
Cytokine Networks as Targets for Preventing and Controlling Chagas Heart Disease
 ,
,  ,
,
Abstract
:1. Introduction
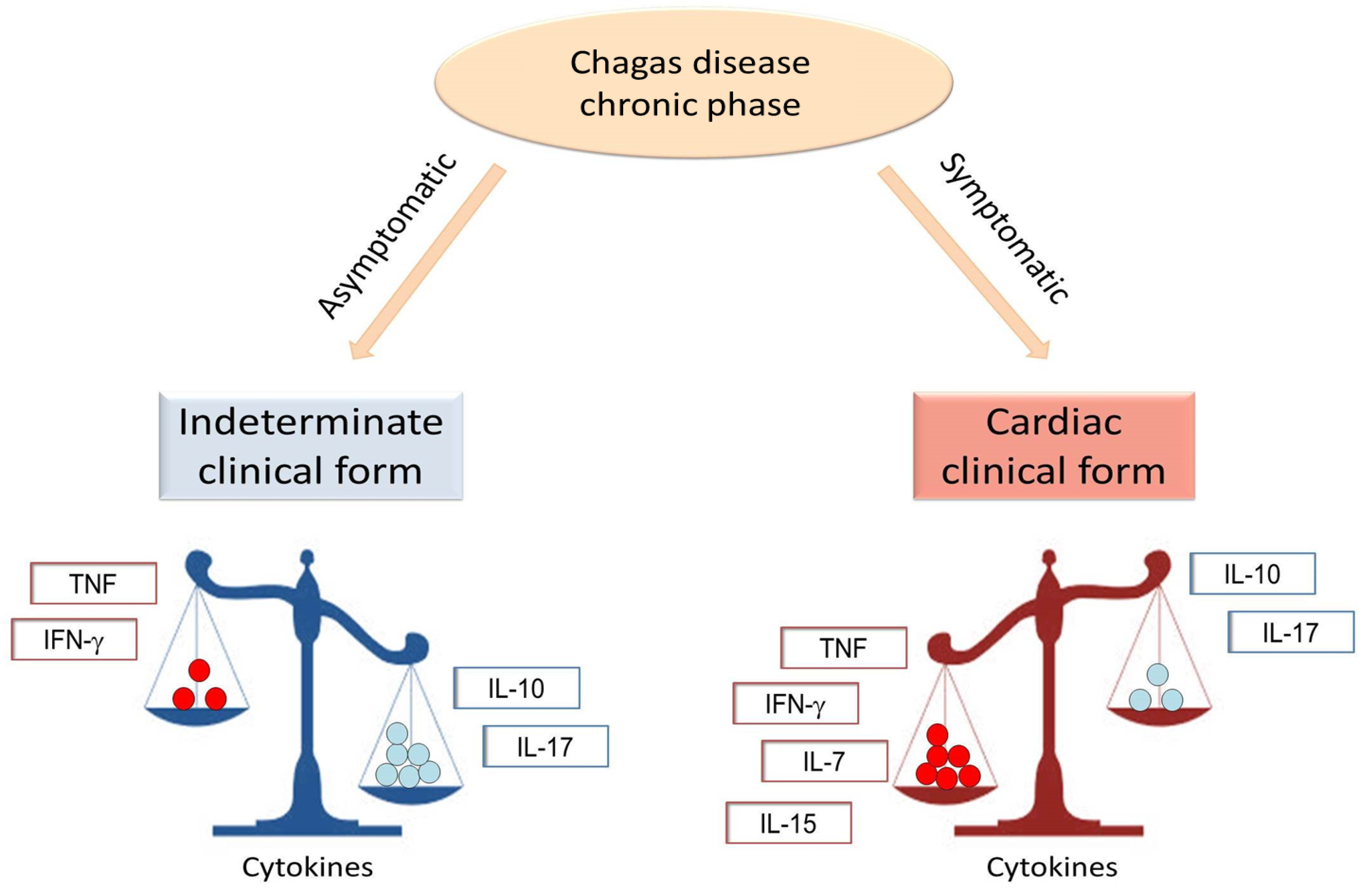
2. The Role of Cytokines in Immune Regulation during the Chronic Phase of Chagas Disease
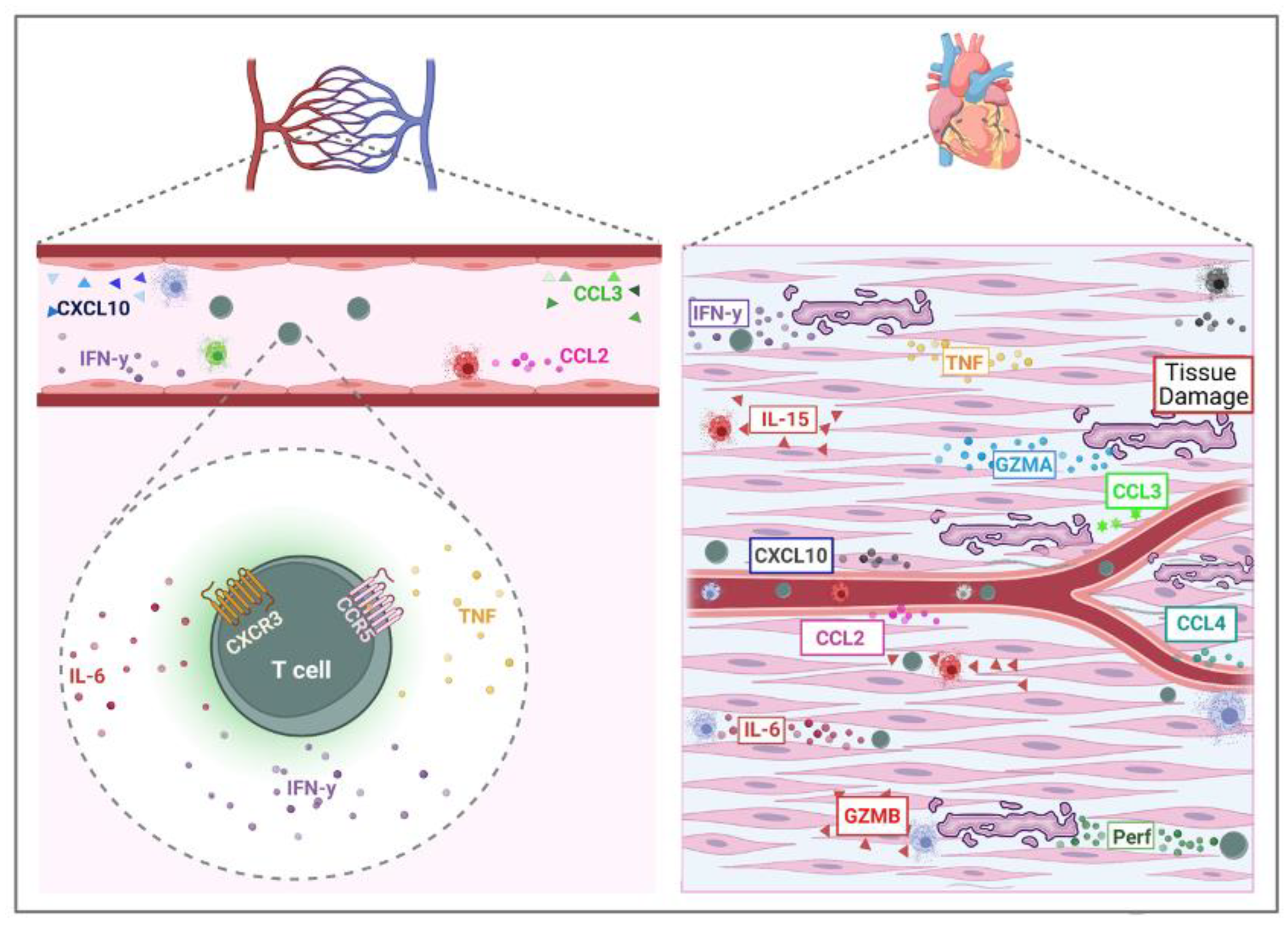
3. Cell Cytotoxicity in the Pathogenesis of Chronic Chagas Cardiomyopathy (CCC)
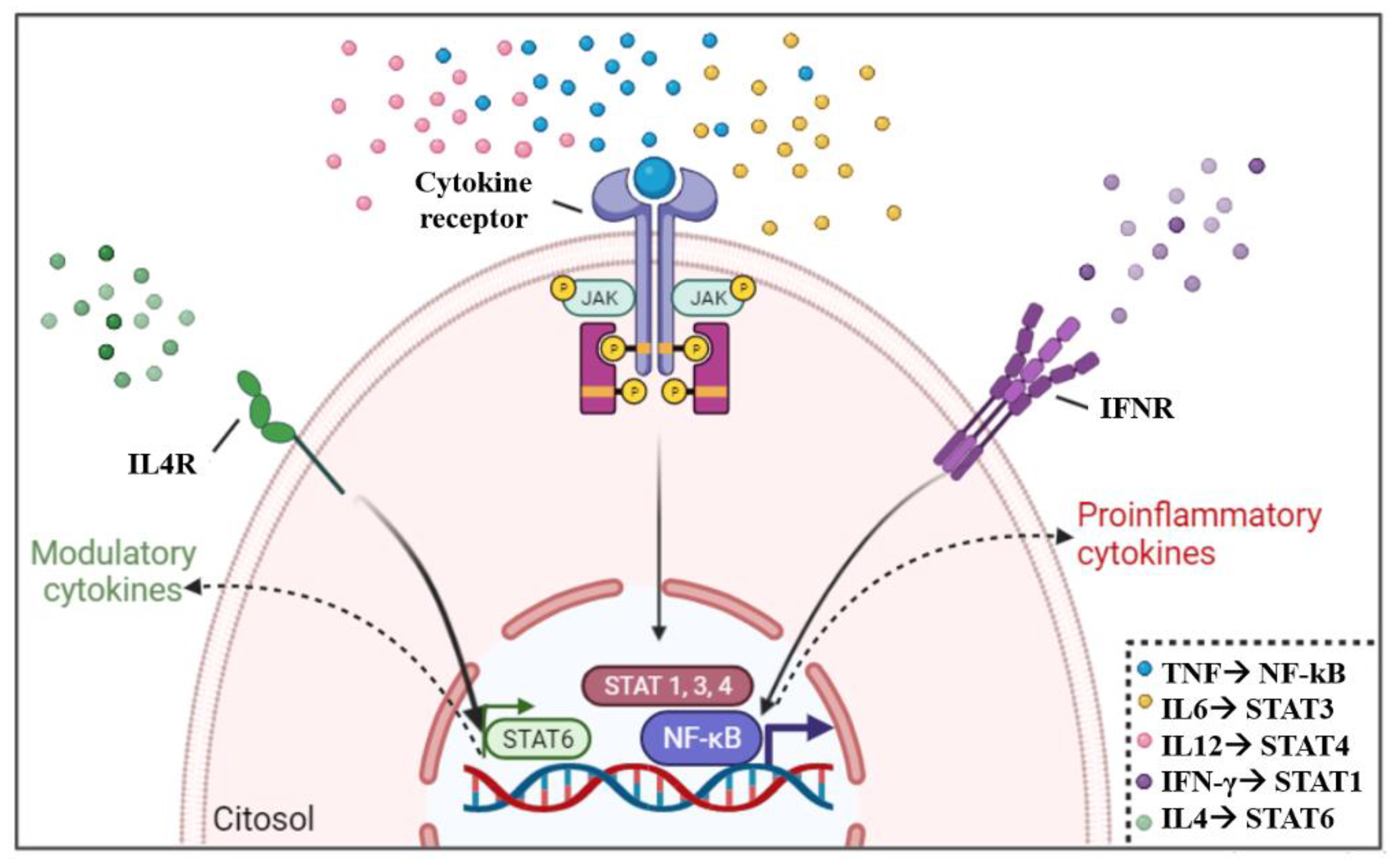
4. Control of Cytokine Expression
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olivera, M.J.; Porras-Villamil, J.F.; Villar, J.C.; Herrera, E.V.; Buitrago, G. Chagas Disease-Related Mortality in Colombia from 1979 to 2018: Temporal and Spatial Trends. Rev. Soc. Bras. Med. Trop. 2021, 54, e07682020. [Google Scholar] [CrossRef] [PubMed]
- Stanaway, J.D.; Roth, G. The Burden of Chagas Disease Estimates and Challenges. Glob. Heart. 2015, 10, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Valdez Tah, A.R. Making Sense of Chagas Disease among Mexican Immigrants in California. Med. Anthropol. 2021, 40, 511–524. [Google Scholar] [CrossRef] [PubMed]
- WHO. Chagas Disease in Latin America: An Epidemiological Update Based on 2010 Estimates. Wkly. Epidemiol. Rec. 2015, 90, 33–43. [Google Scholar]
- Schmunis, G.A.; Yadon, Z.E. Chagas Disease: A Latin American Health Problem Becoming a World Health Problem. Acta. Trop. 2010, 115, 14–21. [Google Scholar] [CrossRef]
- Jackson, Y.; Gétaz, L.; Wolff, H.; Holst, M.; Mauris, A.; Tardin, A.; Sztajzel, J.; Besse, V.; Loutan, L.; Gaspoz, J.M.; et al. Prevalence, Clinical Staging and Risk for Blood-Borne Transmission of Chagas Disease among Latin American Migrants in Geneva, Switzerland. PLoS Negl. Trop. Dis. 2010, 4, e592. [Google Scholar] [CrossRef] [Green Version]
- Shikanai-Yasuda, M.A.; Carvalho, N.B. Oral Transmission of Chagas Disease. Clin. Infect. Dis. 2012, 54, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Bern, C.; Messenger, L.A.; Whitman, J.D.; Maguire, J.H. Chagas Disease in the United States: A Public Health Approach. Clin. Microbiol. Rev. 2019, 33, e00023–e00119. [Google Scholar] [CrossRef]
- Dias, E.; Laranja, F.S.; Miranda, A.; Nobrega, G. Chagas’ Disease: A Clinical, Epidemiologic, and Pathologic Study. Circulation 1956, 14, 1035–1060. [Google Scholar] [CrossRef] [Green Version]
- Echavarría, N.G.; Echeverría, L.E.; Stewart, M.; Gallego, C.; Saldarriaga, C. Chagas Disease: Chronic Chagas Cardiomyopathy. Curr. Probl. Cardiol. 2021, 46, 100507. [Google Scholar] [CrossRef]
- Dutra, W.O.; Rocha, M.O.C.; Teixeira, M.M. The Clinical Immunology of Human Chagas Disease. Trends Parasitol. 2005, 21, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Candia-Puma, M.A.; Machaca-Luque, L.Y.; Roque-Pumahuanca, B.M.; Galdino, A.S.; Giunchetti, R.C.; Coelho, E.A.F.; Chávez-Fumagalli, M.A. Accuracy of Diagnostic Tests for the Detection of Chagas Disease: A Systematic Review and Meta-Analysis. Diagnostics 2022, 12, 2752. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.C.P.; Beaton, A.; Acquatella, H.; Bern, C.; Bolger, A.F.; Echeverría, L.E.; Dutra, W.O.; Gascon, J.; Morillo, C.A.; Oliveira-Filho, J.; et al. Chagas Cardiomyopathy: An Update of Current Clinical Knowledge and Management: A Scientific Statement From the American Heart Association. Circulation 2018, 138, e169–e209. [Google Scholar] [CrossRef] [PubMed]
- Benziger, C.P.; do Carmo, G.A.L.; Ribeiro, A.L.P. Chagas Cardiomyopathy: Clinical Presentation and Management in the Americas. Cardiol. Clin. 2017, 35, 31–47. [Google Scholar] [CrossRef]
- Franco, D.J.; Vago, A.R.; Chiari, E.; Meira, F.C.A.; Galvão, L.M.C.; Machado, C.R.S. Trypanosoma cruzi: Mixture of two populations can modify virulence and tissue tropism in tat. Exp. Parasitol. 2003, 104, 54–61. [Google Scholar] [CrossRef]
- Vago, A.R.; Andrade, L.O.; Leite, A.A.; d’Ávila Reis, D.; Macedo, A.M.; Adad, S.J.; Tostes, S.; da Consolação, M.V.M.; Filho, G.B.; Pena, S.D.J. Genetic Characterization of Trypanosoma cruzi Directly from Tissues of Patients with Chronic Chagas Disease: Differential Distribution of Genetic Types into Diverse Organs. Am. J. Pathol. 2000, 156, 1805–1809. [Google Scholar] [CrossRef]
- Rodrigues, C.M.; Valadares, H.M.S.; Francisco, A.F.; Arantes, J.M.; Campos, C.F.; Teixeira-Carvalho, A.; Martins-Filho, O.A.; Araujo, M.S.S.; Arantes, R.M.E.; Chiari, E.; et al. Coinfection with Different Trypanosoma cruzi Strains Interferes with the Host Immune Response to Infection. PLoS Negl. Trop. Dis. 2010, 4, e846. [Google Scholar] [CrossRef]
- Souza, P.E.A.; Rocha, M.O.C.; Rocha-Vieira, E.; Menezes, C.A.S.; Chaves, A.C.L.; Gollob, K.J.; Dutra, W.O. Monocytes from Patients with Indeterminate and Cardiac Forms of Chagas’ Disease Display Distinct Phenotypic and Functional Characteristics Associated with Morbidity. Infect. Immun. 2004, 72, 5283–5291. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, L.M.D.; Gollob, K.J.; Zingales, B.; Dutra, W.O. Pathogen Diversity, Immunity, and the Fate of Infections: Lessons Learned from Trypanosoma cruzi Human-Host Interactions. Lancet Microbe 2022, 3, e711–e722. [Google Scholar] [CrossRef]
- Casares-Marfil, D.; Strauss, M.; Bosch-Nicolau, P.; lo Presti, M.S.; Molina, I.; Chevillard, C.; Cunha-Neto, E.; Sabino, E.; Ribeiro, A.L.P.; González, C.I.; et al. A Genome-Wide Association Study Identifies Novel Susceptibility Loci in Chronic Chagas Cardiomyopathy. Clin. Infect. Dis. 2021, 73, 672–679. [Google Scholar] [CrossRef]
- Bartekova, M.; Radosinska, J.; Jelemensky, M.; Dhalla, N.S. Role of Cytokines and Inflammation in Heart Function during Health and Disease. Heart Fail Rev. 2018, 23, 733–758. [Google Scholar] [CrossRef] [PubMed]
- Llaguno, M.; da Silva, M.V.; Batista, L.R.; da Silva, D.A.A.; de Sousa, R.C.; de Resende, L.A.P.R.; da Silva, V.J.D.; Lages-Silva, E.; Oliveira, C.J.F.; Machado, J.R.; et al. T-Cell Immunophenotyping and Cytokine Production Analysis in Patients with Chagas Disease 4 Years after Benznidazole Treatment. Infect. Immun. 2019, 87, e00103–e00119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutra, W.O.; Gollob, K.J.; Pinto-Dias, J.C.; Gazzinelli, G.; Correa-Oliveira, R.; Coffman, R.L.; Carvalho-Parra, J.F. Cytokine MRNA Profile of Peripheral Blood Mononuclear Cells Isolated from Individuals with Trypanosoma cruzi Chronic Infection. Scand. J. Immunol. 1997, 45, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Abel, L.C.J.; Rizzo, L.v.; Ianni, B.; Albuquerque, F.; Bacal, F.; Carrara, D.; Bocchi, E.A.; Teixeira, H.C.; Mady, C.; Kalil, J.; et al. Chronic Chagas’ Disease Cardiomyopathy Patients Display an Increased IFN-gammaResponse to Trypanosoma cruzi Infection. J. Autoimmun. 2001, 17, 99–107. [Google Scholar] [CrossRef]
- Benvenuti, L.A.; Roggério, A.; Nishiya, A.S.; Campos, S.V.; Fiorelli, A.I.; Levi, J.E. Trypanosoma Cruzi Persistence in the Native Heart Is Associated with High-Grade Myocarditis, but Not with Chagas’ Disease Reactivation after Heart Transplantation. J. Heart Lung Transplant 2014, 33, 698–703. [Google Scholar] [CrossRef]
- Bestetti, R.B.; Dellalibera-Joviliano, R.; Lopes, G.S.; Faria-Jr, M.; Furlan-Daniel, R.; Lopes, K.C.; Batista, D.R. Determination of the Th1, Th2, Th17, and Treg Cytokine Profile in Patients with Chronic Chagas Heart Disease and Systemic Arterial Hypertension. Heart Vessels 2019, 34, 123–133. [Google Scholar] [CrossRef]
- Ferreira, R.C.; Ianni, B.M.; Abel, L.C.J.; Buck, P.; Mady, C.; Kalil, J.; Cunha-Neto, E. Increased Plasma Levels of Tumor Necrosis Factor-Alpha in Asymptomatic/“indeterminate” and Chagas Disease Cardiomyopathy Patients. Mem. Inst. Oswaldo. Cruz. 2003, 98, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Reis, D.D.; Jones, E.M.; Tostes, S.; Lopes, E.R.; Gazzinelli, G.; Colley, D.G.; McCurley, T.L. Characterization of Inflammatory Infiltrates in Chronic Chagasic Myocardial Lesions: Presence of Tumor Necrosis Factor-Alpha+ Cells and Dominance of Granzyme A+, CD8+ Lymphocytes. Am. J. Trop. Med. Hyg. 1993, 48, 637–644. [Google Scholar] [CrossRef]
- Reis, M.M.; Higuchi, M.D.L.; Benvenuti, L.A.; Aiello, V.D.; Gutierrez, P.S.; Bellotti, G.; Pileggi, F. An in Situ Quantitative Immunohistochemical Study of Cytokines and IL-2R+ in Chronic Human Chagasic Myocarditis: Correlation with the Presence of Myocardial Trypanosoma Cruzi Antigens. Clin. Immunol. Immunopathol. 1997, 83, 165–172. [Google Scholar] [CrossRef]
- Sousa, G.R.; Gomes, J.A.S.; Fares, R.C.G.; Damásio, M.P.D.S.; Chaves, A.T.; Ferreira, K.S.; Nunes, M.C.P.; Medeiros, N.I.; Valente, V.A.A.; Corrêa-Oliveira, R.; et al. Plasma Cytokine Expression Is Associated with Cardiac Morbidity in Chagas Disease. PLoS ONE 2014, 9, e87082. [Google Scholar] [CrossRef]
- Rodrigues, D.B.R.; dos Reis, M.A.; Romano, A.; Pereira, S.A.D.L.; Teixeira, V.D.P.A.; Tostes Junior, S.; Rodrigues, V. In Situ Expression of Regulatory Cytokines by Heart Inflammatory Cells in Chagas’ Disease Patients with Heart Failure. Clin. Dev. Immunol. 2012, 2012, 361730. [Google Scholar] [CrossRef] [PubMed]
- Talvani, A.; Rocha, M.O.C.; Barcelos, L.S.; Gomes, Y.M.; Ribeiro, A.L.; Teixeira, M.M. Elevated Concentrations of CCL2 and Tumor Necrosis Factor-α in Chagasic Cardiomyopathy. Clin. Infect. Dis. 2004, 38, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Neto, E.; Coelho, V.; Guilherme, L.; Fiorelli, A.; Stolf, N.; Kalil, J. Autoimmunity in Chagas’ Disease. Identification of Cardiac Myosin-B13 Trypanosoma cruzi Protein Crossreactive T Cell Clones in Heart Lesions of a Chronic Chagas’ Cardiomyopathy Patient. J. Clin. Investig. 1996, 98, 1709–1712. [Google Scholar] [CrossRef]
- Neves, E.G.A.; Koh, C.C.; Padilha da Silva, J.L.; Passos, L.S.A.; Villani, F.N.A.; dos Santos, J.S.C.; Menezes, C.A.S.; Silva, V.R.; Tormin, J.P.A.S.; Evangelista, G.F.B.; et al. Systemic Cytokines, Chemokines and Growth Factors Reveal Specific and Shared Immunological Characteristics in Infectious Cardiomyopathies. Cytokine 2021, 148, 155711. [Google Scholar] [CrossRef] [PubMed]
- Neves, E.G.A.; Koh, C.C.; Souza-Silva, T.G.; Passos, L.S.A.; Silva, A.C.C.; Velikkakam, T.; Villani, F.; Coelho, J.S.; Brodskyn, C.I.; Teixeira, A.; et al. T-Cell Subpopulations Exhibit Distinct Recruitment Potential, Immunoregulatory Profile and Functional Characteristics in Chagas versus Idiopathic Dilated Cardiomyopathies. Front. Cardiovasc. Med. 2022, 9, 787423. [Google Scholar] [CrossRef]
- Dutra, W.O.; Menezes, C.A.S.; Magalhães, L.M.D.; Gollob, K.J. Immunoregulatory Networks in Human Chagas Disease. Parasite Immunol. 2014, 36, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, L.G.; Santos, R.H.B.; Fiorelli, A.I.; Mairena, E.C.; Benvenuti, L.A.; Bocchi, E.A.; Stolf, N.A.; Kalil, J.; Cunha-Neto, E. Myocardial Gene Expression of T-Bet, GATA-3, Ror-Γt, FoxP3, and Hallmark Cytokines in Chronic Chagas Disease Cardiomyopathy: An Essentially Unopposed TH1-Type Response. Mediat. Inflamm. 2014, 2014, 914326. [Google Scholar] [CrossRef] [Green Version]
- Guedes, P.M.M.; Gutierrez, F.R.S.; Silva, G.K.; Dellalibera-Joviliano, R.; Rodrigues, G.J.; Bendhack, L.M.; Rassi Jr, A.; Rassi, A.; Schmidt, A.; Maciel, B.C.; et al. Deficient Regulatory T Cell Activity and Low Frequency of IL-17-Producing T Cells Correlate with the Extent of Cardiomyopathy in Human Chagas’ Disease. PLoS Negl. Trop. Dis. 2012, 6, e1630. [Google Scholar] [CrossRef]
- Gomes, J.A.S.; Rocha, M.O.C.; Gazzinelli, G.; Gomes, J.A.S.; Rocha, M.O.C. Evidence That Development of Severe Cardiomyopathy in Human Chagas’ Disease Is Due to a Th1-Specific Immune Response Evidence That Development of Severe Cardiomyopathy in Human Chagas’ Disease Is Due to a Th1-Specific Immune Response. Infect. Immun. 2003, 71, 1185–1193. [Google Scholar] [CrossRef] [Green Version]
- Vitelli-Avelar, D.M.; Sathler-Avelar, R.; Teixeira-Carvalho, A.; Pinto Dias, J.C.; Gontijo, E.D.; Faria, A.M.; Elói-Santos, S.M.; Martins-Filho, O.A. Strategy to Assess the Overall Cytokine Profile of Circulating Leukocytes and Its Association with Distinct Clinical Forms of Human Chagas Disease. Scand. J. Immunol. 2008, 68, 516–525. [Google Scholar] [CrossRef]
- Passos, L.S.A.; Magalhães, L.M.D.; Soares, R.P.; Marques, A.F.; Alves, M.L.R.; Giunchetti, R.C.; Nunes, M.d.C.P.; Gollob, K.J.; Dutra, W.O. Activation of Human CD11b+ B1 B-Cells by Trypanosoma cruzi-Derived Proteins Is Associated with Protective Immune Response in Human Chagas Disease. Front. Immunol. 2019, 9, 3015. [Google Scholar] [CrossRef] [PubMed]
- Berbert, L.R.; González, F.B.; Villar, S.R.; Vigliano, C.; Lioi, S.; Beloscar, J.; Bottasso, O.A.; Silva-Barbosa, S.D.; Savino, W.; Pérez, A.R. Enhanced Migratory Capacity of T Lymphocytes in Severe Chagasic Patients Is Correlated With VLA-4 and TNF-α Expression. Front. Cell. Infect. Microbiol. 2021, 11, 782. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.T.; Costa, H.S.; de Lima, V.P.; Xavier, D.M.; Mendonça, V.A.; Lacerda, A.C.R.; Lage, V.K.S.; Lima, M.M.O.; Rocha, M.O.C.; Figueiredo, P.H.S. Plasma Levels of Soluble TNF Receptors Are Associated with Cardiac Function in Patients with Chagas Heart Disease. Int. J. Cardiol. 2020, 316, 101–103. [Google Scholar] [CrossRef] [PubMed]
- de Alba-Alvarado, M.; Salazar-Schettino, P.M.; Jiménez-Álvarez, L.; Cabrera-Bravo, M.; García-Sancho, C.; Zenteno, E.; Vazquez-Antona, C.; Cruz-Lagunas, A.; Zúñiga, J.; Bucio-Torres, M.I. Th-17 Cytokines Are Associated with Severity of Trypanosoma cruzi Chronic Infection in Pediatric Patients from Endemic Areas of Mexico. Acta Trop. 2018, 178, 134–141. [Google Scholar] [CrossRef]
- López, L.; Arai, K.; Giménez, E.; Jiménez, M.; Pascuzo, C.; Rodríguez-Bonfante, C.; Bonfante-Cabarcas, R. Las Concentraciones Séricas de Interleucina-6 y Proteína C Reactiva Se Incrementan a Medida Que La Enfermedad de Chagas Evoluciona Hacia El Deterioro de La Función Cardíaca. Rev. Esp. Cardiol. 2006, 59, 50–56. [Google Scholar] [CrossRef]
- de Araújo, F.F.; Lima Torres, K.C.; Viana Peixoto, S.; Pinho Ribeiro, A.L.; Vaz Melo Mambrini, J.; Bortolo Rezende, V.; Lima Silva, M.L.; Loyola Filho, A.I.; Teixeira-Carvalho, A.; Lima-Costa, M.F.; et al. CXCL9 and CXCL10 Display an Age-Dependent Profile in Chagas Patients: A Cohort Study of Aging in Bambui, Brazil. Infect. Dis. Poverty 2020, 9, 51. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.R.; Waghabi, M.C.; Bailly, S.; Feige, J.J.; Hasslocher-Moreno, A.M.; Saraiva, R.M.; Araujo-Jorge, T.C. The Search for Biomarkers and Treatments in Chagas Disease: Insights From TGF-Beta Studies and Immunogenetics. Front. Cell. Infect. Microbiol. 2022, 11, 1310. [Google Scholar] [CrossRef]
- Cutrullis, R.A.; Petray, P.B.; Schapachnik, E.; Sánchez, R.; Postan, M.; González, M.N.; Martín, V.; Corral, R.S. Elevated Serum Levels of Macrophage Migration Inhibitory Factor Are Associated with Progressive Chronic Cardiomyopathy in Patients with Chagas Disease. PLoS ONE 2013, 8, e57181. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Robles, D.; Vargas-Alarcón, G.; Ortíz-Muñiz, R.; Reyes, P.A.; Monteon, V.M. Serum Cytokines and Activation Ex Vivo of CD4+ and CD8+ T Cells in Chagasic Chronic Mexican Patients. Ann. Parasitol. 2017, 63, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Villani, F.N.A.; Rocha, M.O.D.C.; Nunes, M.D.C.P.; Antonelli, L.R.D.V.; Magalhães, L.M.D.; dos Santos, J.S.C.; Gollob, K.J.; Dutra, W.O. Trypanosoma cruzi-Induced Activation of Functionally Distinct ab and gd CD4- CD8- T Cells in Individuals with Polar Forms of Chagas’ Disease. Infect. Immun. 2010, 78, 4421–4430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, P.E.A.; Rocha, M.O.C.; Menezes, C.A.S.; Coelho, J.S.; Chaves, A.C.L.; Gollob, K.J.; Dutra, W.O. Trypanosoma cruzi Infection Induces Differential Modulation of Costimulatory Molecules and Cytokines by Monocytes and T Cells from Patients with Indeterminate and Cardiac Chagas’ Disease. Infect. Immun. 2007, 75, 1886–1894. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, L.M.D.; Villani, F.N.A.; Nunes, M.D.C.P.; Gollob, K.J.; Rocha, M.O.C.; Dutra, W.O. High Interleukin 17 Expression Is Correlated with Better Cardiac Function in Human Chagas Disease. J. Infect. Dis. 2013, 207, 661–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campi-Azevedo, A.C.; Gomes, J.A.S.; Teixeira-Carvalho, A.; Silveira-Lemos, D.; Vitelli-Avelar, D.M.; Sathler-Avelar, R.; Peruhype-Magalhães, V.; Béla, S.R.; Silvestre, K.F.; Batista, M.A.; et al. Etiological Treatment of Chagas Disease Patients with Benznidazole Lead to a Sustained Pro-Inflammatory Profile Counterbalanced by Modulatory Events. Immunobiology 2015, 220, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, N.I.; Gomes, J.A.S.; Fiuza, J.A.; Sousa, G.R.; Almeida, E.F.; Novaes, R.O.; Rocha, V.L.S.; Chaves, A.T.; Dutra, W.O.; Rocha, M.O.C.; et al. MMP-2 and MMP-9 Plasma Levels Are Potential Biomarkers for Indeterminate and Cardiac Clinical Forms Progression in Chronic Chagas Disease. Sci. Rep. 2019, 9, 14170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista-López, N.L.; Morillo, C.A.; López-Jaramillo, P.; Quiroz, R.; Luengas, C.; Silva, S.Y.; Galipeau, J.; Lalu, M.M.; Schulz, R. Matrix Metalloproteinases 2 and 9 as Diagnostic Markers in the Progression to Chagas Cardiomyopathy. Am. Heart J. 2013, 165, 558–566. [Google Scholar] [CrossRef]
- Fares, R.C.G.; Correa-Oliveira, R.; de Araújo, F.F.; Keesen, T.S.L.; Chaves, A.T.; Fiuza, J.A.; Ferreira, K.S.; Rocha, M.O.C.; Gomes, J.A.S. Identification of Phenotypic Markers of B Cells from Patients with Chagas Disease. Parasite Immunol. 2013, 35, 214–223. [Google Scholar] [CrossRef]
- Okamoto, E.E.; Sherbuk, J.E.; Clark, E.H.; Marks, M.A.; Gandarilla, O.; Galdos-Cardenas, G.; Vasquez-Villar, A.; Choi, J.; Crawford, T.C.; Q, R.; et al. Biomarkers in Trypanosoma cruzi-Infected and Uninfected Individuals with Varying Severity of Cardiomyopathy in Santa Cruz, Bolivia. PLoS Negl. Trop. Dis. 2014, 8, e3227. [Google Scholar] [CrossRef]
- Baron, M.A.; Ferreira, L.R.P.; Teixeira, P.C.; Moretti, A.I.S.; Santos, R.H.B.; Frade, A.F.; Kuramoto, A.; Debbas, V.; Benvenuti, L.A.; Gaiotto, F.A.; et al. Matrix Metalloproteinase 2 and 9 Enzymatic Activities Are Selectively Increased in the Myocardium of Chronic Chagas Disease Cardiomyopathy Patients: Role of TIMPs. Front. Cell. Infect. Microbiol. 2022, 12, 249. [Google Scholar] [CrossRef]
- De Oliveira, A.P.; Ayo, C.M.; Bestetti, R.B.; de Mattos, C.C.B.; Cavasini, C.E.; de Mattos, L.C. The Role of CCR5 in Chagas Disease—A Systematic Review. Infect. Genet. Evol. 2016, 45, 132–137. [Google Scholar] [CrossRef]
- Nogueira, L.G.; Santos, R.H.B.; Ianni, B.M.; Fiorelli, A.I.; Mairena, E.C.; Benvenuti, L.A.; Frade, A.; Donadi, E.; Dias, F.; Saba, B.; et al. Myocardial Chemokine Expression and Intensity of Myocarditis in Chagas Cardiomyopathy Are Controlled by Polymorphisms in CXCL9 and CXCL10. PLoS Negl. Trop. Dis. 2012, 6, e1867. [Google Scholar] [CrossRef]
- Luz, P.R.; Velavan, T.P.; Kremsner, P.G.; Messias-Reason, I.J.T. Association of IP-10 and PDGF-BB Levels with Clinical Forms of Chronic Chagas Disease. Int. J. Cardiol. 2013, 169, e53–e55. [Google Scholar] [CrossRef] [PubMed]
- Roffe, E.; dos Santos, L.I.; Santos, M.O.; Henriques, P.M.; Teixeira-Carvalho, A.; Martins-Filho, O.A.; Rocha, M.O.C.; Eloi-Santos, S.M.; Correa-Oliveira, R.; Antonelli, L.R.V. Increased Frequencies of Circulating CCR5+ Memory T Cells Are Correlated to Chronic Chagasic Cardiomyopathy Progression. J. Leukoc. Biol. 2019, 106, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Batista, A.M.; Alvarado-Arnez, L.E.; Alves, S.M.; Melo, G.; Pereira, I.R.; de Souza Ruivo, L.A.; da Silva, A.A.; Gibaldi, D.; da Silva, T.d.E.S.; de Lorena, V.M.B.; et al. Genetic Polymorphism at CCL5 Is Associated with Protection in Chagas’ Heart Disease: Antagonistic Participation of CCR1+ and CCR5+ Cells in Chronic Chagasic Cardiomyopathy. Front. Immunol. 2018, 9, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, J.A.S.; Bahia-Oliveira, L.M.G.; Rocha, M.O.C.; Busek, S.C.U.; Tekeira, M.M.; Silva, J.S.; Correa-Oliveira, R. Type 1 Chemokine Receptor Expression in Chagas’ Disease Correlates with Morbidity in Cardiac Patients. Infect. Immun. 2005, 73, 7960–7966. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Olarte, S.; Bolaños, N.I.; Echeverry, M.; Rodríguez, A.N.; Cuéllar, A.; Puerta, C.J.; Mariño, A.; González, J.M. Intermediate Monocytes and Cytokine Production Associated with Severe Forms of Chagas Disease. Front. Immunol. 2019, 10, 1671. [Google Scholar] [CrossRef] [Green Version]
- Dutra, W.O.; Martins-Filho, O.A.; Caņado, J.R.; Pinto-dias, J.C.; Brener, Z.; Freeman, G.L.; Colley, D.G.; Gazzinelll, G.; Parra, J.C. Activated T and B Lymphocytes in Peripheral Blood of Patients with Chagas’ Disease. Int. Immunol. 1994, 6, 499–506. [Google Scholar] [CrossRef]
- Menezes, C.A.S.; Rocha, M.O.C.; Souza, P.E.A.; Chaves, A.C.L.; Gollob, K.J.; Dutra, W.O. Phenotypic and Functional Characteristics of CD28+ and CD28- Cells from Chagasic Patients: Distinct Repertoire and Cytokine Expression. Clin. Exp. Immunol. 2004, 137, 129–138. [Google Scholar] [CrossRef]
- Velikkakam, T.; Gollob, K.J.; Dutra, W.O. Double-Negative T Cells: Setting the Stage for Disease Control or Progression. Immunology 2022, 165, 371–385. [Google Scholar] [CrossRef]
- Passos, L.S.A.; Magalhães, L.M.D.; Soares, R.P.; Marques, A.F.; Nunes, M.C.P.; Gollob, K.J.; Dutra, W.O. Specific Activation of CD4- CD8- Double-Negative T Cells by Trypanosoma cruzi-Derived Glycolipids Induces a Proinflammatory Profile Associated with Cardiomyopathy in Chagas Patients. Clin. Exp. Immunol. 2017, 190, 122–132. [Google Scholar] [CrossRef] [Green Version]
- Passos, L.S.A.; Koh, C.C.; Magalhães, L.M.D.; Nunes, M.C.P.; Gollob, K.J.; Dutra, W.O. Distinct CD4-CD8- (Double-Negative) Memory T-Cell Subpopulations Are Associated With Indeterminate and Cardiac Clinical Forms of Chagas Disease. Front. Immunol. 2021, 12, 4667. [Google Scholar] [CrossRef]
- Passos, L.S.A.; Villani, F.N.A.; Magalhães, L.M.D.; Gollob, K.J.; Antonelli, L.R.D.V.; Nunes, M.C.P.; Dutra, W.O. Blocking of CD1d Decreases Trypanosoma cruzi-Induced Activation of CD4-CD8-T Cells and Modulates the Inflammatory Response in Patients with Chagas Heart Disease. J. Infect. Dis. 2016, 214, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Dieter, J.A.; Rothaeusler, K.; Luo, Z.; Baumgarth, N. B-1 Cells in the Bone Marrow Are a Significant Source of Natural IgM. Eur. J. Immunol. 2012, 42, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Ma, Y.; Chen, X.; Liu, M.; Cai, Y.; Hu, X.; Xiang, D.; Nath, S.; Zhang, H.; Ye, H.; et al. Integrin CD11b Negatively Regulates BCR Signalling to Maintain Autoreactive B Cell Tolerance. Nat. Commun. 2013, 4, 2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X. Regulatory Functions of Innate-like B Cells. Cell. Mol. Immunol. 2013, 10, 113–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutra, W.O.; Profeta da Luz, Z.M.; Cançado, J.R.; Pereira, M.E.; Brígido-Nunes, R.M.; Galvão, L.M.C.; Colley, D.G.; Brener, Z.; Gazzinelli, G.; Carvalho-Parra, J.F. Influence of Parasite Presence on the Immunologic Profile of Peripheral Blood Mononuclear Cells from Chagasic Patients after Specific Drug Therapy. Parasite Immunol. 1996, 18, 579–585. [Google Scholar] [CrossRef] [Green Version]
- Cunha-Neto, E.; Dzau, V.J.; Allen, P.D.; Stamatiou, D.; Benvenutti, L.; Higuchi, M.L.; Koyama, N.S.; Silva, J.S.; Kalil, J.; Liew, C.C. Cardiac Gene Expression Profiling Provides Evidence for Cytokinopathy as a Molecular Mechanism in Chagas’ Disease Cardiomyopathy. Am. J. Pathol. 2005, 167, 305–313. [Google Scholar] [CrossRef]
- Teixeira, P.C.; Ducret, A.; Langen, H.; Nogoceke, E.; Santos, R.H.B.; Silva Nunes, J.P.; Benvenuti, L.; Levy, D.; Bydlowski, S.P.; Bocchi, E.A.; et al. Impairment of Multiple Mitochondrial Energy Metabolism Pathways in the Heart of Chagas Disease Cardiomyopathy Patients. Front. Immunol. 2021, 12, 4779. [Google Scholar] [CrossRef]
- Fonseca, S.G.; Reis, M.M.; Coelho, V.; Nogueira, L.G.; Monteiro, S.M.; Mairena, E.C.; Bacal, F.; Bocchi, E.; Guilherme, L.; Zheng, X.X.; et al. Locally Produced Survival Cytokines IL-15 and IL-7 May Be Associated to the Predominance of CD8+ T Cells at Heart Lesions of Human Chronic Chagas Disease Cardiomyopathy. Scand. J. Immunol. 2007, 66, 362–371. [Google Scholar] [CrossRef]
- Barretto, A.C.P.; Mady, C.; Arteage-Fernandez, E.; Stolf, N.; Lopes, E.A.; de Lourdes Higuchi, M.; Bellotti, G.; Pileggi, F. Right Ventricular Endomyocardial Biopsy in Chronic Chagas’ Disease. Am. Heart J. 1986, 111, 307–312. [Google Scholar] [CrossRef]
- Reis, M.M.; Higuchi, M.L.; Aiello, V.D.; Benvenuti, L.A. Growth Factors in the Myocardium of Patients with Chronic Chagasic Cardiomyopathy. Rev. Soc. Bras. Med. Trop. 2000, 33, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Argüello, R.J.; Vigliano, C.; Cabeza-Meckert, P.; Viotti, R.; Garelli, F.; Favaloro, L.E.; Favaloro, R.R.; Laguens, R.; Laucella, S.A. Presence of Antigen-Experienced T Cells with Low Grade of Differentiation and Proliferative Potential in Chronic Chagas Disease Myocarditis. PLoS Negl. Trop. Dis. 2014, 8, e2989. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.d.L.; Gutierrez, P.S.; Aiello, V.D.; Palomino, S.; Bocchi, E.; Kalil, J.; Bellotti, G.; Pileggi, F. Immunohistochemical Characterization of Infiltrating Cells in Human Chronic Chagasic Myocarditis: Comparison with Myocardial Rejection Process. Virchows Archiv. A 1993, 423, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Barry, M.; Bleackley, R.C. Cytotoxic T Lymphocytes: All Roads Lead to Death. Nat. Rev. Immunol. 2002, 2, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Dotiwala, F.; Mulik, S.; Polidoro, R.B.; Ansara, J.A.; Burleigh, B.A.; Walch, M.; Gazzinelli, R.T.; Lieberman, J. Killer Lymphocytes Use Granulysin, Perforin and Granzymes to Kill Intracellular Parasites. Nat. Med. 2016, 22, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Trapani, J.A.; Smyth, M.J. Functional Significance of the Perforin/Granzyme Cell Death Pathway. Nat. Rev. Immunol. 2002, 2, 735–747. [Google Scholar] [CrossRef]
- Ferraz, R.; Cunha, C.F.; Pimentel, M.I.F.; Lyra, M.R.; Pereira-Da-Silva, T.; Schubach, A.O.; Da-Cruz, A.M.; Bertho, A.L. CD3+CD4negCD8neg (Double Negative) T Lymphocytes and NKT Cells as the Main Cytotoxic-Related-CD107a+ Cells in Lesions of Cutaneous Leishmaniasis Caused by Leishmania (Viannia) Braziliensis. Parasites Vectors 2017, 10, 219. [Google Scholar] [CrossRef] [Green Version]
- Lasso, P.; Mateus, J.; Pavía, P.; Rosas, F.; Roa, N.; Thomas, M.C.; López, M.C.; González, J.M.; Puerta, C.J.; Cuéllar, A. Inhibitory Receptor Expression on CD8 + T Cells Is Linked to Functional Responses against Trypanosoma cruzi Antigens in Chronic Chagasic Patients. J. Immunol. 2015, 195, 3748–3758. [Google Scholar] [CrossRef] [Green Version]
- Menezes, C.A.S.; Sullivan, A.K.; Falta, M.T.; Mack, D.G.; Freed, B.M.; Rocha, M.O.C.; Gollob, K.J.; Fontenot, A.P.; Dutra, W.O. Highly Conserved CDR3 Region in Circulating CD4 +Vβ5 + T Cells May Be Associated with Cytotoxic Activity in Chagas Disease. Clin. Exp. Immunol. 2012, 169, 109–118. [Google Scholar] [CrossRef]
- Keesen, T.S.L.; Gomes, J.A.S.; Fares, R.C.G.; de Araújo, F.F.; Ferreira, K.S.; Chaves, A.T.; Rocha, M.O.C.; Correa-Oliveira, R. Characterization of CD4 + Cytotoxic Lymphocytes and Apoptosis Markers Induced by Trypanossoma Cruzi Infection. Scand. J. Immunol. 2012, 76, 311–319. [Google Scholar] [CrossRef]
- Dutra, W.O.; Martins-Filho, O.A.; Cançado, J.R.; Pinto-Dias, J.C.; Brener, Z.; Gazzinelli, G.; Carvalho, J.F.; Colley, D.G. Chagasic Patients Lack CD28 Expression on Many of Their Circulating T Lymphocytes. Scand. J. Immunol. 1996, 43, 88–93. [Google Scholar] [CrossRef]
- Giraldo, N.A.; Bolaños, N.I.; Cuellar, A.; Roa, N.; Cucunubá, Z.; Rosas, F.; Velasco, V.; Puerta, C.J.; González, J.M. T Lymphocytes from Chagasic Patients Are Activated but Lack Proliferative Capacity and Down-Regulate CD28 and CD3ζ. PLoS Negl. Trop. Dis. 2013, 7, e2038. [Google Scholar] [CrossRef] [PubMed]
- Laucella, S.; de Titto, E.H.; Segura, E.L.; Orn, A.; Rottenberg, M.E. Soluble Cell Adhesion Molecules in Human Chagas’ Disease: Association with Disease Severity and Stage of Infection. Am. J. Trop. Med. Hyg. 1996, 55, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Talvani, A.; Rocha, M.O.; Ribeiro, A.L.; Correa-Oliveira, R.; Teixeira, M.M. Chemokine Receptor Expression on the Surface of Peripheral Blood Mononuclear Cells in Chagas Disease. J. Infec. Dis. 2004, 189, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Neto, E.; Chevillard, C. Chagas Disease Cardiomyopathy: Immunopathology and Genetics. Mediat. Inflamm. 2014, 2014, 683230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flórez, O.; Martín, J.; González, C.I. Genetic Variants in the Chemokines and Chemokine Receptors in Chagas Disease. Hum. Immunol. 2012, 73, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Knight, J. Polymorphisms in Tumor Necrosis Factor and Other Cytokines as Risks for Infectious Diseases and the Septic Syndrome. Curr. Infect. Dis. Rep. 2001, 3, 427–439. [Google Scholar] [CrossRef]
- Costa, G.C.; Rocha, M.O.D.C.; Moreira, P.R.; Menezes, C.A.S.; Silva, M.R.; Gollob, K.J.; Dutra, W.O. Functional IL-10 Gene Polymorphism Is Associated with Chagas Disease Cardiomyopathy. J. Infect. Dis. 2009, 199, 451–454. [Google Scholar] [CrossRef] [Green Version]
- Deghaide, N.H.S.; Dantas, R.O.; Donadi, E.A. HLA Class I and II Profiles of Patients Presenting with Chagas’ Disease. Dig. Dis. Sci. 1998, 43, 246–252. [Google Scholar] [CrossRef]
- Alvarado-Arnez, L.E.; Batista, A.M.; Alves, S.M.; Melo, G.; Maria, V.; de Lorena, B.; Cardoso, C.C.; Pereira, I.R.; Carrazzone, C.; Pacheco, A.G.; et al. Single Nucleotide Polymorphisms of Cytokine-Related Genes and Association with Clinical Outcome in a Chagas Disease Case-Control Study from Brazil. Mem. Inst. Oswaldo Cruz 2018, 113, e170489. [Google Scholar] [CrossRef]
- Tajima, F. The Effect of Change in Population Size on DNA Polymorphism. Genetics 1989, 123, 597–601. [Google Scholar] [CrossRef]
- Arnez, L.E.A.; Venegas, E.N.; Ober, C.; Thompson, E.E. Sequence Variation in the IL4 Gene and Resistance to Trypanosoma cruzi Infection in Bolivians. J. Allergy Clin. Immunol. 2011, 127, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Ramasawmy, R.; Faé, K.C.; Cunha-Neto, E.; Borba, S.C.P.; Ianni, B.; Mady, C.; Goldberg, A.C.; Kalil, J. Variants in the Promoter Region of IKBL/NFKBIL1 Gene May Mark Susceptibility to the Development of Chronic Chagas’ Cardiomyopathy among Trypanosoma cruzi-Infected Individuals. Mol. Immunol. 2008, 45, 283–288. [Google Scholar] [CrossRef]
- Frade-Barros, A.F.; Ianni, B.M.; Cabantous, S.; Pissetti, C.W.; Saba, B.; Lin-Wang, H.T.; Buck, P.; Marin-Neto, J.A.; Schmidt, A.; Dias, F.; et al. Polymorphisms in Genes Affecting Interferon-γ Production and Th1 T Cell Differentiation Are Associated with Progression to Chagas Disease Cardiomyopathy. Front. Immunol. 2020, 11, 1386. [Google Scholar] [CrossRef] [PubMed]
- Reis, P.G.; Ayo, C.M.; de Mattos, L.C.; de Mattos, C.D.C.B.; Sakita, K.M.; de Moraes, A.G.; Muller, L.P.; Aquino, J.S.; Macedo, L.C.; Mazini, P.S.; et al. Genetic Polymorphisms of IL17 and Chagas Disease in the South and Southeast of Brazil. J. Immunol. Res. 2017, 2017, 1017621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, D.A.L.; Echeverría, L.E.; González, C.I.; Martin, J. Investigation of the Role of IL17A Gene Variants in Chagas Disease. Genes Immun. 2015, 16, 536–540. [Google Scholar] [CrossRef] [Green Version]
- Strauss, M.; Palma-Vega, M.; Casares-Marfil, D.; Bosch-Nicolau, P.; lo Presti, M.S.; Molina, I.; González, C.I.; Paglini, P.A.; Schijman, A.G.; Robello, C.; et al. Genetic Polymorphisms of IL17A Associated with Chagas Disease: Results from a Meta-Analysis in Latin American Populations. Sci. Rep. 2020, 10, 5015. [Google Scholar] [CrossRef] [Green Version]
- Clipman, S.J.; Henderson-Frost, J.; Fu, K.Y.; Bern, C.; Flores, J.; Gilman, R.H. Genetic Association Study of NLRP1, CARD, and CASP1 Inflammasome Genes with Chronic Chagas Cardiomyopathy among Trypanosoma cruzi Seropositive Patients in Bolivia. PLoS ONE 2018, 13, e0192378. [Google Scholar] [CrossRef] [Green Version]
- Fu, K.Y.J.; Zamudio, R.; Frost, J.H.; Almuedo, A.; Steinberg, H.; Clipman, S.J.; Duran, G.; Marcus, R.; Crawford, T.; Alyesh, D.; et al. Association of Caspase-1 Polymorphisms with Chagas Cardiomyopathy among Individuals in Santa Cruz, Bolivia. Rev. Soc. Bras. Med. Trop. 2017, 50, 516–523. [Google Scholar] [CrossRef] [Green Version]
- Pissetti, C.W.; de Oliveira, R.F.; Correia, D.; Nascentes, G.A.N.; Llaguno, M.M.; Rodrigues, V. Association between the Lymphotoxin-Alpha Gene Polymorphism and Chagasic Cardiopathy. J. Interferon Cytokine Res. 2013, 33, 130–135. [Google Scholar] [CrossRef]
- Ramasawmy, R.; Fae, K.C.; Cunha-Neto, E.; Müller, N.G.; Cavalcanti, V.L.; Ferreira, R.C.; Drigo, S.A.; Ianni, B.; Mady, C.; Goldberg, A.C.; et al. Polymorphisms in the Gene for Lymphotoxin-Alpha Predispose to Chronic Chagas Cardiomyopathy. J. Infect. Dis. 2007, 196, 1836–1843. [Google Scholar] [CrossRef]
- Silva, M.C.; da Silva Medina, T.; Fuzo, C.A.; Dias, F.C.; Freitas-Castro, F.; Fukutani, K.F.; Donadi, E.A.; Cunha-Neto, E.; Cunha, T.M.; Silva, J.S. Polymorphism in the Catalytic Subunit of the PI3Kγ Gene Is Associated with Trypanosoma cruzi-Induced Chronic Chagasic Cardiomyopathy. Infect. Genet. Evol. 2021, 88, 104671. [Google Scholar] [CrossRef] [PubMed]
- Dias, F.C.; Medina, T.d.S.; Mendes-Junior, C.T.; Dantas, R.O.; Pissetti, C.W.; Rodrigues Junior, V.; Dellalibera-Joviliano, R.; Marin-Neto, J.A.; Gutierrez, F.R.S.; Moreau, P.; et al. Polymorphic Sites at the Immunoregulatory CTLA-4 Gene Are Associated with Chronic Chagas Disease and Its Clinical Manifestations. PLoS ONE 2013, 8, e78367. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, M.V.; da Costa Rocha, M.O.; de Sousa, G.R.; Nunes, M.C.P.; Gollob, K.J.; Dutra, W.O.; da Silva Menezes, C.A. Low Levels of Vasoactive Intestinal Peptide Are Associated with Chagas Disease Cardiomyopathy. Hum. Immunol. 2013, 74, 1375–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flórez, O.; Martín, J.; González Rugeles, C.I. Interleukin 4, Interleukin 4 Receptor-α and Interleukin 10 Gene Polymorphisms in Chagas Disease. Parasite Immunol. 2011, 33, 506–511. [Google Scholar] [CrossRef]
- Torres, O.A.; Calzada, J.E.; Beraún, Y.; Morillo, C.A.; González, C.I.; González, A.; Martín, J. Association of the Macrophage Migration Inhibitory Factor −173G/C Polymorphism with Chagas Disease. Hum. Immunol. 2009, 70, 543–546. [Google Scholar] [CrossRef]
- Torres, O.A.; Calzada, J.E.; Beraún, Y.; Morillo, C.A.; González, A.; González, C.I.; Martín, J. Lack of Association between IL-6 -174G/C Gene Polymorphism and Chagas Disease. Tissue Antigens 2010, 76, 131–134. [Google Scholar] [CrossRef]
- Calzada, J.E.; Beraún, Y.; González, C.I.; Martín, J. Transforming Growth Factor Beta 1 (TGFβ1) Gene Polymorphisms and Chagas Disease Susceptibility in Peruvian and Colombian Patients. Cytokine 2009, 45, 149–153. [Google Scholar] [CrossRef]
- Ferreira, R.R.; da Silva Madeira, F.; Alves, G.F.; da Costa Chambela, M.; de Oliveira Vaz Curvo, E.; Moreira, A.D.S.; de Sá, R.A.; Mendonça-Lima, L.; Cabello, P.H.; Bailly, S.; et al. TGF-β Polymorphisms Are a Risk Factor for Chagas Disease. Dis. Markers 2018, 2018, 4579198. [Google Scholar] [CrossRef] [Green Version]
- Zafra, G.; Flórez, O.; Morillo, C.A.; Echeverría, L.E.; Martín, J.; González, C.I. Polymorphisms of Toll-like Receptor 2 and 4 Genes in Chagas Disease. Mem. Inst. Oswaldo Cruz 2008, 103, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Weitzel, T.; Zulantay, I.; Danquah, I.; Hamann, L.; Schumann, R.R.; Apt, W.; Mockenhaupt, F.P. Mannose-Binding Lectin and Toll-like Receptor Polymorphisms and Chagas Disease in Chile. Am. J. Trop. Med. Hyg. 2012, 86, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Cruz, G.; Angelo, A.L.D.; Larocca, T.F.; Macedo, C.T.; Noya-Rabelo, M.; Correia, L.C.L.; Torreão, J.A.; de Freitas Souza, B.S.; dos Santos, R.R.; Soares, M.B.P. Assessment of Galectin-3 Polymorphism in Subjects with Chronic Chagas Disease. Arq. Bras. Cardiol. 2015, 105, 472–478. [Google Scholar] [CrossRef]
- Rodriguez, D.A.L.; González, C.I.; Martin, J. Analysis of Association of FOXO3 Gene with Trypanosoma cruzi Infection and Chronic Chagasic Cardiomyopathy. HLA 2016, 87, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Faé, K.C.; Drigo, S.A.; Cunha-Neto, E.; Ianni, B.; Mady, C.; Kalil, J.; Goldberg, A.C. HLA and β-Myosin Heavy Chain Do Not Influence Susceptibility to Chagas’ Disease Cardiomyopathy. Microbes Infect. 2000, 2, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Layrisse, Z.; Fernandez, M.T.; Montagnani, S.; Matos, M.; Balbas, O.; Herrera, F.; Colorado, I.A.; Catalioti, F.; Acquatella, H. HLA-C*03 Is a Risk Factor for Cardiomyopathy in Chagas Disease. Hum. Immunol. 2000, 61, 925–929. [Google Scholar] [CrossRef] [PubMed]
- Nieto, A.; Beraún, Y.; Callado, M.D.; Caballero, A.; Alonso, A.; González, A.; Martín, J. HLA Haplotypes Are Associated with Differential Susceptibility to Trypanosoma Cruzi Infection. Tissue Antigens 2000, 55, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Del Puerto, F.; Nishizawa, J.E.; Kikuchi, M.; Roca, Y.; Avilas, C.; Gianella, A.; Lora, J.; Velarde, F.U.G.; Miura, S.; Komiya, N.; et al. Protective Human Leucocyte Antigen Haplotype, HLA-DRB1*01-B*14, against Chronic Chagas Disease in Bolivia. PLoS Negl. Trop. Dis. 2012, 6, e1587. [Google Scholar] [CrossRef]
- De Oliveira, A.P.; Bernardo, C.R.; da Silveira Camargo, A.V.; Ronchi, L.S.; Borim, A.A.; de Mattos, C.C.B.; de Campos, E.; Castiglioni, L.; Netinho, J.G.; Cavasini, C.E.; et al. Genetic Susceptibility to Cardiac and Digestive Clinical Forms of Chronic Chagas Disease: Involvement of the CCR5 59029 A/G Polymorphism. PLoS ONE 2015, 10, e0141847. [Google Scholar] [CrossRef] [Green Version]
- Machuca, M.A.; Suárez, E.U.; Echeverría, L.E.; Martín, J.; González, C.I. SNP/Haplotype Associations of CCR2 and CCR5 Genes with Severity of Chagasic Cardiomyopathy. Hum. Immunol. 2014, 75, 1210–1215. [Google Scholar] [CrossRef]
- Fernández-Mestre, M.T.; Montagnani, S.; Layrisse, Z. Is the CCR5-59029-G/G Genotype a Protective Factor for Cardiomyopathy in Chagas Disease? Hum. Immunol. 2004, 65, 725–728. [Google Scholar] [CrossRef]
- Calzada, J.E.; Nieto, A.; Beraún, Y.; Martín, J. Chemokine Receptor CCR5 Polymorphisms and Chagas’ Disease Cardiomyopathy. Tissue Antigens 2001, 58, 154–158. [Google Scholar] [CrossRef]
- Cruz-Robles, D.; Chvez-Gonzlez, J.P.; Cavazos-Quero, M.M.; Prez-Mndez, O.; Reyes, P.A.; Vargas-Alarcn, G. Association between IL-1B and IL-1RN Gene Polymorphisms and Chagas’ Disease Development Susceptibility. Immunol. Investig. 2009, 38, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Flórez, O.; Zafra, G.; Morillo, C.; Martín, J.; González, C.I. Interleukin-1 Gene Cluster Polymorphism in Chagas Disease in a Colombian Case-Control Study. Hum. Immunol. 2006, 67, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Torres, O.A.; Calzada, J.E.; Beraún, Y.; Morillo, C.A.; González, A.; González, C.I.; Martín, J. Role of the IFNG +874T/A Polymorphism in Chagas Disease in a Colombian Population. Infect. Genet. Evol. 2010, 10, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Beraún, Y.; Nieto, A.; Collado, M.D.; González, A.; Martín, J. Polymorphisms at Tumor Necrosis Factor (TNF) Loci Are Not Associated with Chagas’ Disease. Tissue Antigens 1998, 52, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Drigo, S.A.; Cunha-Neto, E.; Ianni, B.; Mady, C.; Faé, K.C.; Buck, P.; Kalil, J.; Goldberg, A.C. Lack of Association of Tumor Necrosis Factor-Alpha Polymorphisms with Chagas Disease in Brazilian Patients. Immunol. Lett. 2007, 108, 109–111. [Google Scholar] [CrossRef]
- Pissetti, C.W.; Correia, D.; de Oliveira, R.F.; Llaguno, M.M.; Balarin, M.A.S.; Silva-Grecco, R.L.; Rodrigues, V. Genetic and Functional Role of TNF-Alpha in the Development Trypanosoma Cruzi Infection. PLoS Negl. Trop. Dis. 2011, 5, e976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Criado, L.; Flórez, O.; Martín, J.; González, C.I. Genetic Polymorphisms in TNFA/TNFR2 Genes and Chagas Disease in a Colombian Endemic Population. Cytokine 2012, 57, 398–401. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Pérez, J.M.; Cruz-Robles, D.; Hernández-Pacheco, G.; Pérez-Hernández, N.; Murguía, L.E.; Granados, J.; Reyes, P.A.; Vargas-Alarcón, G. Tumor Necrosis Factor-Alpha Promoter Polymorphism in Mexican Patients with Chagas’ Disease. Immunol. Lett. 2005, 98, 97–102. [Google Scholar] [CrossRef]
- Ramasawmy, R.; Cunha-Neto, E.; Fae, K.C.; Borba, S.C.P.; Teixeira, P.C.; Ferreira, S.C.P.; Goldberg, A.C.; Lanni, B.; Mady, C.; Kalil, J. Heterozygosity for the S180L Variant of MAL/TIRAP, a Gene Expressing an Adaptor Protein in the Toll-like Receptor Pathway, Is Associated with Lower Risk of Developing Chronic Chagas Cardiomyopathy. J. Infect. Dis. 2009, 199, 1838–1845. [Google Scholar] [CrossRef] [Green Version]
- Frade, A.F.; Pissetti, C.W.; Ianni, B.M.; Saba, B.; Lin-Wang, H.T.; Nogueira, L.G.; de Melo Borges, A.; Buck, P.; Dias, F.; Baron, M.; et al. Genetic Susceptibility to Chagas Disease Cardiomyopathy: Involvement of Several Genes of the Innate Immunity and Chemokine-Dependent Migration Pathways. BMC Infect. Dis. 2013, 13, 587. [Google Scholar] [CrossRef]
- Ramasawmy, R.; Cunha-Neto, E.; Faé, K.C.; Müller, N.G.; Cavalcanti, V.L.; a Drigo, S.; Ianni, B.; Mady, C.; Kalil, J.; Goldberg, A.C. BAT1, a Putative Anti-Inflammatory Gene, Is Associated with Chronic Chagas Cardiomyopathy. J. Infect. Dis. 2006, 193, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Ramasawmy, R.; Cunha-Neto, E.; Faé, K.C.; Martello, F.G.; Müller, N.G.; Cavalcanti, V.L.; Iaani, B.; Mady, C.; Kalil, J.; Goldberg, A.C. The Monocyte Chemoattractant Protein-1 Gene Polymorphism Is Associated with Cardiomyopathy in Human Chagas Disease. Clin. Infect. Dis. 2006, 43, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Peschansky, V.J.; Wahlestedt, C. Non-Coding RNAs as Direct and Indirect Modulators of Epigenetic Regulation. Epigenetics 2014, 9, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinberg, A.P. The Key Role of Epigenetics in Human Disease Prevention and Mitigation. N. Engl. J. Med. 2018, 378, 1323–1334. [Google Scholar] [CrossRef]
- Saxonov, S.; Berg, P.; Brutlag, D.L. A Genome-Wide Analysis of CpG Dinucleotides in the Human Genome Distinguishes Two Distinct Classes of Promoters. Proc. Natl. Acad. Sci. USA 2006, 103, 1412–1417. [Google Scholar] [CrossRef] [Green Version]
- McMahon, K.W.; Karunasena, E.; Ahuja, N. The Roles of DNA Methylation in the Stages of Cancer. Cancer J. 2017, 23, 257–261. [Google Scholar] [CrossRef]
- Alaskhar Alhamwe, B.; Khalaila, R.; Wolf, J.; Bülow, V.; Harb, H.; Alhamdan, F.; Hii, C.S.; Prescott, S.L.; Ferrante, A.; Renz, H.; et al. Histone Modifications and Their Role in Epigenetics of Atopy and Allergic Diseases. Allergy Asthma Clin. Immunol. 2018, 14, 39. [Google Scholar] [CrossRef] [Green Version]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell. Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Esteller, M. Epigenetics in Cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef]
- Rossetto, D.; Avvakumov, N.; Côté, J. Histone Phosphorylation: A Chromatin Modification Involved in Diverse Nuclear Events. Epigenetics 2012, 7, 1098–1108. [Google Scholar] [CrossRef] [Green Version]
- Frías-Lasserre, D.; Villagra, C.A. The Importance of NcRNAs as Epigenetic Mechanisms in Phenotypic Variation and Organic Evolution. Front. Microbiol. 2017, 8, 2483. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.C.; Sharma, A.; Mehta, R.; Paul, S. Genome Wide Computational Identification of Tuna (Thunnus Orientalis) MicroRNAs and Their Targets. Ocean. Sci. J. 2018, 53, 727–734. [Google Scholar] [CrossRef]
- Miska, E.A. How MicroRNAs Control Cell Division, Differentiation and Death. Curr. Opin. Genet. Dev. 2005, 15, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Alemohammad, H.; Najafzadeh, B.; Asadzadeh, Z.; Baghbanzadeh, A.; Ghorbaninezhad, F.; Najafzadeh, A.; Safarpour, H.; Bernardini, R.; Brunetti, O.; Sonnessa, M.; et al. The Importance of Immune Checkpoints in Immune Monitoring: A Future Paradigm Shift in the Treatment of Cancer. Biomed. Pharmacother. 2022, 146, 112516. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.R.P.; Frade, A.F.; Santos, R.H.B.; Teixeira, P.C.; Baron, M.A.; Navarro, I.C.; Benvenuti, L.A.; Fiorelli, A.I.; Bocchi, E.A.; Stolf, N.A.; et al. MicroRNAs MiR-1, MiR-133a, MiR-133b, MiR-208a and MiR-208b Are Dysregulated in Chronic Chagas Disease Cardiomyopathy. Int. J. Cardiol. 2014, 175, 409–417. [Google Scholar] [CrossRef]
- Linhares-Lacerda, L.; Granato, A.; Gomes-Neto, J.F.; Conde, L.; Freire-de-Lima, L.; de Freitas, E.O.; Freire-de-Lima, C.G.; Barroso, S.P.C.; Guerra, R.J.d.A.; Pedrosa, R.C.; et al. Circulating Plasma MicroRNA-208a as Potential Biomarker of Chronic Indeterminate Phase of Chagas Disease. Front. Microbiol. 2018, 9, 269. [Google Scholar] [CrossRef]
- Nonaka, C.K.V.; Macêdo, C.T.; Cavalcante, B.R.R.; de Alcântara, A.C.; Silva, D.N.; da Rocha Bezerra, M.; Caria, A.C.I.; Tavora, F.R.F.; de Souza Neto, J.D.; Noya-Rabelo, M.M.; et al. Circulating MiRNAs as Potential Biomarkers Associated with Cardiac Remodeling and Fibrosis in Chagas Disease Cardiomyopathy. Int. J. Mol. Sci. 2019, 20, 4064. [Google Scholar] [CrossRef] [Green Version]
- Navarro, I.C.; Ferreira, F.M.; Nakaya, H.I.; Baron, M.A.; Vilar-Pereira, G.; Pereira, I.R.; Silva, A.M.G.; Real, J.M.; de Brito, T.; Chevillard, C.; et al. MicroRNA Transcriptome Profiling in Heart of Trypanosoma cruzi-Infected Mice: Parasitological and Cardiological Outcomes. PLoS Negl. Trop. Dis. 2015, 9, 3828. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, L.R.P.; Ferreira, F.M.; Laugier, L.; Cabantous, S.; Navarro, I.C.; Cândido, D.D.S.; Rigaud, V.C.; Real, J.M.; Pereira, G.V.; Pereira, I.R.; et al. Integration of MiRNA and Gene Expression Profiles Suggest a Role for MiRNAs in the Pathobiological Processes of Acute Trypanosoma Cruzi Infection. Sci. Rep. 2017, 7, 17990. [Google Scholar] [CrossRef] [Green Version]
- Ballinas-Verdugo, M.A.; Jiménez-Ortega, R.F.; Martínez-Martínez, E.; Rivas, N.; Contreras-López, E.A.; Carbó, R.; Sánchez, F.; Bojalil, R.; Márquez-Velasco, R.; Sánchez-Muñoz, F.; et al. Circulating MiR-146a as a Possible Candidate Biomarker in the Indeterminate Phase of Chagas Disease. Biol. Res. 2021, 54, 21. [Google Scholar] [CrossRef]
- Nonaka, C.K.V.; Sampaio, G.L.; Silva, K.N.; Khouri, R.; Macedo, C.T.; Consortium, C.T.R.; Rogatto, S.R.; dos Santos, R.R.; de Freitas Souza, B.S.; Soares, M.B.P. Therapeutic Mir-21 Silencing Reduces Cardiac Fibrosis and Modulates Inflammatory Response in Chronic Chagas Disease. Int. J. Mol. Sci. 2021, 22, 3307. [Google Scholar] [CrossRef] [PubMed]
- Jha, B.K.; Varikuti, S.; Seidler, G.R.; Volpedo, G.; Satoskar, A.R.; McGwire, B.S. MicroRNA-155 Deficiency Exacerbates Trypanosoma cruzi Infection. Infect. Immun. 2020, 88, e00948–e01019. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stark, G.R.; Cheon, H.; Wang, Y. Responses to Cytokines and Interferons That Depend upon JAKs and STATs. Cold Spring Harb. Perspect. Biol. 2018, 10, a028555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, H.; Tanaka, T.; Shi, W.; Matsumoto, M.; Minami, M.; Kashiwamura, S.I.; Nakanishi, K.; Yoshida, N.; Kishimoto, T.; Akira, S. Essential Role of Stat6 in IL-4 Signalling. Nature 1996, 380, 627–630. [Google Scholar] [CrossRef]
- Choudhuri, S.; Garg, N.J. PARP1-CGAS-NF-ΚB Pathway of Proinflammatory Macrophage Activation by Extracellular Vesicles Released during Trypanosoma cruzi Infection and Chagas Disease. PLoS Pathog. 2020, 16, e1008474. [Google Scholar] [CrossRef] [Green Version]
- Campos-Estrada, C.; Liempi, A.; González-Herrera, F.; Lapier, M.; Kemmerling, U.; Pesce, B.; Ferreira, J.; López-Muñoz, R.; Maya, J.D. Simvastatin and Benznidazole-Mediated Prevention of Trypanosoma cruzi-Induced Endothelial Activation: Role of 15-Epi-Lipoxin A4 in the Action of Simvastatin. PLoS Negl. Trop. Dis. 2015, 9, e0003770. [Google Scholar] [CrossRef]
- Cevey, Á.C.; Mirkin, G.A.; Penas, F.N.; Goren, N.B. Low-Dose Benznidazole Treatment Results in Parasite Clearance and Attenuates Heart Inflammatory Reaction in an Experimental Model of Infection with a Highly Virulent Trypanosoma cruzi Strain. Int. J. Parasitol. Drugs Drug Resist. 2015, 6, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Wan, X.; Wen, J.J.; Koo, S.J.; Liang, L.Y.; Garg, N.J. SIRT1-PGC1α-NFκB Pathway of Oxidative and Inflammatory Stress during Trypanosoma Cruzi Infection: Benefits of SIRT1-Targeted Therapy in Improving Heart Function in Chagas Disease. PLoS Pathog. 2016, 12, e1005954. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cytokines and Their Receptors | Role | Source | References |
| Association withthe Development of CCC | |||
| IFN-gamma | Inflammation, T-cell activation | PBMC, heart | [22,24,29,30,37,38,39] |
| TNF | Inflammation, T-cell activation, worse cardiac function | Plasma, blood, PBMC, heart | [18,28,31,32,38,40,41,42] |
| sTNFR1 and sTNFR2 | Inflammation, worse cardiac function | Plasma | [43] |
| IL-6 | Inflammation, worse cardiac function | Plasma | [30,44,45] |
| IL-1β | Inflammation | Plasma | [30,46] |
| TGF-β | Tissue fibrosis | Serum, heart | [47] |
| MIF | Inflammation, progression | Serum | [48] |
| Association with protection to CCC | |||
| IL-2 | Low in CCC | PBMC | [49] |
| IL-10 | Protective immune response, better cardiac function | Plasma, PBMC, whole blood | [18,30,38,39,40,41,50] |
| IL-17 | Protective immune response, better cardiac function | Plasma, PBMC | [38,51,52,53] |
| Fibrotic and Cytotoxic Molecules | Description | Source | References |
| Association withthe Development of CCC | |||
| MMP-2 | Tissue fibrosis | Plasma | [54,55] |
| MMP-9 | Tissue fibrosis | Plasma, PBMC | [54,56] |
| MMP-2/MMP-9 ratio | Pathological cardiac remodeling | Plasma, heart | [57,58] |
| Fibronectin | Recruitment of inflammatory T cells to the heart | Heart | [42] |
| Granzyme A | Myocyte dysfunction | Heart | [28] |
| Chemokines and Their Receptors | Description | Source | References |
| Association withthe Development of CCC | |||
| CXCL9 | Recruitment of inflammatory cells | Plasma, heart | [46,59,60] |
| CXCL10 | Recruitment of inflammatory cells | Plasma | [46,61,62] |
| CCL5 | Recruitment of inflammatory cells | Serum | [63] |
| CCL2 | Recruitment of inflammatory cells | Serum and plasma | [32,63] |
| CCR5 | Recruitment of inflammatory cells, Worse prognosis | PBMC | [62,64] |
| CXCR3 | Recruitment of inflammatory cells | PBMC | [64] |
| Gene Polymorphism | Description | References |
| Association with Development of CCC | ||
| IL4RA | Interleukin 4 receptor | [101] |
| IKBL/NFKBIL1 | NF-kappa-B inhibitor-like protein 1 | [102] |
| IL12ß; IL12 | Pro-inflammatory cytokine | [99,103] |
| IL17A; IL17F | Protective cytokine | [104,105,106] |
| NLRP1 | Protein involved in inflammasome | [107] |
| CASP1 | Protein involved in inflammatory cascade | [108] |
| Lymphotoxin | Member of the TNF superfamily of cytokines; are responsible for regulating the growth and function of lymphocytes | [109,110] |
| PI3 kgamma | Molecules involved in signaling pathway of the efficient immune response against T. cruzi | [111] |
| Association with Protection to CCC | ||
| CXCL10, CCL5, CXCL9 | Chemokine Ligand | [60,63] |
| CTLA-4 | Cytotoxic T-lymphocyte-associated antigen 4 | [112] |
| VPAC1 | Vasoactive intestinal peptide (VIP) receptors 1 | [113] |
| No association with CCC | ||
| IL4 | Anti-inflammatory cytokine | [101,103,114] |
| MIF | Macrophage migration inhibitory factor | [115] |
| IL1A, IL6 | Pro-inflammatory cytokine | [103,116] |
| TGF-β1 | Multifunctional cytokine | [99,117,118] |
| TLR1, TLR2 TLR4, TLR6 | Toll-like receptor (TLR) | [119,120] |
| TNFR1; TNFR2; | Tumor necrosis factor receptor | [99] |
| Galectin-3 | Member of the lectin family/ cell–cell adhesion | [121] |
| CARD11 | Protein involved in the function of immune system cells | [107] |
| FOXP3 | Protein involved in immune system responses | [122] |
| Variable According to the Population Studied | ||
| MHC genes | Major histocompatibility complex/presentation of internal or external antigens to the T cells | [123,124,125,126] |
| CCR5; CCR2 | Chemokine receptor type | [60,109,127,128,129,130] |
| TNFA; TNFB; IL1B; IFN-g | Pro-inflammatory cytokine | [99,103,131,132,133,134,135,136,137,138] |
| IL10 | Anti-inflammatory cytokine | [97,99,114] |
| IL1RN | Interleukin-1 receptor antagonist | [131,132] |
| MAL/TIRAP | Encodes an adaptor protein for TLR | [120,139,140] |
| BAT-1 | Anti-inflammatory activity associated with reduced expression of HLA-B-1 | [99,141] |
| CCL2/MCP-1 | Chemokine ligand 2 | [63,142] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koh, C.C.; Neves, E.G.A.; de Souza-Silva, T.G.; Carvalho, A.C.; Pinto, C.H.R.; Galdino, A.S.; Gollob, K.J.; Dutra, W.O. Cytokine Networks as Targets for Preventing and Controlling Chagas Heart Disease. Pathogens 2023, 12, 171. https://doi.org/10.3390/pathogens12020171
Koh CC, Neves EGA, de Souza-Silva TG, Carvalho AC, Pinto CHR, Galdino AS, Gollob KJ, Dutra WO. Cytokine Networks as Targets for Preventing and Controlling Chagas Heart Disease. Pathogens. 2023; 12(2):171. https://doi.org/10.3390/pathogens12020171
Chicago/Turabian StyleKoh, Carolina Cattoni, Eula G. A. Neves, Thaiany Goulart de Souza-Silva, Ana Carolina Carvalho, Cecília Horta Ramalho Pinto, Alexsandro Sobreira Galdino, Kenneth J. Gollob, and Walderez Ornelas Dutra. 2023. "Cytokine Networks as Targets for Preventing and Controlling Chagas Heart Disease" Pathogens 12, no. 2: 171. https://doi.org/10.3390/pathogens12020171