microbiol. 2019, 10, 1668. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.J.; Luan, P.Y.; Liu, S.F.; Shan, Y.T.; Hou, Z.P.; Zhao, S.Y.; Jia, S.; Li, R.F. Efficacy of Bacillus subtilis XZ18-3 as a Biocontrol Agent against Rhizoctonia cerealis on Wheat. Agriculture 2022, 12, 258. [Google Scholar] [CrossRef]
- Davis, W.W.; Stout, T.R. Disc plate method of microbiological antibiotic assay. I. Factors influencing variability and error. Appl. Microbiol. 1971, 22, 659–665. [Google Scholar] [CrossRef]
- Li, S.; Xu, J.; Fu, L.; Xu, G.; Lin, X.; Qiao, J.; Xia, Y. Biocontrol of Wheat Crown Rot Using Bacillus halotolerans QTH8. Pathogens 2022, 11, 595. [Google Scholar] [CrossRef]
- Liang, Z.; Qiao, J.Q.; Li, P.P.; Zhang, L.L.; Qiao, Z.X.; Lin, L.; Yu, C.J.; Yang, Y.; Zubair, M.; Gu, Q.; et al. A novel Rap-Phr system in Bacillus velezensis NAU-B3 regulates surfactin production and sporulation via interaction with ComA. Appl. Microbiol. Biotechnol. 2020, 104, 10059–10074. [Google Scholar] [CrossRef]




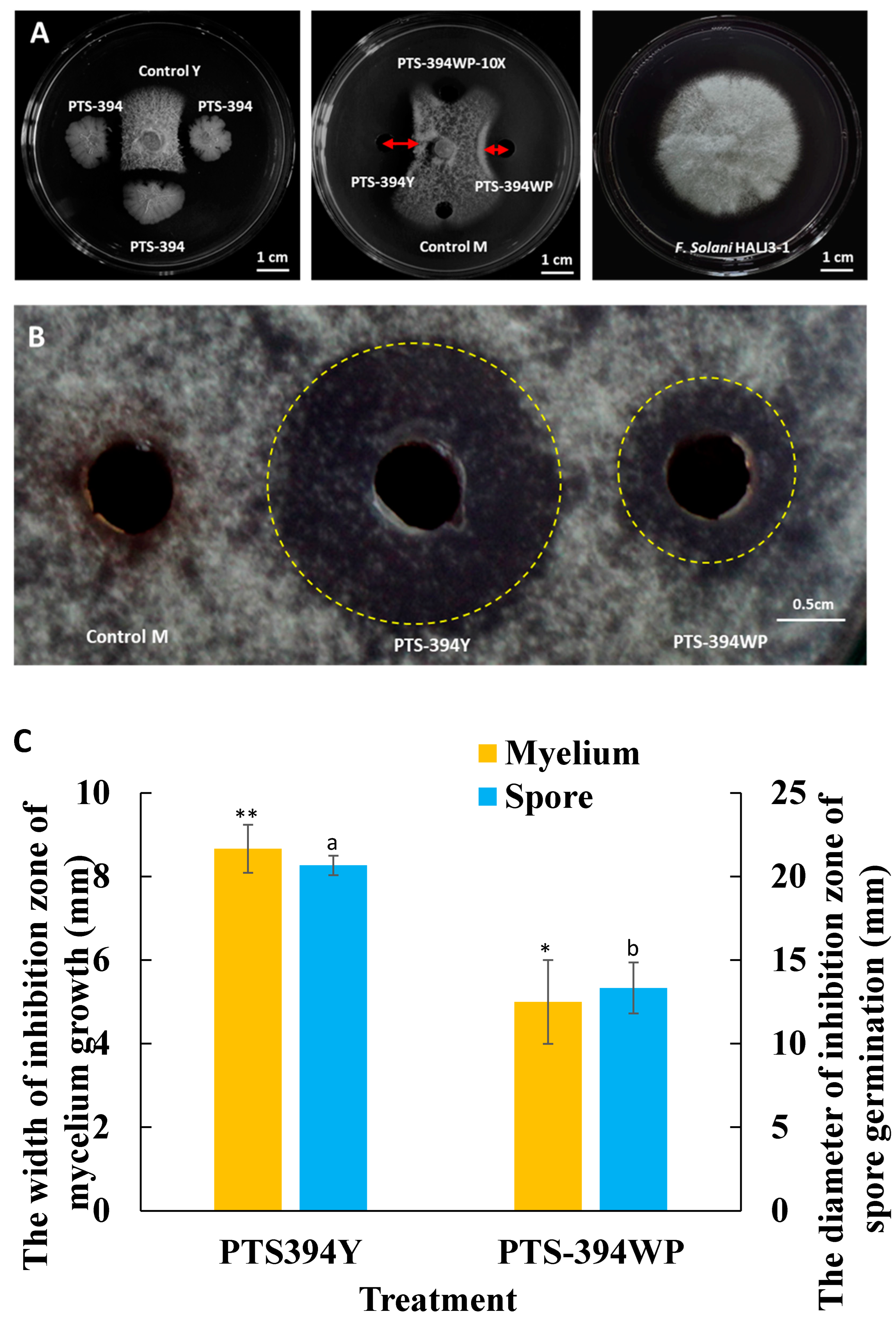{kind=link}
{kind=link}
{kind=link}
| Mass Peaks (M/Z) | Products | Mass Peaks in PTS-394 YPG M/Z (Intensity) [M + H, Na, K]+ | Mass Peaks in PTS-394WP M/Z (Intensity) [M + H, Na, K]+ |
|---|---|---|---|
| Surfactin | |||
| 993.6 | C13 Surfactin B | 1016.6 (15,521.25) [M + Na]+ | - |
| 1007.6 | C13 Surfactin A C14 Surfactin B C13 Surfactin C | 1030.6 (84,069.20)/1046.6 (27,524.07) [M + Na, K]+ | - |
| 1021.6 | C14 Surfactin A C15 Surfactin B C14 Surfactin C | 1044.6 (110,886.84)/1060.6 (44,451.90) [M + Na, K]+ | 1044.6 (356.55) [M + Na] + |
| 1035.7 | C15 Surfactin A C16 Surfactin B C15 Surfactin C | 1058.7 (121,828.87)/1074.6 (67,291.75) [M + Na, K]+ | 1058.7 (910.61)/1074.6 (408.72) [M + Na, K]+ |
| 1049.6 | C16 Surfactin A C16 Surfactin C | 1072.7 (25,248.00)/1088.7 (7889.02) [M + Na, K]+ | 1088.7 (382.82) [M + K]+ |
| Iturin | |||
| 1043.5 | C14 Iturin B | 1066.7 (6712.2) [M + Na]+ | - |
| 1057.5 | C15 Iturin B | 1080.7 (9859.62) [M + Na]+ | 1096.5 (339.58) [M + Na]+ |
| 1085.6 | C17 Iturin B | 1086.7 (6015.7) [M + H]+ | - |
| Fengycin | |||
| 1476.8 | C15 Fengycin B C17 Fengycin A | 1499.8(18,130.15) [M + Na ]+ | - |
| 1490.8 | C18 Fengycin A C16 Fengycin B | 1513.8 (21,506.17)/1529.8 (6258.52) [M + Na, K]+ | 1491.8 (221.47) [M + H]+ |
| 1504.8 | C17 Fengycin B | 1527.8 (18,134.60)/1543.8 (5152.01) [M + Na, K]+ | - |
| Treatment | Germination Percentage on Plate (%) | Germination Percentage in Pot (%) | Height of Plant (cm) |
|---|---|---|---|
| PTS-394 WP 1:10 | 46.67 ± 3.33 c | No | No |
| PTS-394 WP 1:100 | 80.00 ± 3.33 a | 72.22 ± 3.85 a | 21.21 ± 2.38 a |
| PTS-394 WP 1:1000 | 81.11 ± 5.09 a | 75.56 ± 1.92 a | 20.75 ± 2.59 a |
| Control | 71.11 ± 1.92 b | 63.33 ± 3.33 b | 17.54 ± 2.52 b |
| Treatment | Incidence of Disease (%) | Control Efficiency (%) |
|---|---|---|
| PTS-394 WP 1:100 | 2.06 ± 0.16 c | 74.44 ± 0.96 a |
| PTS-394 WP 1:200 | 2.50 ± 0.28 b | 68.57 ± 3.78 b |
| Control | 7.96 ±0.32 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, J.; Zhang, R.; Liu, Y.; Liu, Y. Evaluation of the Biocontrol Efficiency of Bacillus subtilis Wettable Powder on Pepper Root Rot Caused by Fusarium solani. Pathogens 2023, 12, 225. https://doi.org/10.3390/pathogens12020225
Qiao J, Zhang R, Liu Y, Liu Y. Evaluation of the Biocontrol Efficiency of Bacillus subtilis Wettable Powder on Pepper Root Rot Caused by Fusarium solani. Pathogens. 2023; 12(2):225. https://doi.org/10.3390/pathogens12020225
Chicago/Turabian StyleQiao, Junqing, Rongsheng Zhang, Yongfeng Liu, and Youzhou Liu. 2023. "Evaluation of the Biocontrol Efficiency of Bacillus subtilis Wettable Powder on Pepper Root Rot Caused by Fusarium solani" Pathogens 12, no. 2: 225. https://doi.org/10.3390/pathogens12020225