



Identifying Knowledge Gaps through the Systematic Review of Temperature-Driven Variability in the Competence of Aedes aegypti and Ae. albopictus for Chikungunya Virus


Abstract
:1. Introduction
2. Materials and Methods
3. Results
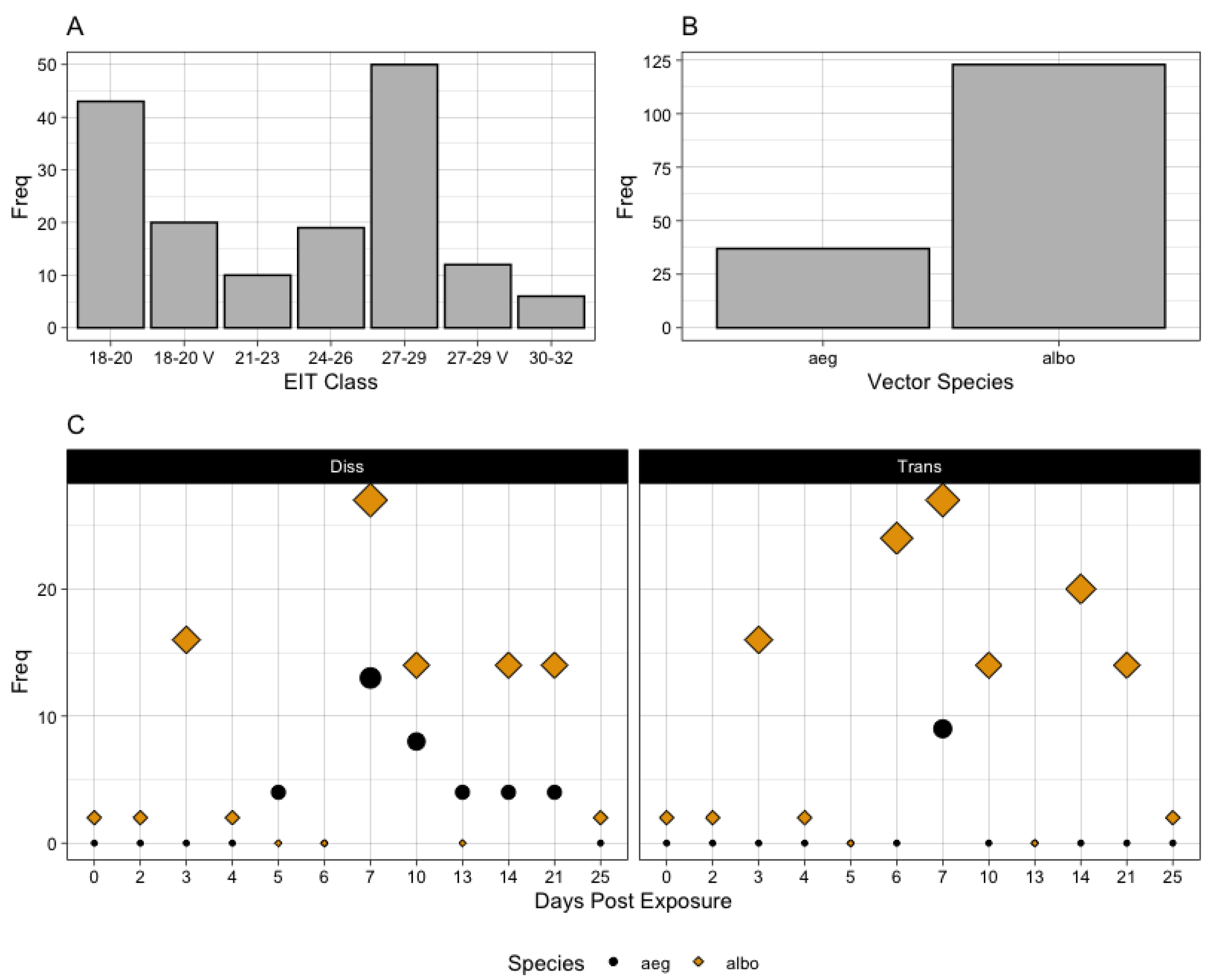
3.1. State of the Literature
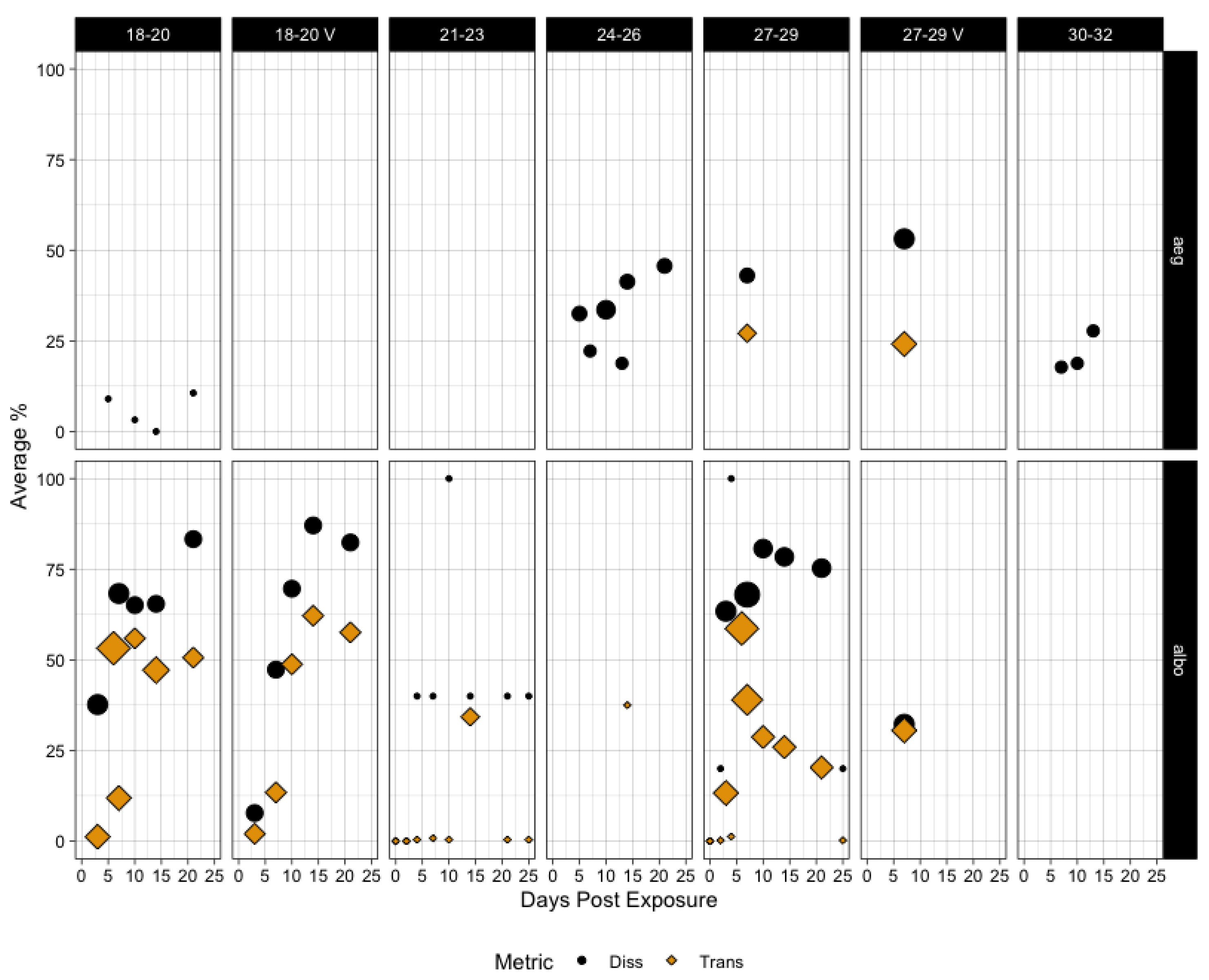
3.2. State of Knowledge Regarding Temperature Dependence and In Vivo Dynamics in Aedes aegypti and albopictus
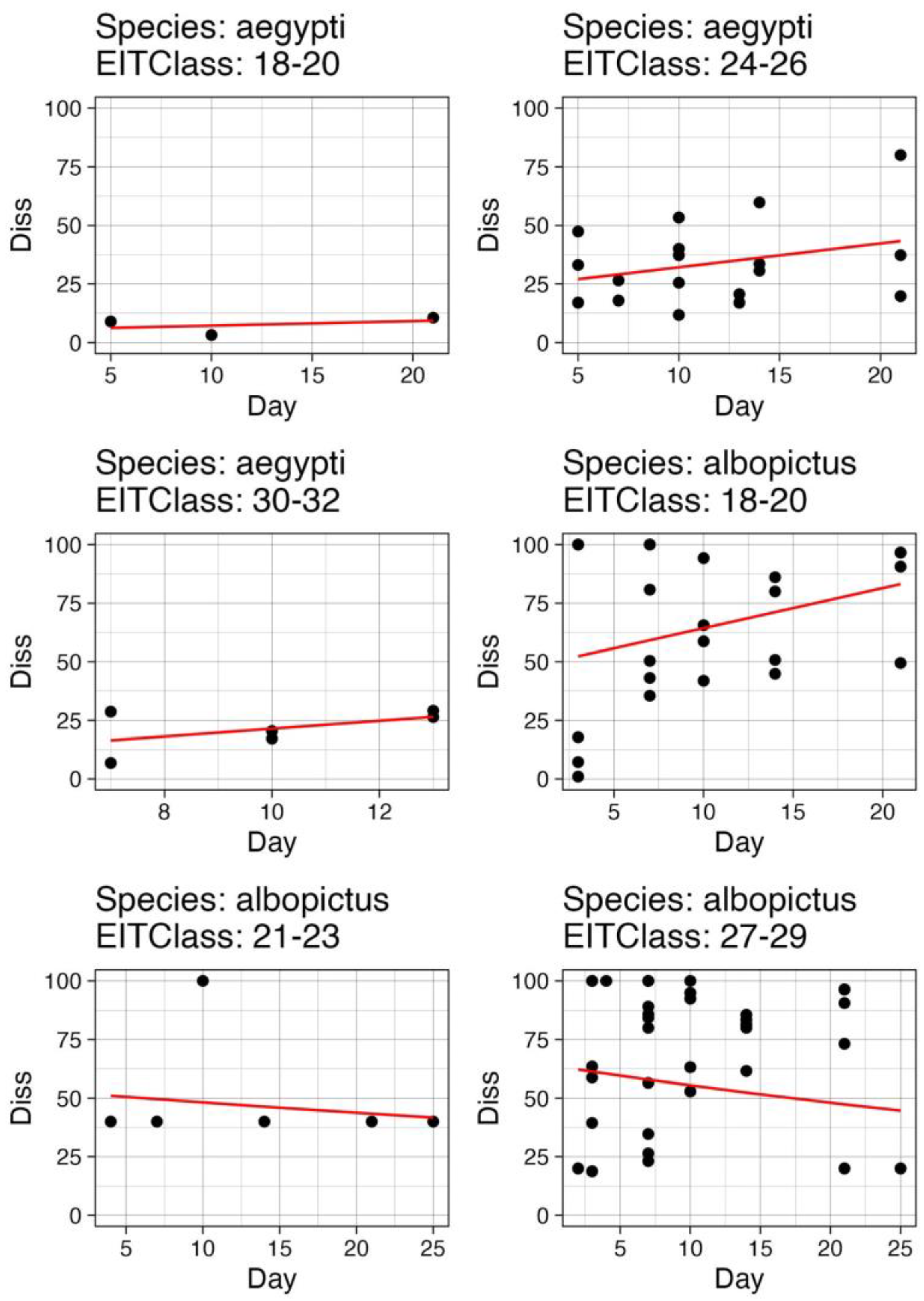
3.3. Functional Fits to Available Data
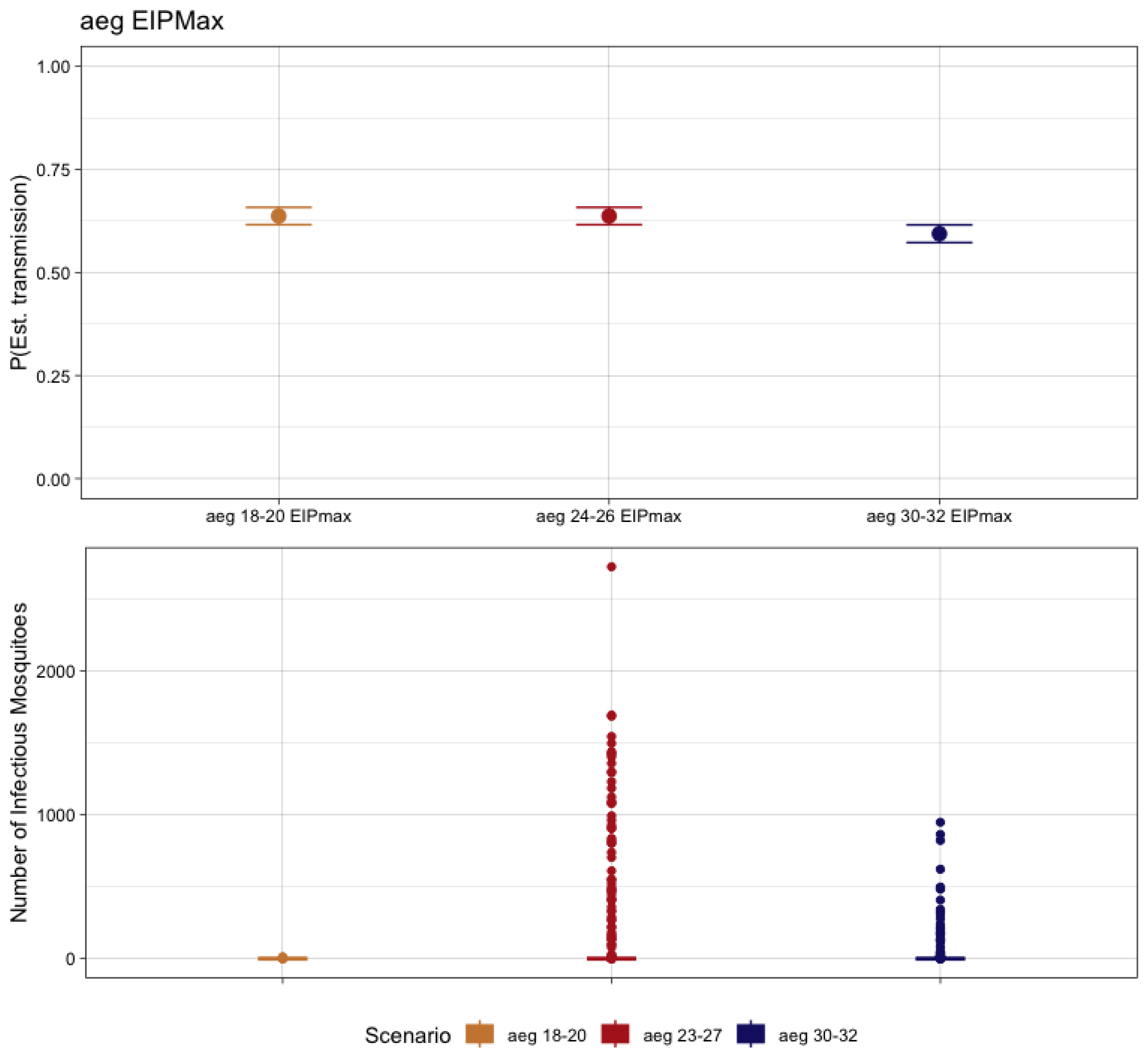
3.4. Ae. aegypti-CHIKV Transmission and Temperature
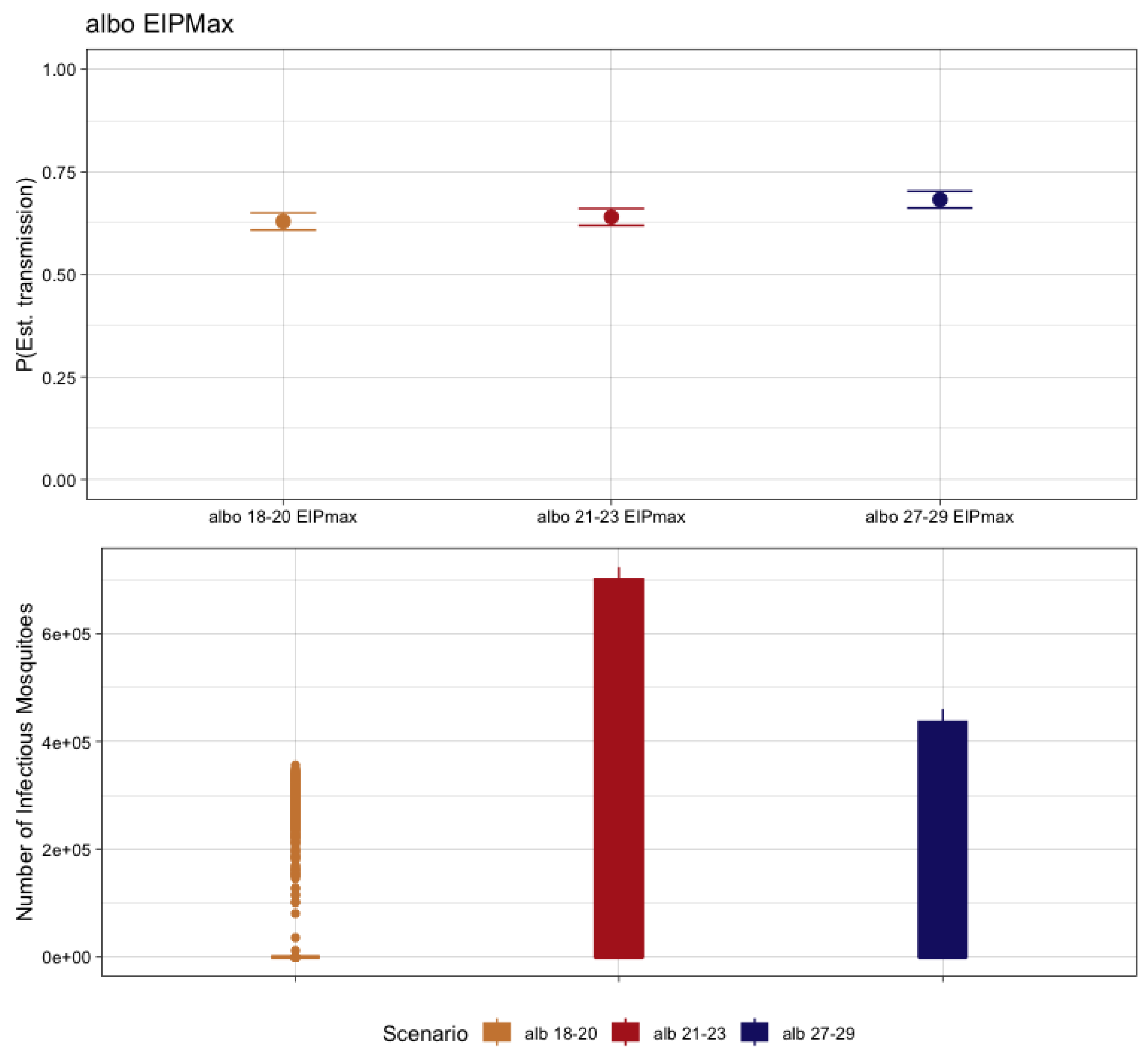
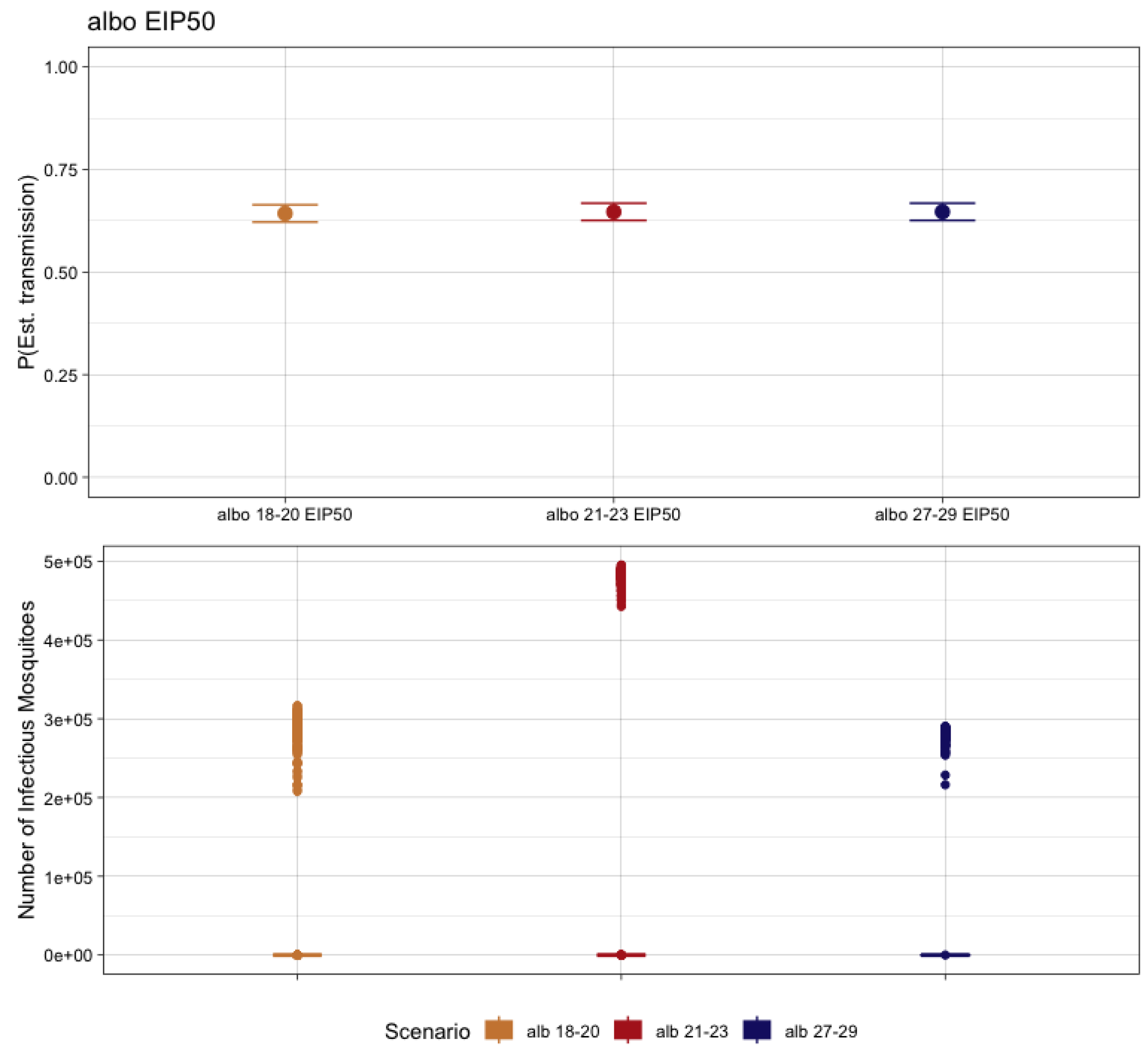
3.5. Ae. albopictus-CHIKV Transmission and Temperature
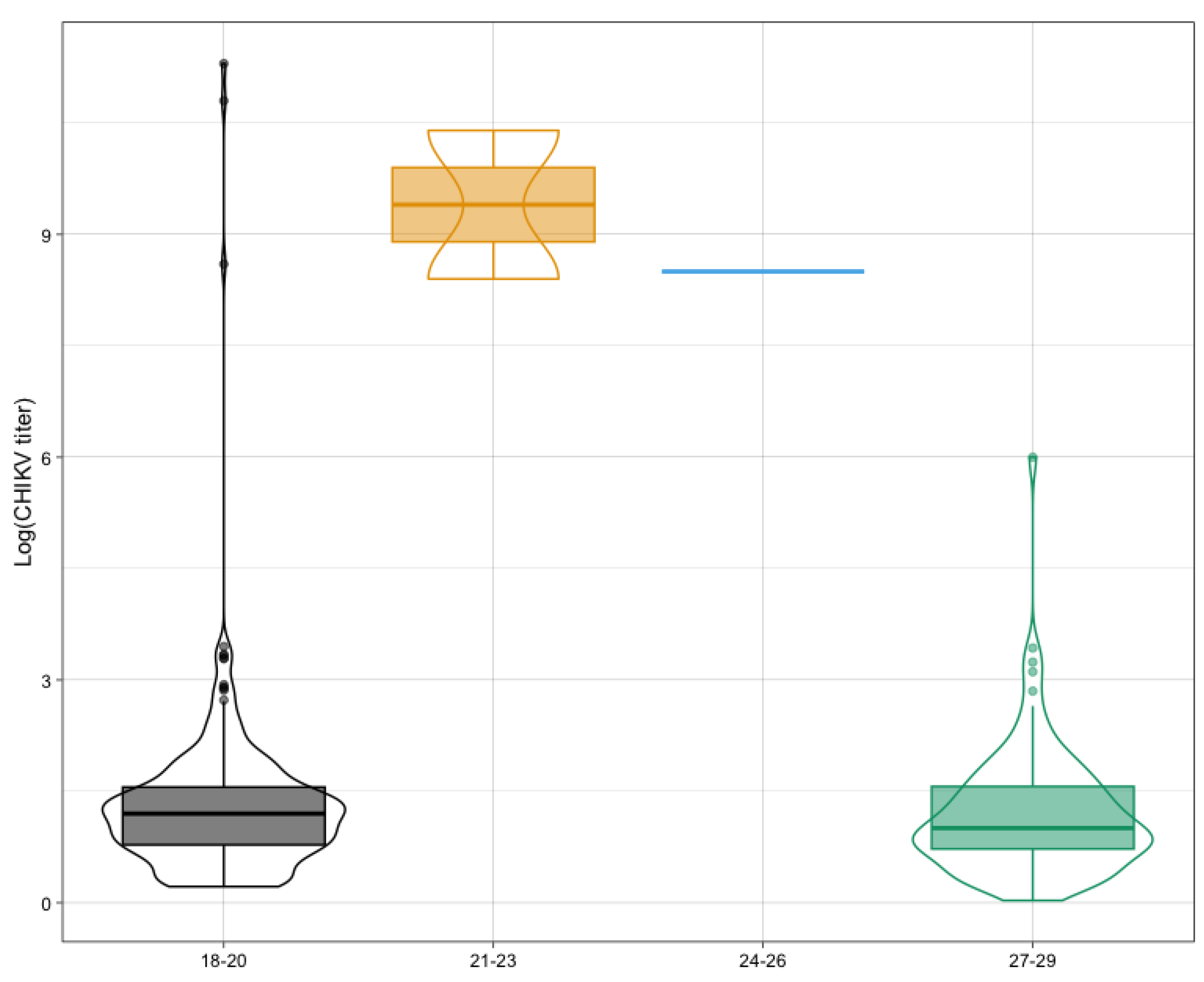
3.6. Temperature and Titer
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lumsden, W.H. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952-53. II. General description and epidemiology. Trans. R Soc. Trop. Med. Hyg. 1955, 49, 33–57. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Chikungunya. Available online: https://www.who.int/health-topics/chikungunya#tab=tab_1 (accessed on 15 November 2023).
- Ross, R.W. The Newala epidemic: III. The virus: Isolation, pathogenic properties and relationship to the epidemic. J. Hyg. 1956, 54, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Carey, D.E. Chikungunya and dengue: A Case of Mistaken Identity? J. Hist. Med. Allied Sci. 1971, XXVI, 243–262. [Google Scholar] [CrossRef]
- Farnesi, L.C.; Martins, A.J.; Valle, D.; Rezende, G.L. Embryonic development of Aedes aegypti (Diptera: Culicidae): Influence of different constant temperatures. Memórias Inst. Oswaldo Cruz 2009, 104, 124–126. [Google Scholar] [CrossRef] [PubMed]
- Tun-Lin, W.; Burkot, T.R.; Kay, B.H. Effects of temperature and larval diet on development rates and survival of the dengue vector Aedes aegypti in north Queensland, Australia. Med. Vet. Entomol. 2000, 14, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Valdez, L.D.; Sibona, G.J.; Condat, C.A. Impact of rainfall on Aedes aegypti populations. Ecol. Model. 2018, 385, 96–105. [Google Scholar] [CrossRef]
- Powers, A.M.; Logue, C.H. Changing patterns of chikungunya virus: Re-emergence of a zoonotic arbovirus. J. Gen. Virol. 2007, 88, 2363–2377. [Google Scholar] [CrossRef] [PubMed]
- De Lima Cavalcanti, T.Y.V.; Pereira, M.R.; De Paula, S.O.; Franca, R.F.D.O. A Review on Chikungunya Virus Epidemiology, Pathogenesis and Current Vaccine Development. Viruses 2022, 14, 969. [Google Scholar] [CrossRef]
- Powers, A.M.; Brault, A.C.; Tesh, R.B.; Weaver, S.C. Re-emergence of chikungunya and o’nyong-nyong viruses: Evidence for distinct geographical lineages and distant evolutionary relationships. Microbiology 2000, 81, 471–479. [Google Scholar] [CrossRef]
- Vu, D.M.; Jungkind, D.; Angelle Desiree, L. Chikungunya Virus. Clin. Lab. Med. 2017, 37, 371–382. [Google Scholar] [CrossRef]
- Weaver, S.C.; Chen, R.; Diallo, M. Chikungunya Virus: Role of Vectors in Emergence from Enzootic Cycles. Annu. Rev. Entomol. 2020, 65, 313–332. [Google Scholar] [CrossRef]
- Vazeille, M.; Moutailler, S.; Coudrier, D.; Rousseaux, C.; Khun, H.; Huerre, M.; Thiria, J.; Dehecq, J.-S.; Fontenille, D.; Schuffenecker, I.; et al. Two Chikungunya Isolates from the Outbreak of La Reunion (Indian Ocean) Exhibit Different Patterns of Infection in the Mosquito, Aedes albopictus. PLoS ONE 2007, 2, e1168. [Google Scholar] [CrossRef] [PubMed]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.-C.; Lavenir, R.; Pardigon, N.; Reynes, J.-M.; Pettinelli, F.; et al. Genome Microevolution of Chikungunya Viruses Causing the Indian Ocean Outbreak. PLoS Med. 2006, 3, e263. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A Single Mutation in Chikungunya Virus Affects Vector Specificity and Epidemic Potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef] [PubMed]
- Christofferson, R.C.; Chisenhall, D.M.; Wearing, H.J.; Mores, C.N. Chikungunya Viral Fitness Measures within the Vector and Subsequent Transmission Potential. PLoS ONE 2014, 9, e110538. [Google Scholar] [CrossRef] [PubMed]
- Leparc-Goffart, I.; Nougairede, A.; Cassadou, S.; Prat, C.; De Lamballerie, X. Chikungunya in the Americas. Lancet 2014, 383, 514. [Google Scholar] [CrossRef] [PubMed]
- Pan American Health Organization; WHO. Cases of Chikungunya Virus Disease; Pan American Health Organization: Washington, DC, USA, 2022. [Google Scholar]
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global expansion and redistribution of Aedes-borne virus transmission risk with climate change. PLoS Negl. Trop. Dis. 2019, 13, e0007213. [Google Scholar] [CrossRef]
- Rezza, G.; Nicoletti, L.; Angelini, R.; Romi, R.; Finarelli, A.; Panning, M.; Cordioli, P.; Fortuna, C.; Boros, S.; Magurano, F. Infection with chikungunya virus in Italy: An outbreak in a temperate region. Lancet 2007, 370, 1840–1846. [Google Scholar] [CrossRef]
- Adepoju, O.A.; Afinowi, O.A.; Tauheed, A.M.; Danazumi, A.U.; Dibba, L.B.; Balogun, J.B.; Flore, G.; Saidu, U.; Ibrahim, B.; Balogun, O.O. Multisectoral Perspectives on Global Warming and Vector-borne Diseases: A Focus on Southern Europe. Curr. Trop. Med. Rep. 2023, 10, 47–70. [Google Scholar] [CrossRef]
- Petersen, L.R.; Holcomb, K.; Beard, C.B. Climate change and vector-borne disease in North America and Europe. J. Health Monit. 2022, 7, 13–14. [Google Scholar]
- Robert, M.A.; Stewart-Ibarra, A.M.; Estallo, E.L. Climate change and viral emergence: Evidence from Aedes-borne arboviruses. Curr. Opin. Virol. 2020, 40, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Torales, M. Notes from the Field: Chikungunya Outbreak—Paraguay, 2022–2023. MMWR. Morb. Mortal. Wkly. Rep. 2023, 72, 636–638. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Wiggins, K.; Eastmond, B.; Ortiz, S.; Zirbel, K.; Lounibos, L.P. Diurnal temperature range and chikungunya virus infection in invasive mosquito vectors. J. Med. Entomol. 2018, 55, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Chin, P.A.; Ehrbar, D.J.; Micieli, M.V.; Fonseca, D.M.; Kramer, L.D. Differential effects of temperature and mosquito genetics determine transmissibility of arboviruses by Aedes aegypti in Argentina. Am. J. Trop. Med. Hyg. 2018, 99, 417. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, A.; Jansen, S.; Lühken, R.; Helms, M.; Pluskota, B.; Becker, N.; Kuhn, C.; Schmidt-Chanasit, J.; Tannich, E. Experimental risk assessment for chikungunya virus transmission based on vector competence, distribution and temperature suitability in Europe, 2018. Eurosurveillance 2018, 23, 1800033. [Google Scholar] [CrossRef] [PubMed]
- Mbaika, S.; Lutomiah, J.; Chepkorir, E.; Mulwa, F.; Khayeka-Wandabwa, C.; Tigoi, C.; Oyoo-Okoth, E.; Mutisya, J.; Ng’ang’a, Z.; Sang, R. Vector competence of Aedes aegypti in transmitting Chikungunya virus: Effects and implications of extrinsic incubation temperature on dissemination and infection rates. Virol. J. 2016, 13, 1–9. [Google Scholar] [CrossRef]
- Mercier, A.; Obadia, T.; Carraretto, D.; Velo, E.; Gabiane, G.; Bino, S.; Vazeille, M.; Gasperi, G.; Dauga, C.; Malacrida, A.R. Impact of temperature on dengue and chikungunya transmission by the mosquito Aedes albopictus. Sci. Rep. 2022, 12, 6973. [Google Scholar] [CrossRef]
- Nicholson, J.; Ritchie, S.; Van Den Hurk, A. Aedes albopictus (Diptera: Culicidae) as a potential vector of endemic and exotic arboviruses in Australia. J. Med. Entomol. 2014, 51, 661–669. [Google Scholar] [CrossRef]
- Westbrook, C.J.; Reiskind, M.H.; Pesko, K.N.; Greene, K.E.; Lounibos, L.P. Larval environmental temperature and the susceptibility of Aedes albopictus Skuse (Diptera: Culicidae) to Chikungunya virus. Vector-Borne Zoonotic Dis. 2010, 10, 241–247. [Google Scholar] [CrossRef]
- Wimalasiri-Yapa, B.R.; Stassen, L.; Hu, W.; Yakob, L.; McGraw, E.A.; Pyke, A.T.; Jansen, C.C.; Devine, G.J.; Frentiu, F.D. Chikungunya virus transmission at low temperature by Aedes albopictus mosquitoes. Pathogens 2019, 8, 149. [Google Scholar] [CrossRef]
- Zouache, K.; Fontaine, A.; Vega-Rua, A.; Mousson, L.; Thiberge, J.-M.; Lourenco-De-Oliveira, R.; Caro, V.; Lambrechts, L.; Failloux, A.-B. Three-way interactions between mosquito population, viral strain and temperature underlying chikungunya virus transmission potential. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141078. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Meola, M.A.; Moudy, R.M.; Kramer, L.D. Temperature, viral genetics, and the transmission of West Nile virus by Culex pipiens mosquitoes. PLoS Pathog. 2008, 4, e1000092. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Rossi, C.A.; Bailey, C.L. Effect of extrinsic incubation temperature on the ability of Aedes taeniorhynchus and Culex pipiens to transmit Rift Valley fever virus. Am. J. Trop. Med. Hyg. 1985, 34, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Z.; Lai, Z.; Zhou, T.; Jia, Z.; Gu, J.; Wu, K.; Chen, X.G. Temperature Increase Enhances Aedes albopictus Competence to Transmit Dengue Virus. Front. Microbiol. 2017, 8, 2337. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Keyel, A.C. The Role of Temperature in Transmission of Zoonotic Arboviruses. Viruses 2019, 11, 1013. [Google Scholar] [CrossRef] [PubMed]
- Christofferson, R.C.; Wearing, H.J.; Turner, E.A.; Walsh, C.S.; Salje, H.; Tran-Kiem, C.; Cauchemez, S. How do i bite thee? let me count the ways: Exploring the implications of individual biting habits of Aedes aegypti for dengue transmission. PLoS Negl. Trop. Dis. 2022, 16, e0010818. [Google Scholar] [CrossRef]
- Christofferson, R.C.; Mores, C.N. Potential for Extrinsic Incubation Temperature to Alter Interplay Between Transmission Potential and Mortality of Dengue-Infected Aedes aegypti. Environ. Health Insights 2016, 10, 119–123. [Google Scholar] [CrossRef]
- Tesla, B.; Demakovsky, L.R.; Mordecai, E.A.; Ryan, S.J.; Bonds, M.H.; Ngonghala, C.N.; Brindley, M.A.; Murdock, C.C. Temperature drives Zika virus transmission: Evidence from empirical and mathematical models. Proc. Biol. Sci. 2018, 285, 20180795. [Google Scholar] [CrossRef]
- Dye, C. Vectorial capacity: Must we measure all its components? Parasitol. Today 1986, 2, 203–209. [Google Scholar] [CrossRef]
- Massad, E.; Coutinho, F.A.B. Vectorial capacity, basic reproduction number, force of infection and all that: Formal notation to complete and adjust their classical concepts and equations. Memórias Inst. Oswaldo Cruz 2012, 107, 564–567. [Google Scholar] [CrossRef]
- Kramer, L.D.; Ciota, A.T. Dissecting vectorial capacity for mosquito-borne viruses. Curr. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Lounibos, L.P.; Kramer, L.D. Invasiveness of Aedes aegypti and Aedes albopictus and Vectorial Capacity for Chikungunya Virus. J. Infect Dis. 2016, 214, S453–S458. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, G. The Epidemiology and Control of Malaria; Oxford University Press: London, UK, 1957; pp. xv + 201 + xl + 11. [Google Scholar]
- Garrett-Jones, C. Prognosis for interruption of malaria transmission through assessment of the mosquito’s vectorial capacity. Nature 1964, 204, 1173–1175. [Google Scholar] [CrossRef] [PubMed]
- Anez, G.; Rios, M. Dengue in the United States of America: A worsening scenario? Biomed Res. Int. 2013, 2013, 678645. [Google Scholar] [CrossRef]
- Christofferson, R.C.; Mores, C.N.; Wearing, H.J. Bridging the Gap Between Experimental Data and Model Parameterization for Chikungunya Virus Transmission Predictions. J. Infect. Dis. 2016, 214, S466–S470. [Google Scholar] [CrossRef] [PubMed]
- Wearing, H.J.; Robert, M.A.; Christofferson, R.C. Dengue and chikungunya: Modelling the expansion of mosquito-borne viruses into naive populations. Parasitology 2016, 143, 860–873. [Google Scholar] [CrossRef]
- Mordecai, E.A.; Cohen, J.M.; Evans, M.V.; Gudapati, P.; Johnson, L.R.; Lippi, C.A.; Miazgowicz, K.; Murdock, C.C.; Rohr, J.R.; Ryan, S.J.; et al. Detecting the impact of temperature on transmission of Zika, dengue, and chikungunya using mechanistic models. PLoS Negl. Trop. Dis. 2017, 11, e0005568. [Google Scholar] [CrossRef]
- Gillespie, D.T. Approximate accelerated stochastic simulation of chemically reacting systems. J. Chem. Phys. 2001, 115, 1716–1733. [Google Scholar] [CrossRef]
- Clinical Evaluation & Disease (Chikungunya). Available online: https://www.cdc.gov/chikungunya/hc/clinicalevaluation.html#:~:text=The%20majority%20of%20people%20infected,%2C%201–12%20days (accessed on 15 November 2023).
- Adelman, Z.N.; Anderson, M.A.; Wiley, M.R.; Murreddu, M.G.; Samuel, G.H.; Morazzani, E.M.; Myles, K.M. Cooler temperatures destabilize RNA interference and increase susceptibility of disease vector mosquitoes to viral infection. PLoS Negl. Trop. Dis. 2013, 7, e2239. [Google Scholar] [CrossRef]
- Robison, A.; Young, M.C.; Byas, A.D.; Ruckert, C.; Ebel, G.D. Comparison of Chikungunya Virus and Zika Virus Replication and Transmission Dynamics in Aedes aegypti Mosquitoes. Am. J. Trop. Med. Hyg. 2020, 103, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Souza, T.M.L.; Morel, C.M. The COVID-19 pandemics and the relevance of biosafety facilities for metagenomics surveillance, structured disease prevention and control. Biosaf. Health 2021, 3, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rua, A.; Zouache, K.; Girod, R.; Failloux, A.B.; Lourenco-de-Oliveira, R. High level of vector competence of Aedes aegypti and Aedes albopictus from ten American countries as a crucial factor in the spread of Chikungunya virus. J. Virol. 2014, 88, 6294–6306. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.G.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Climate Central. Mosquito Disease Danger Days. In Climate Matters; Climate Central: Princeton, NJ, USA, 2018. [Google Scholar]
- Ward, H.M.; Qualls, W.A. Integrating vector and nuisance mosquito control for severe weather response. J. Am. Mosq. Control. Assoc. 2020, 36, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Caillouët, K.A.; Robertson, S.L. Temporal and spatial impacts of hurricane damage on West Nile virus transmission and human risk. J. Am. Mosq. Control. Assoc. 2020, 36, 106–119. [Google Scholar] [CrossRef]
- American Mosquito Control Association. Mosquito Management during A Public Health Emergency; American Mosquito Control Association: Sacramento, CA, USA, 2022. [Google Scholar]
- CDC. CDC and the American Mosquito Control Association Collaborate on First-ever Comprehensive Resources for Mosquito Control in Areas Affected by Natural Disasters; CDC: Atlanta, GA, USA, 2020. [Google Scholar]
- Turell, M.J.; Beaman, J.R.; Tammariello, R.F. Susceptibility of selected strains of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) to chikungunya virus. J. Med. Entomol. 1992, 29, 49–53. [Google Scholar] [CrossRef]
- Lindsey, N.P.; Staples, J.E.; Fischer, M. Chikungunya Virus Disease among Travelers-United States, 2014–2016. Am. J. Trop. Med. Hyg. 2018, 98, 192–197. [Google Scholar] [CrossRef]
- Rowe, D.; McDermott, C.; Veliz, Y.; Kerr, A.; Whiteside, M.; Coss, M.; Huff, C.; Leal, A.; Kopp, E.; LaCrue, A.; et al. Dengue Outbreak Response during COVID-19 Pandemic, Key Largo, Florida, USA, 2020. Emerg. Infect. Dis. 2023, 29, 1643–1647. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Imported dengue—Florida, 1997–1998. Can. Commun. Dis. Rep. 2000, 26, 77–79. [Google Scholar]
- Alto, B.W.; Smartt, C.T.; Shin, D.; Bettinardi, D.; Malicoate, J.; Anderson, S.L.; Richards, S.L. Susceptibility of Florida Aedes aegypti and Aedes albopictus to dengue viruses from Puerto Rico. J. Vector. Ecol. 2014, 39, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Sharp, T.M.; Morris, S.; Morrison, A.; de Lima Corvino, D.; Santiago, G.A.; Shieh, W.J.; Rico, E.; Kopp, E.; Munoz-Jordan, J.L.; Marttos, A.; et al. Fatal Dengue Acquired in Florida. N. Engl. J. Med. 2021, 384, 2257–2259. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.L.; Santiago, G.A.; Abeyta, R.; Hinojosa, S.; Torres-Velasquez, B.; Adam, J.K.; Evert, N.; Caraballo, E.; Hunsperger, E.; Munoz-Jordan, J.L.; et al. Reemergence of Dengue in Southern Texas, 2013. Emerg. Infect Dis. 2016, 22, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.J.; Miller, B.R.; Gubler, D.J. Vector competence of Aedes albopictus from Houston, Texas, for dengue serotypes 1 to 4, yellow fever and Ross River viruses. J Am Mosq Control Assoc 1987, 3, 460–465. [Google Scholar] [PubMed]
- Brunkard, J.M.; Robles Lopez, J.L.; Ramirez, J.; Cifuentes, E.; Rothenberg, S.J.; Hunsperger, E.A.; Moore, C.G.; Brussolo, R.M.; Villarreal, N.A.; Haddad, B.M. Dengue fever seroprevalence and risk factors, Texas-Mexico border, 2004. Emerg. Infect. Dis. 2007, 13, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Ramin, B.; Svoboda, T. Health of the homeless and climate change. J. Urban Health 2009, 86, 654–664. [Google Scholar] [CrossRef] [PubMed]
- de Jesus Crespo, R.; Rogers, R.E. Habitat Segregation Patterns of Container Breeding Mosquitos: The Role of Urban Heat Islands, Vegetation Cover, and Income Disparity in Cemeteries of New Orleans. Int. J. Environ. Res. Public Health 2021, 19, 245. [Google Scholar] [CrossRef]
- City of New Orleans Health Department. Available online: https://nola.gov/health-department/homelessness-en/ (accessed on 20 May 2023).
- de Sousa, T.; Andrichik, A.; Ceullar, M.; Marson, J.; Prestera, E.; Rush, K. The 2022 Annual Homelessness Assessment REport (AHAR) to Congress; U.S. Department of Housing and Urban Development: Washington, DC, USA, 2022. [Google Scholar]
- de Jesus Crespo, R.; Harrison, M.; Rogers, R.; Vaeth, R. Mosquito Vector Production across Socio-Economic Divides in Baton Rouge, Louisiana. Int. J. Environ. Res. Public Health 2021, 18, 1420. [Google Scholar] [CrossRef]
- Yitbarek, S.; Chen, K.; Celestin, M.; McCary, M. Urban mosquito distributions are modulated by socioeconomic status and environmental traits in the USA. Ecol. Appl. 2023, 33, e2869. [Google Scholar] [CrossRef]
- Hotez, P.J.; Murray, K.O.; Buekens, P. The Gulf Coast: A new American underbelly of tropical diseases and poverty. PLoS Negl. Trop. Dis. 2014, 8, e2760. [Google Scholar] [CrossRef] [PubMed]
- Agha, S.B.; Chepkorir, E.; Mulwa, F.; Tigoi, C.; Arum, S.; Guarido, M.M.; Ambala, P.; Chelangat, B.; Lutomiah, J.; Tchouassi, D.P.; et al. Vector competence of populations of Aedes aegypti from three distinct cities in Kenya for chikungunya virus. PLoS Negl. Trop. Dis. 2017, 11, e0005860. [Google Scholar] [CrossRef] [PubMed]
- Heath, C.J.; Grossi-Soyster, E.N.; Ndenga, B.A.; Mutuku, F.M.; Sahoo, M.K.; Ngugi, H.N.; Mbakaya, J.O.; Siema, P.; Kitron, U.; Zahiri, N.; et al. Evidence of transovarial transmission of Chikungunya and Dengue viruses in field-caught mosquitoes in Kenya. PLoS Negl. Trop. Dis. 2020, 14, e0008362. [Google Scholar] [CrossRef] [PubMed]
- Mease, L.E.; Coldren, R.L.; Musila, L.A.; Prosser, T.; Ogolla, F.; Ofula, V.O.; Schoepp, R.J.; Rossi, C.A.; Adungo, N. Seroprevalence and distribution of arboviral infections among rural Kenyan adults: A cross-sectional study. Virol. J. 2011, 8, 371. [Google Scholar] [CrossRef] [PubMed]
- Inziani, M.; Adungo, F.; Awando, J.; Kihoro, R.; Inoue, S.; Morita, K.; Obimbo, E.; Onyango, F.; Mwau, M. Seroprevalence of yellow fever, dengue, West Nile and chikungunya viruses in children in Teso South Sub-County, Western Kenya. Int. J. Infect. Dis. 2020, 91, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Bisanzio, D.; Mutuku, F.; Ndenga, B.; Grossi-Soyster, E.N.; Jembe, Z.; Maina, P.W.; Chebii, P.K.; Ronga, C.O.; Okuta, V.; et al. Spatiotemporal overlapping of dengue, chikungunya, and malaria infections in children in Kenya. BMC Infect. Dis. 2023, 23, 183. [Google Scholar] [CrossRef] [PubMed]
- Mordecai, E.A.; Ryan, S.J.; Caldwell, J.M.; Shah, M.M.; LaBeaud, A.D. Climate change could shift disease burden from malaria to arboviruses in Africa. Lancet Planet Health 2020, 4, e416–e423. [Google Scholar] [CrossRef]
- Christofferson, R.C.; Cormier, S.A. Beyond the Unknown: A Broad Framing for Preparedness for Emerging Infectious Threats. Am. J. Trop. Med. Hyg. 2022, 107, 1159–1161. [Google Scholar] [CrossRef]
- Ragavan, M.I.; Marcil, L.E.; Garg, A. Climate Change as a Social Determinant of Health. Pediatrics 2020, 145, e20193169. [Google Scholar] [CrossRef]
- Benedict, M.Q.; Levine, R.S.; Hawley, W.A.; Lounibos, L.P. Spread of The Tiger: Global Risk of Invasion by The Mosquito Aedes albopictus. Vector-Borne Zoonotic Dis. 2007, 7, 76–85. [Google Scholar] [CrossRef]
- Khormi, H.M.; Kumar, L. Climate change and the potential global distribution of Aedes aegypti: Spatial modelling using geographical information system and CLIMEX. Geospat. Health 2014, 8, 405. [Google Scholar] [CrossRef]
- Rocklov, J.; Dubrow, R. Climate change: An enduring challenge for vector-borne disease prevention and control. Nat. Immunol. 2020, 21, 479–483. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Temperature | Longevity (days) | Biting Rate (1/day) |
|---|---|---|---|
| Ae. aegypti | 19 °C | 26.6 | 0.09 |
| 22 °C | 29.2 | 0.15 | |
| 25 °C | 29.3 | 0.22 | |
| 28 °C | 26.5 | 0.28 | |
| 31 °C | 21.1 | 0.32 | |
| Ae. albopictus | 19 °C | 95.0 | 0.14 |
| 22 °C | 110.5 | 0.20 | |
| 25 °C | 102.5 | 0.25 | |
| 28 °C | 68.9 | 0.30 | |
| 31 °C | 13.4 | 0.33 |
| Parameter | Value |
|---|---|
| Mosquito Emergence Rate | 5000/7 days |
| P (Transmission|Bite) from mosquito to human | 1 |
| Human Incubation Period | 5 days [53] |
| Human Infectious Period | 8.5 days [53] |
| Species | EITClass | Fit | Parameters |
|---|---|---|---|
| Ae. aegypti | 18–20 | Linear | Int = 5.269 Slope = 0.195 |
| 24–26 | Linear | Int = 21.910 Slope = 1.019 | |
| 30–32 | Linear | Int = 4.767 Slope = 1.667 | |
| Ae. albopictus | 18–20 * | Linear | Int = 47.185 Slope = 1.713 |
| 21–23 * | Exponential | Start = 53.059 Rate = −0.0096 | |
| 27–29 * | Exponential | Start = 64.044 Rate = −0.014 |
| Mosquito Species | EIT Class | EIPMAX (Dissemination %) | EIP50 |
|---|---|---|---|
| Ae. aegypti | 18–20 | 21 (10.6%) | NA |
| 24–26 | 21 (45.7%) | NA | |
| 30–32 | 13 (27.7%) | NA | |
| Ae. albopictus | 18–20 | 21 (83.3%) | 1.643 |
| 21–23 | 10 (100%) | 6.176 | |
| 27–29 | 4 (100%) | 17.213 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christofferson, R.C.; Turner, E.A.; Peña-García, V.H. Identifying Knowledge Gaps through the Systematic Review of Temperature-Driven Variability in the Competence of Aedes aegypti and Ae. albopictus for Chikungunya Virus. Pathogens 2023, 12, 1368. https://doi.org/10.3390/pathogens12111368
Christofferson RC, Turner EA, Peña-García VH. Identifying Knowledge Gaps through the Systematic Review of Temperature-Driven Variability in the Competence of Aedes aegypti and Ae. albopictus for Chikungunya Virus. Pathogens. 2023; 12(11):1368. https://doi.org/10.3390/pathogens12111368
Chicago/Turabian StyleChristofferson, Rebecca C., Erik A. Turner, and Víctor Hugo Peña-García. 2023. "Identifying Knowledge Gaps through the Systematic Review of Temperature-Driven Variability in the Competence of Aedes aegypti and Ae. albopictus for Chikungunya Virus" Pathogens 12, no. 11: 1368. https://doi.org/10.3390/pathogens12111368