
Impact of MgtC on the Fitness of Yersinia pseudotuberculosis

Abstract
:1. Introduction
2. Material and Method
2.1. Strains, Plasmids, Macrophages and Culture Conditions
2.2. Plasmid Construction
2.3. Strain Construction
2.4. Determination of Bacterial Growth Curve in High or Low Mg2+ Minimal Medium
2.5. RNA Extraction and Real-Time PCR
2.6. Bacterial Intracellular Survival Assay
2.7. Lactate Dehydrogenase (LDH) Activity Assay
2.8. RNA-Seq
2.9. Animal Infection
2.10. Statistical Analysis
3. Results
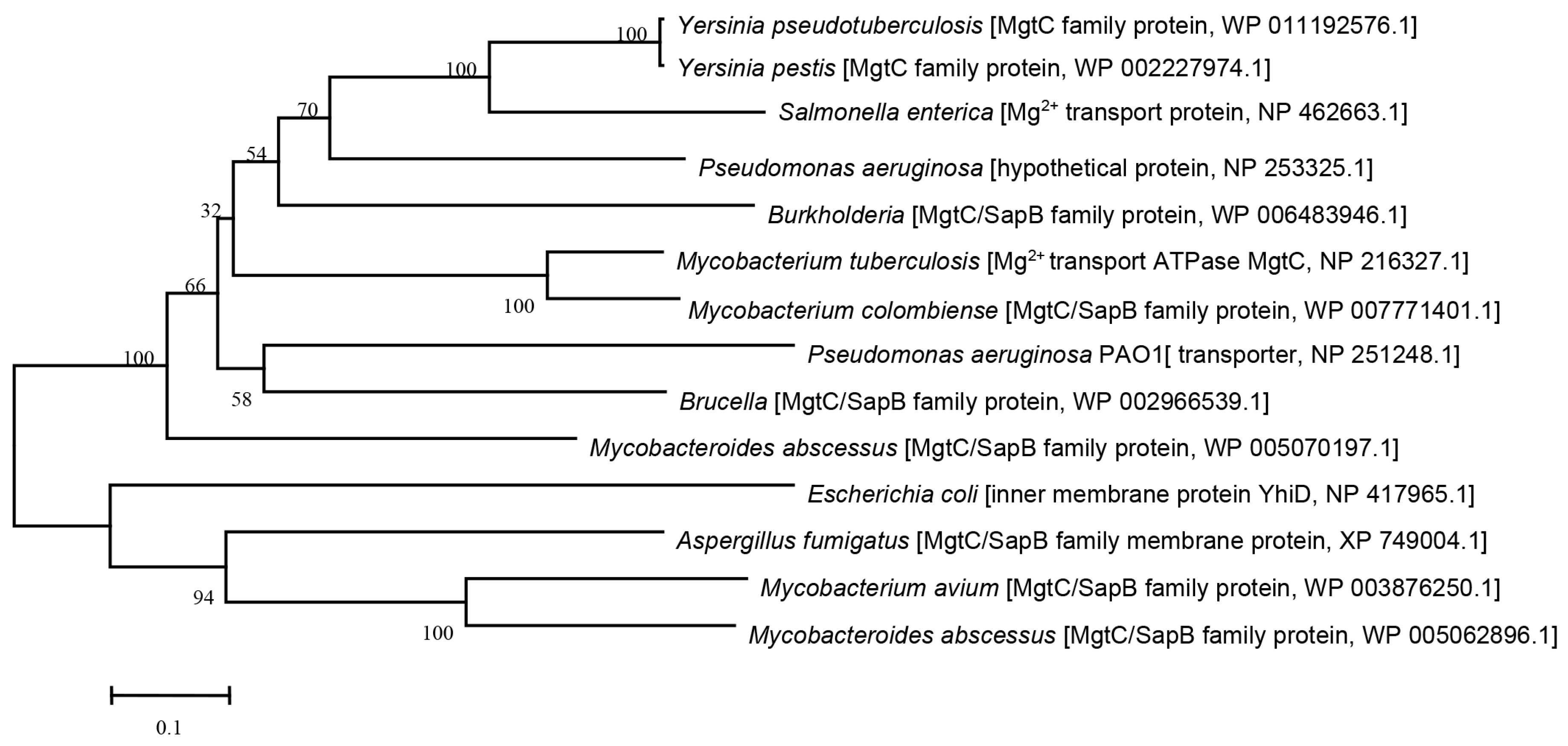
3.1. Phylogenetic Analysis of MgtC-like Proteins in Different Bacteria
3.2. MgtC Is Required for Y. pseudotuberculosis Growth under Mg2+ Starvation and mtgC Transcription Is Strongly Induced under Mg2+ Starvation and Intracellular Conditions
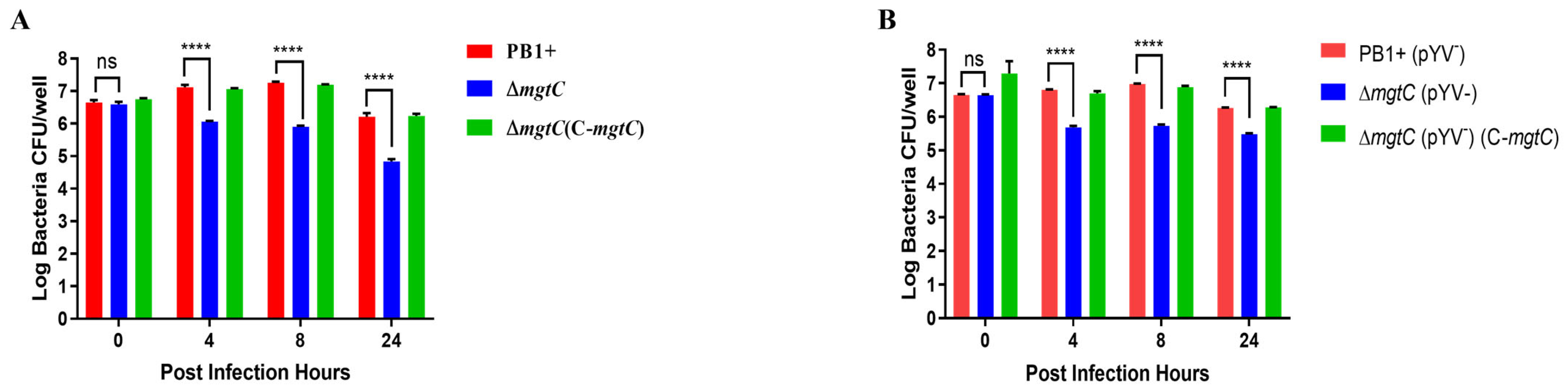
3.3. MgtC Plays a Critical Role for Y. pseudotuberculosis Replication in Macrophages
3.4. Genes Analysis of mgtC Mutant within Macrophages by RNA-Seq
3.5. Pathogenicity of the ΔmgtC Mutant In Vivo
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fukushima, H.; Gomyoda, M.; Ishikura, S.; Nishio, T.; Moriki, S.; Endo, J.; Kaneko, S.; Tsubokura, M. Cat-contaminated environmental substances lead to Yersinia pseudotuberculosis infection in children. J. Clin. Microbiol. 1989, 27, 2706–2709. [Google Scholar] [CrossRef]
- Laukkanen, R.; Martinez, P.O.; Siekkinen, K.M.; Ranta, J.; Maijala, R.; Korkeala, H. Transmission of Yersinia pseudotuberculosis in the pork production chain from farm to slaughterhouse. Appl. Environ. Microbiol. 2008, 74, 5444–5450. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Hayashidani, H.; Okabe, N.; Une, Y. Aberrant forms of Yersinia pseudotuberculosis as spheroplasts and filaments in yersiniosis in squirrel monkeys. Vet. Pathol. 2015, 52, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Dube, P. Interaction of Yersinia with the gut: Mechanisms of pathogenesis and immune evasion. Curr. Top. Microbiol. Immunol. 2009, 337, 61–91. [Google Scholar] [PubMed]
- Singh, A.K.; Curtiss, R., 3rd; Sun, W. A Recombinant Attenuated Yersinia pseudotuberculosis Vaccine Delivering a Y. pestis YopENt138-LcrV Fusion Elicits Broad Protection against Plague and Yersiniosis in Mice. Infect. Immun. 2019, 87, 00296-19. [Google Scholar] [CrossRef] [PubMed]
- Grabenstein, J.P.; Marceau, M.; Pujol, C.; Simonet, M.; Bliska, J.B. The response regulator PhoP of Yersinia pseudotuberculosis is important for replication in macrophages and for virulence. Infect. Immun. 2004, 72, 4973–4984. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.D.; Finlay, B.B. Isolation and characterization of Salmonella typhimurium and Yersinia pseudotuberculosis-containing phagosomes from infected mouse macrophages: Y. pseudotuberculosis traffics to terminal lysosomes where they are degraded. Eur. J. Cell Biol. 1998, 77, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Pujol, C.; Bliska, J.B. The ability to replicate in macrophages is conserved between Yersinia pestis and Yersinia pseudotuberculosis. Infect. Immun. 2003, 71, 5892–5899. [Google Scholar] [CrossRef]
- Sarantis, H.; Grinstein, S. Subversion of phagocytosis for pathogen survival. Cell Host Microbe 2012, 12, 419–431. [Google Scholar] [CrossRef]
- Belon, C.; Blanc-Potard, A.B. Intramacrophage Survival for Extracellular Bacterial Pathogens: MgtC As a Key Adaptive Factor. Front. Cell Infect. Microbiol. 2016, 6, 52. [Google Scholar] [CrossRef]
- Alix, E.; Blanc-Potard, A.B. MgtC: A key player in intramacrophage survival. Trends Microbiol. 2007, 15, 252–256. [Google Scholar] [CrossRef]
- Buchmeier, N.; Blanc-Potard, A.; Ehrt, S.; Piddington, D.; Riley, L.; Groisman, E.A. A parallel intraphagosomal survival strategy shared by Mycobacterium tuberculosis and Salmonella enterica. Mol. Microbiol. 2000, 35, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Grabenstein, J.P.; Fukuto, H.S.; Palmer, L.E.; Bliska, J.B. Characterization of phagosome trafficking and identification of PhoP-regulated genes important for survival of Yersinia pestis in macrophages. Infect. Immun. 2006, 74, 3727–3741. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, J.P.; O’Callaghan, D.; Blanc-Potard, A.B. Requirement of MgtC for Brucella suis intramacrophage growth: A potential mechanism shared by Salmonella enterica and Mycobacterium tuberculosis for adaptation to a low-Mg2+ environment. Infect. Immun. 2005, 73, 3160–3163. [Google Scholar] [CrossRef] [PubMed]
- Maloney, K.E.; Valvano, M.A. The mgtC gene of Burkholderia cenocepacia is required for growth under magnesium limitation conditions and intracellular survival in macrophages. Infect. Immun. 2006, 74, 5477–5486. [Google Scholar] [CrossRef]
- Bernut, A.; Belon, C.; Soscia, C.; Bleves, S.; Blanc-Potard, A.B. Intracellular phase for an extracellular bacterial pathogen: MgtC shows the way. Microb. Cell. 2015, 2, 353–355. [Google Scholar] [CrossRef]
- Lee, E.J.; Pontes, M.H.; Groisman, E.A. A bacterial virulence protein promotes pathogenicity by inhibiting the bacterium’s own F1Fo ATP synthase. Cell 2013, 154, 146–156. [Google Scholar] [CrossRef]
- Pontes, M.H.; Lee, E.J.; Choi, J.; Groisman, E.A. Salmonella promotes virulence by repressing cellulose production. Proc. Natl. Acad. Sci. USA 2015, 112, 5183–5188. [Google Scholar] [CrossRef]
- Choi, S.; Choi, E.; Cho, Y.J.; Nam, D.; Lee, J.; Lee, E.J. The Salmonella virulence protein MgtC promotes phosphate uptake inside macrophages. Nat. Commun. 2019, 10, 3326. [Google Scholar] [CrossRef]
- Moncrief, M.B.; Maguire, M.E. Magnesium and the role of MgtC in growth of Salmonella typhimurium. Infect. Immun. 1998, 66, 3802–3809. [Google Scholar] [CrossRef]
- Garcia Vescovi, E.; Soncini, F.C.; Groisman, E.A. Mg2+ as an extracellular signal: Environmental regulation of Salmonella virulence. Cell 1996, 84, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Tao, T.; Snavely, M.D.; Farr, S.G.; Maguire, M.E. Magnesium transport in Salmonella typhimurium: mgtA encodes a P-type ATPase and is regulated by Mg2+ in a manner similar to that of the mgtB P-type ATPase. J. Bacteriol. 1995, 177, 2654–2662. [Google Scholar] [CrossRef] [PubMed]
- Blanc-Potard, A.B.; Groisman, E.A. The Salmonella selC locus contains a pathogenicity island mediating intramacrophage survival. EMBO J. 1997, 16, 5376–5385. [Google Scholar] [CrossRef]
- Sun, W.; Sanapala, S.; Rahav, H.; Curtiss, R., 3rd. Oral administration of a recombinant attenuated Yersinia pseudotuberculosis strain elicits protective immunity against plague. Vaccine 2015, 33, 6727–6735. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Sanapala, S.; Henderson, J.C.; Sam, S.; Olinzock, J.; Trent, M.S.; Curtiss, R., 3rd. LcrV delivered via type III secretion system of live attenuated Yersinia pseudotuberculosis enhances immunogenicity against pneumonic plague. Infect. Immun. 2014, 82, 4390–4404. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Six, D.A.; Reynolds, C.M.; Chung, H.S.; Raetz, C.R.; Curtiss, R., 3rd. Pathogenicity of Yersinia pestis synthesis of 1-dephosphorylated lipid A. Infect. Immun. 2013, 81, 1172–1185. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Tian, M.; Hu, H.; Yin, Y.; Guan, X.; Ding, C.; Wang, S.; Yu, S. Lable-free based comparative proteomic analysis of secretory proteins of rough Brucella mutants. J. Proteom. 2019, 195, 66–75. [Google Scholar] [CrossRef]
- Wang, S.; Li, Y.; Scarpellini, G.; Kong, W.; Shi, H.; Baek, C.H.; Gunn, B.; Wanda, S.Y.; Roland, K.L.; Zhang, X.; et al. Salmonella vaccine vectors displaying delayed antigen synthesis in vivo to enhance immunogenicity. Infect. Immun. 2010, 78, 3969–3980. [Google Scholar] [CrossRef]
- McClure, R.; Balasubramanian, D.; Sun, Y.; Bobrovskyy, M.; Sumby, P.; Genco, C.A.; Vanderpool, C.K.; Tjaden, B. Computational analysis of bacterial RNA-Seq data. Nucleic Acids Res. 2013, 41, e140. [Google Scholar] [CrossRef]
- Diler, E.; Saul, S.; Bogeski, I.; Meier, C.; Tschernig, T. Influence of extracellular magnesium on phagocytosis and free cytosolic Mg levels in differentiated U937 and MH-S cells. Magnes. Res. 2015, 28, 23–31. [Google Scholar] [CrossRef]
- Cornelis, G.R.; Van Gijsegem, F. Assembly and function of type III secretory systems. Annu. Rev. Microbiol. 2000, 54, 735–774. [Google Scholar] [CrossRef] [PubMed]
- Brubaker, R.R. Interleukin-10 and inhibition of innate immunity to Yersiniae: Roles of Yops and LcrV (V antigen). Infect. Immun. 2003, 71, 3673–3681. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, G.R. The Yersinia Ysc-Yop ‘type III’ weaponry. Nat. Rev. Mol. Cell Biol. 2002, 3, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.W. Salmonella acid shock proteins are required for the adaptive acid tolerance response. J. Bacteriol. 1991, 173, 6896–6902. [Google Scholar] [CrossRef]
- Lestrate, P.; Delrue, R.M.; Danese, I.; Didembourg, C.; Taminiau, B.; Mertens, P.; De Bolle, X.; Tibor, A.; Tang, C.M.; Letesson, J.J. Identification and characterization of in vivo attenuated mutants of Brucella melitensis. Mol. Microbiol. 2000, 38, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Smith, I. Mycobacterium tuberculosis pathogenesis and molecular determinants of virulence. Clin. Microbiol. Rev. 2003, 16, 463–496. [Google Scholar] [CrossRef] [PubMed]
- Snavely, M.D.; Miller, C.G.; Maguire, M.E. The mgtB Mg2+ transport locus of Salmonella typhimurium encodes a P-type ATPase. J. Biol. Chem. 1991, 266, 815–823. [Google Scholar] [CrossRef]
- Blanc-Potard, A.B.; Lafay, B. MgtC as a horizontally-acquired virulence factor of intracellular bacterial pathogens: Evidence from molecular phylogeny and comparative genomics. J. Mol. Evol. 2003, 57, 479–486. [Google Scholar] [CrossRef]
- Groisman, E.A.; Hollands, K.; Kriner, M.A.; Lee, E.J.; Park, S.Y.; Pontes, M.H. Bacterial Mg2+ homeostasis, transport, and virulence. Annu. Rev. Genet. 2013, 47, 625–646. [Google Scholar] [CrossRef]
- Cornelis, G.R.; Boland, A.; Boyd, A.P.; Geuijen, C.; Iriarte, M.; Neyt, C.; Sory, M.P.; Stainier, I. The virulence plasmid of Yersinia, an antihost genome. Microbiol. Mol. Biol. Rev. 1998, 62, 1315–1352. [Google Scholar] [CrossRef]
- Tsukano, H.; Kura, F.; Inoue, S.; Sato, S.; Izumiya, H.; Yasuda, T.; Watanabe, H. Yersinia pseudotuberculosis blocks the phagosomal acidification of B10.A mouse macrophages through the inhibition of vacuolar H(+)-ATPase activity. Microb. Pathog. 1999, 27, 253–263. [Google Scholar] [CrossRef]
- Zhang, Y.; Murtha, J.; Roberts, M.A.; Siegel, R.M.; Bliska, J.B. Type III secretion decreases bacterial and host survival following phagocytosis of Yersinia pseudotuberculosis by macrophages. Infect. Immun. 2008, 76, 4299–4310. [Google Scholar] [CrossRef]
- Bartra, S.; Cherepanov, P.; Forsberg, A.; Schesser, K. The Yersinia YopE and YopH type III effector proteins enhance bacterial proliferation following contact with eukaryotic cells. BMC Microbiol. 2001, 1, 22. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.W. The acid tolerance response of Salmonella typhimurium involves transient synthesis of key acid shock proteins. J. Bacteriol. 1993, 175, 1981–1987. [Google Scholar] [CrossRef] [PubMed]
- Bearson, B.L.; Wilson, L.; Foster, J.W. A low pH-inducible, PhoPQ-dependent acid tolerance response protects Salmonella typhimurium against inorganic acid stress. J. Bacteriol. 1998, 180, 2409–2417. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Characteristics | Source of Reference |
|---|---|---|
| Strains | ||
| E. coli strain | ||
| DH5α | F– φ80lacZΔM15 Δ(lacZYA-argF)U169 recA1 endA1 hsdR17(rK−, K+) phoA supE44 λ− thi-1 gyrA96 relA1 | Invitrogen |
| χ7213 | thi-1 thr-1 leuB6 fhuA21 lacY1 glnV44 ΔasdA4 recA1 RP4 2-Tc::Mu [λpir]; Kmr | [24] |
| Y. pseudotuberculosis strain | ||
| PB1+ | Wild-type, serotype O:1b | Lab collection |
| PB1+ pYV− | Lack of virulence plasmid pYV | This study |
| YPtbS33 | ΔmgtC in PB1+ strain | This study |
| YPtbS34 | ΔmgtC in PB1+ pYV− strain | This study |
| YPtbS35 | ΔYPTS_2313 in PB1+ strain | This study |
| YPtbS36 | ΔYPTS_3071 in PB1+ strain | This study |
| YPtbS37 | ΔYPTS_2313ΔYPTS_3071 in PB1+ strain | This study |
| YPtbS33(C-mgtC) | Ampr, ΔmgtC strain carrying the complementary plasmid pYA4454-mgtC | This study |
| YPtbS34 (C-mgtC) | Ampr, ΔmgtC pYV− strain carrying the complementary plasmid pYA4454-mgtC | This study |
| YPtbS35 (C-YPTS_2313) | Ampr, ΔYPTS_2313 strain carrying the complementary plasmid pYA4454-YPTS_2313 | This study |
| YPtbS36 (C-YPTS_2307) | Ampr, ΔYPTS_3071 strain carrying the complementary plasmid pYA4454-YPTS_3071 | This study |
| Plasmids | ||
| pRE112 | Suicide vector, Cmr, mob− (RP4)R6K ori, sacB | [25] |
| pYA4454 | Complemental plasmid, Ampr | [26] |
| pSMV55 | Cmr, pRE112 plasmid containing the ΔmgtC fragment | This study |
| pSMV56 | Cmr, pRE112 plasmid containing the ΔYPTS_2313 fragment | This study |
| pSMV57 | Cmr, pRE112 plasmid containing the ΔYPTS_3071 fragment | This study |
| pSMV58 | Ampr, pYA4454 plasmid containing the mgtC gene | This study |
| pSMV59 | Ampr, pYA4454 plasmid containing the YPTS_2313 gene | This study |
| pSMV60 | Ampr, pYA4454 plasmid containing the YPTS_3071gene | This study |
| Gene Number | Gene Name | Fold Change ΔmgtC/PB1+ | p-Value |
|---|---|---|---|
| YPTS_0907 | Transporter substrate-binding domain-containing protein | 6.494781 | 1.04 × 10−8 |
| YPTS_1642 | Sugar ABC transporter substrate-binding protein | 6.125791 | 1.59 × 10−6 |
| YPTS_3843 | Malate synthase A | 5.255804 | 1.16 × 10−10 |
| YPTS_2011 | Aspartate aminotransferase family protein | 5.083721 | 1.85 × 10−7 |
| YPTS_3842 | Isocitrate lyase | 5.081315 | 4.39 × 10−9 |
| YPTS_2313 | Acid shock protein | −3.712933 | 8.92 × 10−24 |
| YPTS_0225 | Glycerol-3-phosphate dehydrogenase subunit GlpB | −3.838456 | 7.86 × 10−35 |
| YPTS_0224 | Anaerobic glycerol-3-phosphate dehydrogenase subunit A | −4.235415 | 9.02 × 10−37 |
| YPTS_0091 | Aquaporin family protein | −4.564323 | 1.58 × 10−31 |
| YPTS_3071 | Acid shock protein | −5.886112 | 1.59 × 10−9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Wang, H.; Sun, W.; Ding, J. Impact of MgtC on the Fitness of Yersinia pseudotuberculosis. Pathogens 2023, 12, 1428. https://doi.org/10.3390/pathogens12121428
Li P, Wang H, Sun W, Ding J. Impact of MgtC on the Fitness of Yersinia pseudotuberculosis. Pathogens. 2023; 12(12):1428. https://doi.org/10.3390/pathogens12121428
Chicago/Turabian StyleLi, Peng, Hengtai Wang, Wei Sun, and Jiabo Ding. 2023. "Impact of MgtC on the Fitness of Yersinia pseudotuberculosis" Pathogens 12, no. 12: 1428. https://doi.org/10.3390/pathogens12121428