
Higher Resistance of Yersinia enterocolitica in Comparison to Yersinia pseudotuberculosis to Antibiotics and Cinnamon, Oregano and Thyme Essential Oils

Abstract
:1. Introduction
2. Materials and Methods
2.1. Yersinia Strains
2.2. Essential Oils
2.3. Determination of Minimum Inhibitory Concentration of Antibiotics
2.4. Determination of Minimum Inhibitory Concentration of Essential Oils
2.5. Statistical Analysis
3. Results and Discussion
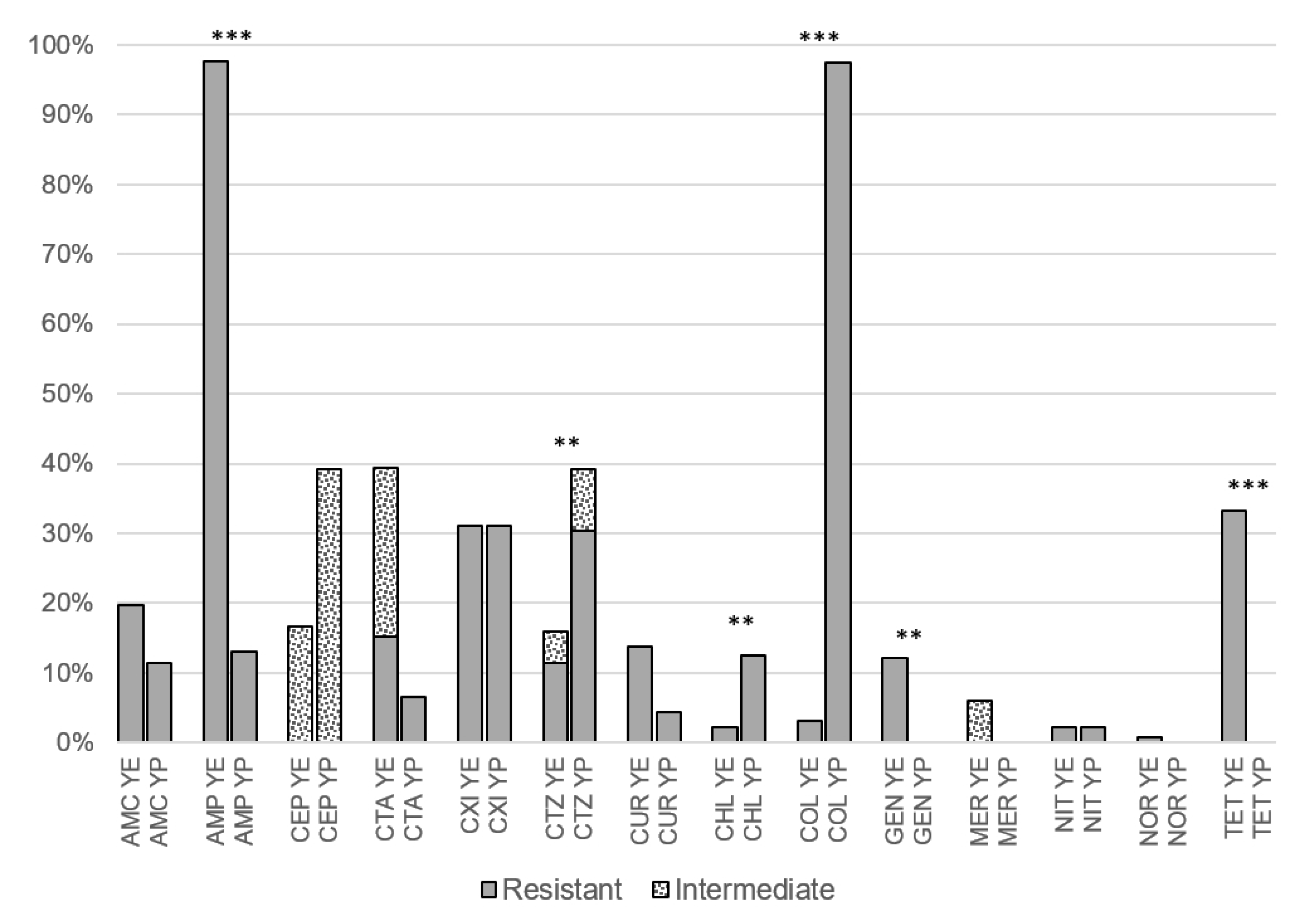
3.1. Antibiotic Resistance
3.2. Essential Oils
4. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA and ECDC (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, 6971. [Google Scholar] [CrossRef]
- Cabanel, N.; Galimand, M.; Bouchier, C.; Chesnokova, M.; Klimov, V.; Carniel, E. Molecular bases for multidrug resistance in Yersinia pseudotuberculosis. Int. J. Med. Microbiol. 2017, 307, 371–381. [Google Scholar] [CrossRef]
- Bancerz-Kisiel, A.; Szweda, W. Yersiniosis—A zoonotic foodborne disease of relevance to public health. Ann. Agric. Environ. Med. 2015, 22, 397–402. [Google Scholar] [CrossRef]
- Fàbrega, A.; Ballesté-Delpierre, C.; Vila, J. Antimicrobial Resistance in Yersinia enterocolitica. In Antimicrobial Resistance and Food Safety, 1st ed.; Chen, C.-Y., Yan, X., Jackson, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; Chapter 5; pp. 77–104. [Google Scholar] [CrossRef]
- Koskinen, J.; Ortiz-Martinez, P.; Keto-Timonen, R.; Joutsen, S.; Fredriksson-Ahomaa, M.; Korkeala, H. Prudent antimicrobial use is essential to prevent the emergence of antimicrobial resistance in Yersinia enterocolitica 4/O:3 strains in pigs. Front. Microbiol. 2022, 13, 841. [Google Scholar] [CrossRef]
- EUCAST (The European Committee on Antimicrobial Susceptibility Testing). Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 11 February 2022).
- Tavassoli, M.; Afshari, A.; Drăgănescu, D.; Arsene, A.L.; Burykina, T.I.; Rezaee, R. Antimicrobial resistance of Yersinia enterocolitica in different foods. A review. Farmacia 2018, 66, 399–407. [Google Scholar]
- Mittal, R.P.; Rana, A.; Jaitak, V. Essential oils: An impending substitute of synthetic antimicrobial agents to overcome antimicrobial resistance. Curr. Drug Targets 2019, 20, 605–624. [Google Scholar] [CrossRef]
- Bonardi, S.; Bruini, I.; D’Incau, M.; Van Damme, I.; Carniel, E.; Brémont, S.; Cavallini, P.; Tagliabue, S.; Brindani, F. Detection, seroprevalence and antimicrobial resistance of Yersinia enterocolitica and Yersinia pseudotuberculosis in pig tonsils in Northern Italy. Int. J. Food Microbiol. 2016, 235, 125–132. [Google Scholar] [CrossRef]
- Verbikova, V.; Borilova, G.; Babak, V.; Moravkova, M. Prevalence, characterization and antimicrobial susceptibility of Yersinia enterocolitica and other Yersinia species found in fruits and vegetables from the European Union. Food Control 2018, 85, 161–167. [Google Scholar] [CrossRef]
- EUCAST (The European Committee on Antimicrobial Susceptibility Testing). Broth Microdilution—EUCAST Reading Guide. Version 3.0. 2021. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Disk_test_documents/2022_manuals/Reading_guide_BMD_v_4.0_2022.pdf (accessed on 24 January 2022).
- Hulankova, R. The influence of liquid medium choice in determination of minimum inhibitory concentration of essential oils against pathogenic bacteria. Antibiotics 2022, 11, 150. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. 2021. Available online: https://www.R-project.org/ (accessed on 27 April 2021).
- Christensen, R.H.B. Ordinal—Regression Models for Ordinal Data. R Package Version 2019.12-10. 2019. Available online: https://CRAN.R-project.org/package=ordinal (accessed on 10 August 2022).
- Russel, L. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.5.1. 2020. Available online: https://CRAN.R-project.org/package=emmeans, (accessed on 7 May 2021).
- Modesto, P.; De Ciucis, C.G.; Vencia, W.; Pugliano, M.C.; Mignone, W.; Berio, E.; Masotti, C.; Ercolini, C.; Serracca, L.; Andreoli, T.; et al. Evidence of antimicrobial resistance and presence of pathogenicity genes in Yersinia enterocolitica isolate from wild boars. Pathogens 2021, 10, 398. [Google Scholar] [CrossRef]
- Ossiprandi, M.C.; Zerbini, L. Prevalence and antibiotic susceptibilities of pathogenic Yersinia enterocolitica strains in pigs slaughtered in northern Italy. J. Adv. Biol. 2014, 5, 603–609. [Google Scholar] [CrossRef]
- Reinhardt, M.; Hammerl, J.A.; Kunz, K.; Barac, A.; Nöckler, K.; Hertwig, S. Yersinia pseudotuberculosis prevalence and diversity in wild boars in Northeast Germany. Appl. Environ. Microbiol. 2018, 84, e00675. [Google Scholar] [CrossRef] [Green Version]
- Kechagia, N.; Nicolaou, C.; Ioannidou, V.; Kourti, E.; Ioannidis, A.; Legakis, N.J.; Chatzipanagiotou, S. Detection of chromosomal and plasmid-encoded virulence determinants in Yersinia enterocolitica and other Yersinia spp. isolated from food animals in Greece. Int. J. Food Microbiol. 2007, 118, 326–331. [Google Scholar] [CrossRef]
- Terentjeva, M.; Bẽrziņs, A. Prevalence and antimicrobial resistance of Yersinia enterocolitica and Yersinia pseudotuberculosis in slaughter pigs in Latvia. J. Food Prot. 2010, 73, 1335–1338. [Google Scholar] [CrossRef]
- Blomme, S.; Andre, E.; Delmée, M.; Verhaegen, J. Antimicrobial susceptibility testing of Yersinia enterocolitica and Yersinia pseudotuberculosis. In Proceedings of the 27th European Congress of Clinical Microbiology and Infectious Diseases, Vienna, Austria, 22–25 April 2017; p. 0521. [Google Scholar]
- Simonova, J.; Borilova, G.; Steinhauserova, I. Occurence of pathogenic strains of Yersinia enterocolitica in pigs and their antimicrobial resistance. Bull. Vet. Inst. Pulawy 2008, 52, 39–43. [Google Scholar]
- Bengoechea, J.A.; Lindner, B.; Seydel, U.; Díaz, R.; Moriyón, I. Yersinia pseudotuberculosis and Yersinia pestis are more resistant to bactericidal cationic peptides than Yersinia enterocolitica. Microbiology 1998, 144, 1509–1515. [Google Scholar] [CrossRef] [Green Version]
- Durofil, A.; Maddela, N.R.; Naranjo, R.A.; Radice, M. Evidence on antimicrobial activity of essential oils and herbal extracts against Yersinia enterocolitica—A review. Food Biosci. 2022, 47, 101712. [Google Scholar] [CrossRef]
- Ebani, V.V.; Nardoni, S.; Bertelloni, F.; Giovanelli, S.; Rocchigiani, G.; Pistelli, L.; Mancianti, F. Antibacterial and antifungal activity of essential oils against some pathogenic bacteria and yeasts shed from poultry. Flavour Fragr. J. 2016, 31, 302–309. [Google Scholar] [CrossRef]
- Görmez, A.; Yanmiş, D.; Bozari, S.; Gürkök, S. Antibacterial activity of essential oils extracted from Satureja hortensis against selected clinical pathogens. AIP Conf. Proc. 2017, 1833, 020059. [Google Scholar] [CrossRef]
- Owen, L.; Laird, K. Synchronous application of antibiotics and essential oils: Dual mechanisms of action as a potential solution to antibiotic resistance. Crit. Rev. Microbiol. 2018, 44, 414–435. [Google Scholar] [CrossRef]
- Pesingi, P.V.; Singh, B.R.; Pesingi, P.K.; Bhardwaj, M.; Singh, S.V.; Kumawat, M.; Sinha, D.K.; Gandham, K.R. MexAB-OprM efflux pump of Pseudomonas aeruginosa offers resistance to carvacrol: A herbal antimicrobial agent. Front. Microbiol. 2019, 10, 2664. [Google Scholar] [CrossRef]
- Hulankova, R.; Borilova, G. Modeling dependence of growth inhibition of Salmonella Typhimurium and Listeria monocytogenes by oregano or thyme essential oils on the chemical composition of minced pork. J Food Saf. 2020, 40, e12818. [Google Scholar] [CrossRef]
- Boskovic, M.; Djordjevic, J.; Ivanovic, J.; Janjic, J.; Zdravkovic, N.; Glisic, M.; Glamoclija, N.; Baltic, B.; Djordjevic, V.; Baltic, M. Inhibition of Salmonella by thyme essential oil and its effect on microbiological and sensory properties of minced pork meat packaged under vacuum and modified atmosphere. Int. J. Food Microbiol. 2017, 258, 58–67. [Google Scholar] [CrossRef]


{kind=link}
| Species | Bioserotype | Origin | Number of Resistances | Number of Strains |
|---|---|---|---|---|
| Y. enterocolitica | 1A/O:5, O:8 | Vegetables | 6 | 1 |
| 5 | 1 | |||
| 4 | 8 | |||
| 3 | 2 | |||
| Y. enterocolitica | 4/O:3 | Domestic swine | 8 | 1 |
| 6 | 5 | |||
| 5 | 6 | |||
| Y. pseudotuberculosis | - | Wild boar | 5 | 2 |
| 4 | 5 | |||
| 3 | 5 |
| Antibiotic | Y. enterocolitica | Y. pseudotuberculosis | p Level | ||||
|---|---|---|---|---|---|---|---|
| MIC90 * | MIC50 † | Min–Max § | MIC90 | MIC50 | Min–Max | ||
| Amoxiclav | 16 | 2 | 2–32 | 16 | 2 | 2–32 | 0.011 |
| Ampicillin | 32 | 32 | 4–32 | 12 | 2 | 2–16 | <0.001 |
| Cefepime | 4 | 1 | 1–4 | 2 | 1 | 1–4 | 0.017 |
| Cefotaxime | 4 | 1 | 1–8 | 1 | 1 | 1–4 | <0.001 |
| Cefoxitin | 32 | 8 | 4–66 | 32 | 4 | 4–64 | 0.134 |
| Ceftazidime | 8 | 1 | 1–64 | 32 | 1 | 1–64 | 0.002 |
| Cefuroxime | 16 | 8 | 2–64 | 8 | 2 | 1–32 | <0.001 |
| Chloramphenicol | 8 | 4 | 1–16 | 16 | 4 | 4–16 | 0.575 |
| Ciprofloxacin | 0.25 | 0.25 | 0.025–0.5 | 0.25 | 0.25 | 0.25–0.25 | 0.552 |
| Colistin | 0.5 | 0.5 | 0.5–16 | 16 | 16 | 0.5–16 | <0.001 |
| Erythromycin | 64 | 32 | 4–128 | 64 | 64 | 32–64 | <0.001 |
| Gentamicin | 4 | 1 | 1–4 | 1 | 1 | 1–1 | <0.001 |
| Imipenem | 0.475 | 0.25 | 0.25–0.5 | 0.25 | 0.25 | 0.25–0.25 | 0.039 |
| Meropenem | 1 | 0.25 | 0.25–4 | 0.25 | 0.25 | 0.25–0.25 | <0.001 |
| Nitrofurantoin | 64 | 32 | 16–128 | 64 | 64 | 16–128 | 0.001 |
| Norfloxacin | 0.5 | 0.5 | 0.5–16 | 0.5 | 0.5 | 0.5–0.5 | 0.564 |
| Streptomycin | 16 | 8 | 1–32 | 64 | 8 | 2–64 | 0.242 |
| Tetracycline | 8 | 4 | 0.5–16 | 2.2 | 2 | 1–4 | <0.001 |
| Tobramycin | 2 | 1 | 1–2 | 1 | 1 | 1–1 | 0.008 |
| Trimethoprim | 2 | 2 | 0.5–4 | 1 | 1 | 0.5–2 | <0.001 |
| Trimethoprim–Sulfamethoxazole | 20 | 20 | 20–20 | 20 | 20 | 20–20 | 1.000 |
| Phenotype | Y. enterocolitica | Phenotype | Y. pseudotuberculosis | ||
|---|---|---|---|---|---|
| R * | N † | R | N | ||
| Amp | 1 | 30 (22.7%) | Col | 1 | 14 (10.6%) |
| AmpTet | 2 | 22 (16.7%) | ChlCol | 2 | 6 (4.5%) |
| AmpCxi | 2 | 14 (10.6%) | CxiCol | 2 | 4 (3.0%) |
| AmcAmpCxi | 3 | 9 (6.8%) | AmcCol | 2 | 3 (2.3%) |
| AmpGen | 2 | 7 (5.3%) | AmcAmpCxiCol | 4 | 2 (1.5%) |
| AmpGenTet | 3 | 6 (4.5%) | AmpCxiCtzCol | 4 | 2 (1.5%) |
| AmcAmp | 2 | 4 (3.0%) | CtaCtz | 2 | 2 (1.5%) |
| AmpCtaCtzCurGenTet | 6 | 3 (2.3%) | AmpCxiCtzCurCol | 5 | 1 (0.8%) |
| AmpCtaCtzCurTet | 5 | 3 (2.3%) | CtaCxiCtzCurCol | 5 | 1 (0.8%) |
| AmcAmpCtaCxi | 4 | 3 (2.3%) | CxiCtzChlCol | 4 | 1 (0.8%) |
| AmpCxiTet | 3 | 3 (2.3%) | AmpCtzCol | 3 | 1 (0.8%) |
| AmpCur | 2 | 3 (2.3%) | CxiCtzCol | 3 | 1 (0.8%) |
| AmcAmpCxiCur | 4 | 2 (1.5%) | CxiCtzNit | 3 | 1 (0.8%) |
| AmcAmpCxiNit | 4 | 2 (1.5%) | CxiChlCol | 3 | 1 (0.8%) |
| AmpCurTet | 3 | 2 (1.5%) | CtzChlCol | 3 | 1 (0.8%) |
| AmpCta | 2 | 2 (1.5%) | CxiCtz | 2 | 1 (0.8%) |
| AmcAmpCtaCxiCtzCurChlNit | 8 | 1 (0.8%) | CtzCol | 2 | 1 (0.8%) |
| AmcAmpCtaCxiCtzCol | 6 | 1 (0.8%) | Ctz | 1 | 1 (0.8%) |
| AmpCtaCxiCtzCurTet | 6 | 1 (0.8%) | Chl | 1 | 1 (0.8%) |
| AmcAmpCtaCxiChl | 5 | 1 (0.8%) | sensitive | 0 | 1 (0.8%) |
| AmpCtaCxiCtzCur | 5 | 1 (0.8%) | |||
| AmpCtaCxiCtzCol | 5 | 1 (0.8%) | |||
| AmpCtaCtzTet | 4 | 1 (0.8%) | |||
| AmcAmpCur | 3 | 1 (0.8%) | |||
| AmcAmpChl | 3 | 1 (0.8%) | |||
| AmpCtaCtz | 3 | 1 (0.8%) | |||
| AmpCtaTet | 3 | 1 (0.8%) | |||
| AmpCxiCol | 3 | 1 (0.8%) | |||
| AmcCxiTet | 3 | 1 (0.8%) | |||
| AmpCtzCur | 3 | 1 (0.8%) | |||
| AmpNorTet | 3 | 1 (0.8%) | |||
| CtzCol | 2 | 1 (0.8%) | |||
| sensitive | 0 | 1 (0.8%) | |||
| YE 1A (n = 12) | YE 4/O:3 (n = 12) | YP (n = 12) | |
|---|---|---|---|
| median [range] | median [range] | median [range] | |
| Origin | vegetable | domestic swine | wild boar |
| Cinnamon | 414 Aa [207; 517] | 310 Aa [207; 414] | 207 Ab [103; 414] |
| Oregano | 379 Aa [284; 474] | 379 Aa [284; 474] | 284 Ab [190; 474] |
| Thyme | 738 Ba [553; 922] | 738 Ba [553; 922] | 553 Bb [369; 738] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hulankova, R. Higher Resistance of Yersinia enterocolitica in Comparison to Yersinia pseudotuberculosis to Antibiotics and Cinnamon, Oregano and Thyme Essential Oils. Pathogens 2022, 11, 1456. https://doi.org/10.3390/pathogens11121456
Hulankova R. Higher Resistance of Yersinia enterocolitica in Comparison to Yersinia pseudotuberculosis to Antibiotics and Cinnamon, Oregano and Thyme Essential Oils. Pathogens. 2022; 11(12):1456. https://doi.org/10.3390/pathogens11121456
Chicago/Turabian StyleHulankova, Radka. 2022. "Higher Resistance of Yersinia enterocolitica in Comparison to Yersinia pseudotuberculosis to Antibiotics and Cinnamon, Oregano and Thyme Essential Oils" Pathogens 11, no. 12: 1456. https://doi.org/10.3390/pathogens11121456