
Ticks Infesting Dogs in Khyber Pakhtunkhwa, Pakistan: Detailed Epidemiological and Molecular Report
 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Consent
2.2. Study Location
2.3. Study Design, Tick Sampling Strategy, and Morpho-taxonomic Identification
2.4. Tick Genomic DNA Extraction and Target Genes Amplification
2.5. Amplicons Purification and Sequencing
2.6. Phylogenetic Analysis
2.7. Statistical Analysis of Empirical Data
3. Results
3.1. Host Demographic Profile
3.2. Prevalence and Distribution of Ixodid Ticks
3.2.1. Total and District-wise Prevalence of Ticks
3.2.2. Tick Prevalence with Respect to Host Demography
3.2.3. Tick Prevalence with Respect to Host Management Practices
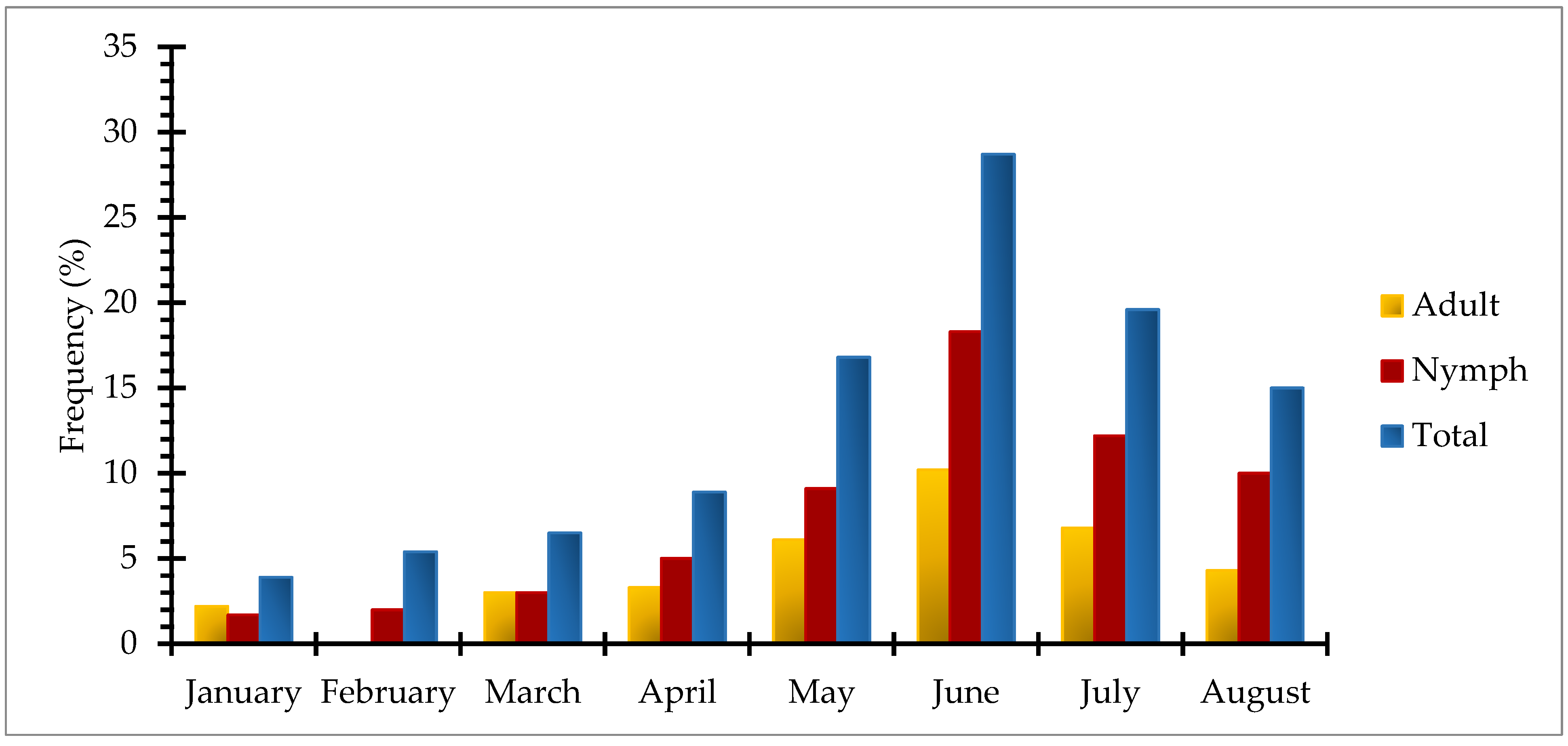
3.2.4. Spatio-temporal Distribution of Tick Species
3.3. Potential Risk Factors for Tick Infestation
3.4. Molecular Attributes of Phylogenetic Markers of Collected Tick Species
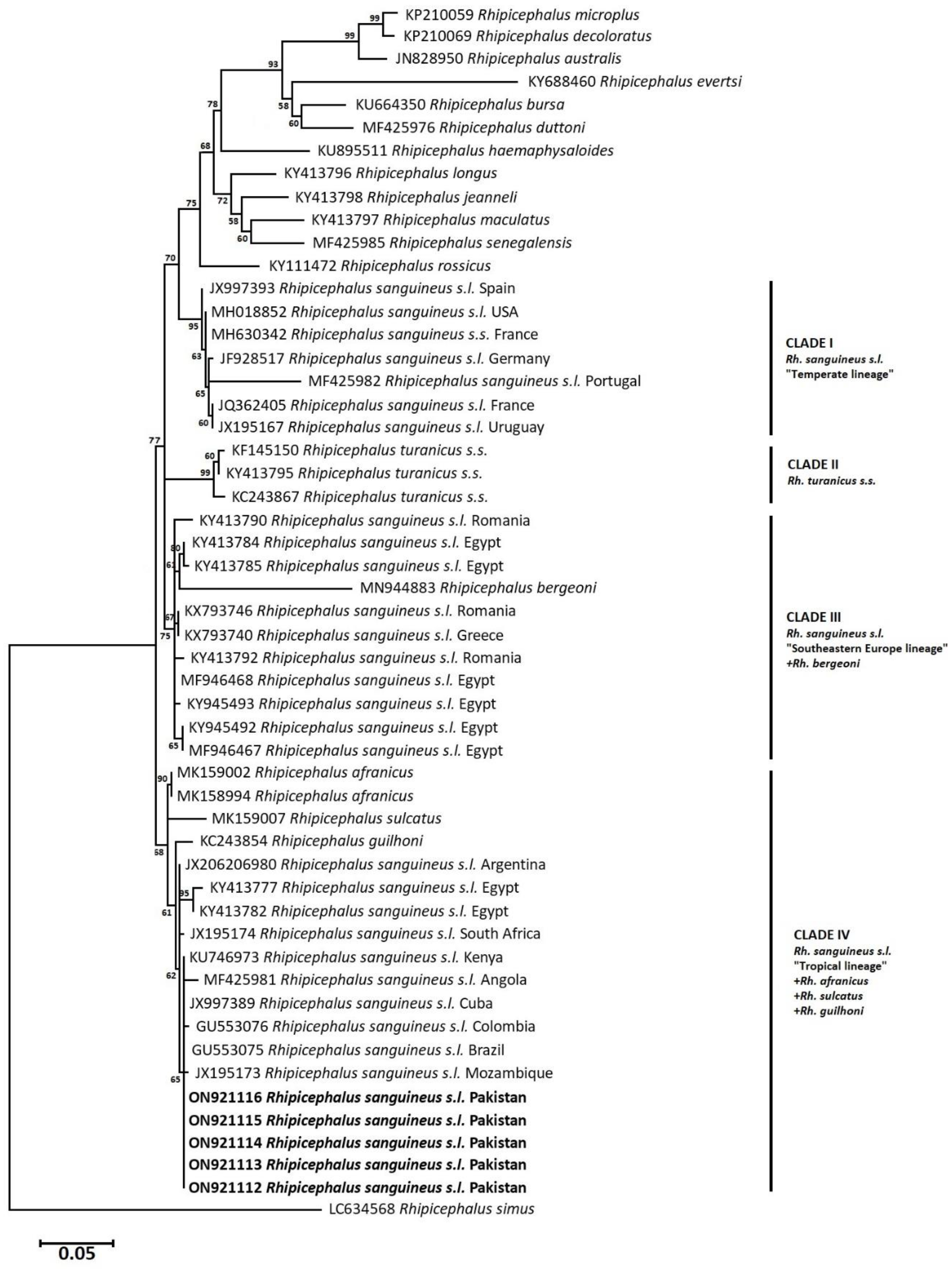
3.5. Phylogenetic Analysis: Neighbor-Joining
3.6. Phylogenetic Analysis: Maximum Likelihood (Rh. sanguineus complex)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- De la Fuente, J.; Antunes, S.; Bonnet, S.; Cabezas-Cruz, A.; Domingos, A.G.; Estrada-Peña, A.; Johnson, N.; Kocan, K.M.; Mansfield, K.L.; Nijhof, A.M.; et al. Tick-pathogen interactions and vector competence: Identification of molecular drivers for tick-borne diseases. Front. Cell. Infect. Microbiol. 2017, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Moumouni, P.F.A.; Thekisoe, O.; Gao, Y.; Liu, M.; Li, J.; May Galon, E.; Efstratiou, A.; Wang, G.; Jirapattharasate, C.; et al. Genetic characterization of tick-borne pathogens in ticks infesting cattle and sheep from three South African provinces. Ticks Tick-Borne Dis. 2019, 10, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Mossaad, E.; Gaithuma, A.; Mohamed, Y.O.; Suganuma, K.; Umemiya-Shirafuji, R.; Ohari, Y.; Salim, B.; Liu, M.; Xuan, X. Molecular Characterization of Ticks and Tick-Borne Pathogens in Cattle from Khartoum State and East Darfur State, Sudan. Pathogens 2021, 10, 580. [Google Scholar] [CrossRef] [PubMed]
- Elelu, N.; Bankole, A.A.; Daphne, H.P.; Rabiu, M.; Ola-Fadunsin, S.D.; Ambali, H.M.; Cutler, S.J. Molecular characterisation of Rhipicephalus sanguineus sensu lato ticks from domestic dogs in Nigeria. Vet. Med. Sci. 2022, 8, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Powell, L.; Edwards, K.M.; McGreevy, P.; Bauman, A.; Podberscek, A.; Neilly, B.; Sherrington, C.; Stamatakis, E. Companion dog acquisition and mental well-being: A community-based three-arm controlled study. BMC Public Health 2019, 19, 1–10. [Google Scholar] [CrossRef]
- Losson, B.; Daelemans, A.C.; De Cat, A.; Madder, M.; Saegerman, C.; Heyman, P.; Lempereur, L. Ticks and associated pathogens collected from dogs and cats in Belgium. Parasites Vectors 2013, 6, 1–9. [Google Scholar]
- Guglielmone, A.A.; Petney, T.N.; Robbins, R.G. Ixodidae (Acari: Ixodoidea): Descriptions and redescriptions of all known species from 1758 to December 31, 2019. Zootaxa 2020, 487, 1–322. [Google Scholar] [CrossRef]
- Hoogstraal, H. African Ixodoidea. VoI. I. Ticks of the Sudan (with Special Reference to Equatoria Province and with Preliminary Reviews of the Genera Boophilus, Margaropus, and Hyalomm; Naval Medical Research Unit Three: Cairo, Egypt, 1956; p. 1101. [Google Scholar]
- Šlapeta, J.; Chandra, S.; Halliday, B. The “tropical lineage” of the brown dog tick Rhipicephalus sanguineus sensu lato identified as Rhipicephalus linnaei. Int. J. Parasitol 2021, 51, 431–436. [Google Scholar] [CrossRef]
- Walker, A.R. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: London, UK, 2003. [Google Scholar]
- Chitimia-Dobler, L.; Langguth, J.; Pfeffer, M.; Kattner, S.; Küpper, T.; Friese, D.; Dobler, G.; Guglielmone, A.A.; Nava, S. Genetic analysis of Rhipicephalus sanguineus sensu lato ticks parasites of dogs in Africa north of the Sahara based on mitochondrial DNA sequences. Vet. Parasitol. 2017, 239, 1–6. [Google Scholar] [CrossRef]
- Nava, S.; Beati, L.; Venzal, J.M.; Labruna, M.B.; Szabó, M.P.; Petney, T.; Saracho-Bottero, M.N.; Tarragona, E.L.; Dantas-Torres, F.; Santos Silva, M.M.; et al. Rhipicephalus sanguineus (Latreille, 1806): Neotype designation, morphological re-description of all parasitic stages and molecular characterization. Ticks Tick-Borne Dis. 2018, 9, 1573–1585. [Google Scholar] [CrossRef]
- Dantas-Torres, F. Biology and ecology of the brown dog tick, Rhipicephalus sanguineus. Parasit Vectors 2010, 3, 26. [Google Scholar] [CrossRef]
- Estrada-Pena, A. Tick-borne pathogens, transmission rates and climate change. Front. Biosci. Landmark 2009, 14, 2674–2687. [Google Scholar] [CrossRef] [Green Version]
- Sajid, M.S.; Iqbal, Z.; Khan, M.N.; Muhammad, G.; Khan, M.K. Prevalence and associated risk factors for bovine tick infestation in two districts of lower Punjab, Pakistan. Prev. Vet. Med. 2009, 92, 386–391. [Google Scholar] [CrossRef]
- Iqbal, A.; Sajid, M.S.; Khan, M.N.; Khan, M.K. Frequency distribution of hard ticks (Acari: Ixodidae) infesting bubaline population of district Toba Tek Singh, Punjab, Pakistan. Parasitol. Res. 2013, 112, 535–541. [Google Scholar] [CrossRef]
- Kamran, K. Ticks Prevalence and Possible Risk Factors Assessment on Domestic Dogs in Quetta District Balochistan, Pakistan. Egypt. J. Vet. Sci. 2021, 52, 87–94. [Google Scholar] [CrossRef]
- Smith, F.D.; Ballantyne, R.; Morgan, E.R.; Wall, R. Prevalence, distribution and risk associated with tick infestation of dogs in Great Britain. Med. Vet. Entomol. 2011, 25, 377–384. [Google Scholar] [CrossRef]
- Lv, J.; Wu, S.; Zhang, Y.; Chen, Y.; Feng, C.; Yuan, X. Assessment of four DNA fragments (COI, 16S rDNA, ITS2, 12S rDNA) for species identification of the Ixodida (Acari: Ixodida). Parasites Vectors 2014, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duron, O.; Noël, V.; McCoy, K.D.; Bonazzi, M.; Sidi-Boumedine, K.; Morel, O.; Vavre, F.; Zenner, L.; Jourdain, E.; Durand, P.; et al. The recent evolution of a maternally-inherited endosymbiont of ticks led to the emergence of the Q fever pathogen, Coxiella burnetii. PLoS Pathog. 2015, 11, e1004892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasi, K.K.; Sas, M.A.; Sauter-Louis, C.; von Arnim, F.; Gethmann, J.M.; Schulz, A.; Wernike, K.; Groschup, M.H.; Conraths, F.J. Epidemiological investigations of Crimean-Congo haemorrhagic fever virus infection in sheep and goats in Balochistan, Pakistan. Ticks Tick-Borne Dis. 2020, 11, 101324. [Google Scholar] [CrossRef]
- Kasi, K.K.; Sas, M.A.; Sauter-Louis, C.; von Arnim, F.; Gethmann, J.M.; Schulz, A.; Wernike, K.; Groschup, M.H.; Conraths, F.J. Crimean-Congo haemorrhagic fever virus in ticks collected from livestock in Balochistan, Pakistan. Transbound. Emerg. Dis. 2020, 67, 1543–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeb, J.; Szekeres, S.; Takács, N.; Kontschán, J.; Shams, S.; Ayaz, S.; Hornok, S. Genetic diversity, piroplasms and trypanosomes in Rhipicephalus microplus and Hyalomma anatolicum collected from cattle in northern Pakistan. Exp. Appl. Acarol. 2019, 79, 233–243. [Google Scholar] [CrossRef]
- Ullah, R.; Shams, S.; Khan, M.A.; Ayaz, S.; Akbar, N.u.; Din, Q.u.; Khan, A.; Leon, R.; Zeb, J. Epidemiology and molecular characterization of Theileria annulata in cattle from central Khyber Pakhtunkhwa, Pakistan. PloS ONE 2021, 16, e0249417. [Google Scholar] [CrossRef]
- Sultan, S.; Zeb, J.; Ayaz, S.; Rehman, S.U.; Hussain, M.; Senbill, H.; Husain, S.; Sparagano, O.A. Epidemiologic profile of hard ticks and molecular characterization of Rhipicephalus microplus infesting cattle in central part of Khyber Pakhtunkhwa, Pakistan. Parasitol. Res. 2022, 121, 2481–2493. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Khan, M.A.; Zahid, H.; Yaseen, P.M.; Qayash Khan, M.; Nawab, J.; Ur Rehman, Z.; Ateeq, M.; Khan, S.; Ibrahim, M. Seasonal dynamics, record of ticks infesting humans, wild and domestic animals and molecular phylogeny of Rhipicephalus microplus in Khyber Pakhtunkhwa Pakistan. Front. Physiol. 2019, 10, 793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-Peña, A.; Bouattour, A.; Camicas, J.L.; Walker, A.R. Ticks of Domestic Animals in the Mediterranean Region; University of Zaragoza: Zaragoza, Spain, 2004; p. 131. [Google Scholar]
- Walker, J.B.; Keirans, J.E.; Horak, I.G. The Genus Rhipicephalus (Acari, Ixodidae): A Guide to the Brown Ticks of the World; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Prates, L.; Otomura, F.H.; Mota, L.T.; Toledo, M.J.d.O. Impact of antiparasitic treatment on the prevalence of ectoparasites in dogs from an indigenous territory, state of Parana, Brazil. Rev. Patol. Trop. 2013, 42, 339–351. [Google Scholar]
- Hornok, S.; Szőke, K.; Kováts, D.; Estok, P.; Görföl, T.; Boldogh, S.A.; Takács, N.; Kontschán, J.; Földvári, G.; Barti, L.; et al. DNA of piroplasms of ruminants and dogs in ixodid bat ticks. PLoS ONE 2016, 11, e0167735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Black, W.C.; Piesman, J. Phylogeny of hardand soft-tick taxa (Acari: Ixodida) based on mitochon drial 16S rDNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Lanave, C.; Preparata, G.; Sacone, C.; Serio, G. A new method for calculating evolutionary substitution rates. J. Mol. Evol. 1984, 20, 86–93. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+ C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [PubMed] [Green Version]
- Rehman, A.; Conraths, F.J.; Sauter-Louis, C.; Krücken, J.; Nijhof, A.M. Epidemiology of tick-borne pathogens in the semi-arid and the arid agro-ecological zones of Punjab province, Pakistan. Transbound. Emerg. Dis. 2019, 66, 526–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghafar, A.; Cabezas-Cruz, A.; Galon, C.; Obregon, D.; Gasser, R.B.; Moutailler, S.; Jabbar, A. Bovine ticks harbour a diverse array of microorganisms in Pakistan. Parasites Vectors 2020, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ghafar, A.; Gasser, R.B.; Rashid, I.; Ghafoor, A.; Jabbar, A. Exploring the prevalence and diversity of bovine ticks in five agro-ecological zones of Pakistan using phenetic and genetic tools. Ticks Tick-Borne Dis. 2020, 11, 101472. [Google Scholar] [CrossRef] [PubMed]
- Rooman, M.; Assad, Y.; Tabassum, S.; Sultan, S.; Ayaz, S.; Khan, M.F.; Khan, S.N.; Ali, R. A cross-sectional survey of hard ticks and molecular characterization of Rhipicephalus microplus parasitizing domestic animals of Khyber Pakhtunkhwa, Pakistan. PloS ONE 2021, 16, e0255138. [Google Scholar] [CrossRef]
- Khan, M.; Hayar, C.; Iqbal, Z.; Hayat, B. Prevalence of ticks on livestock in Faisalabad (Pakistan). Pak. Vet. J. 1993, 13, 182. [Google Scholar]
- Zaman, A. K: Prevalence and chemotherapy of ticks among cattle in various refugee camps at Hangu area. Master’s Thesis, NWFP Pakistan, Agriculture University, Peshawar, Pakistan, 1997. [Google Scholar]
- Durrani, A.Z.; Kamal, N. Identification of ticks and detection of blood protozoa in friesian cattle by polmerase chain reacton test and estimation of blood parameters in district Kasur, Pakistan. Trop. Anim. Health Prod. 2008, 40, 441–447. [Google Scholar] [CrossRef]
- Sajid, M.S.; Iqbal, Z.; Khan, M.N.; Muhammad, G. Point prevalence of hard ticks (Ixodids) infesting domestic ruminants of lower Punjab, Pakistan. Int. J. Agric. Biol. 2008, 10, 349–351. [Google Scholar]
- Irshad, N.; Qayyum, M.; Hussain MQasim Khan, M. Prevalence of Tick Infestation and Theileriosis in Sheep and Goats. Pak. Vet. J. 2010, 30, 178–180. [Google Scholar]
- Ali, Z.; Maqbool, A.; Muhammad, K.; Khan, M.; Younis, M. Prevalence of Theileria annulata infected hard ticks of cattle and buffalo in Punjab, Pakistan. DNA 2013, 862, 846. [Google Scholar]
- Rehman, A.; Ard, M.N.; Carola, S.; Birgit, S.; Christoph, S.; Franz, J.C. Distribution of ticks infesting ruminants and risk factors associated with high tick prevalence in livestock farms in the semi-arid and arid agro-ecological zones of Pakistan. Parasites Vectors 2017, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zeb, J.; Shams, S.; Ayaz, S.; Din, I.U.; Khan, A.; Adil, N.; Raza, A. Epidemiology of ticks and molecular characterization of Rhipicephalus microplus in cattle population in North-Western Pakistan. Int. J. Acarol. 2020, 46, 335–343. [Google Scholar] [CrossRef]
- Ghafar, A.; Khan, A.; Cabezas-Cruz, A.; Gauci, C.G.; Niaz, S.; Ayaz, S.; Mateos-Hernandez, L.; Galon, C.; Nasreen, N.; Moutailler, S.; et al. An assessment of the molecular diversity of ticks and tick-borne microorganisms of small ruminants in Pakistan. Microorganisms 2020, 8, 1428. [Google Scholar] [CrossRef] [PubMed]
- Ekanem, M.; Mbagwu, H.; Opara, K.; Agbata, Q. Ticks infestation of domestic dogs (Canis familiaris lupus) in Uyo, Akwa Ibom state, Nigeria. World J. Appl. Sci. Technol. 2010, 2, 191–196. [Google Scholar]
- Jafri, S.A.; Rabbani, M. Prevalence of canine diseases in Lahore area. Pak Vet, J. 1999, 19, 40–42. [Google Scholar]
- Odeniran, P. Prevalence and Risk Factors of Tick Infestation in Dogs in Ibadan, Nigeria. Afr. J. Biomed. Res. 2021, 24, 135–140. [Google Scholar]
- Abdulkareem, B.O.; Christy, A.L.; Samuel, U.U. Prevalence of ectoparasite infestations in owned dogs in Kwara State, Nigeria. Parasite Epidemiol. Control. 2019, 4, e00079. [Google Scholar] [CrossRef]
- James-Rugu, N.; Jidayi, S. A survey on the ectoparasites of some livestock from some areas of Borno and Yobe States. Niger. Vet. J. 2004, 25, 48–55. [Google Scholar] [CrossRef]
- Földvári, G.; Farkas, R. Ixodid tick species attaching to dogs in Hungary. Vet. Parasitol. 2005, 129, 125–131. [Google Scholar] [CrossRef]
- Duscher, G.G.; Feiler, A.; Leschnik, M.; Joachim, A. Seasonal and spatial distribution of ixodid tick species feeding on naturally infested dogs from Eastern Austria and the influence of acaricides/repellents on these parameters. Parasites Vectors 2013, 6, 76. [Google Scholar] [CrossRef]
- Moghaddar, S.; Shorigeh, J.; Gastrodashty, A. Prevalence of ectoparasites and its seasonal prevalence in dogs in Shiraz (Iran). XII Natl. Congr. Vet. Parasitol. 2001, 62. [Google Scholar]
- Raut, P.; Maske, D.; Jayraw, A.; Sonkusale, V. Ectoparasitism in dogs from the eastern zone of Maharashtra state. J. Parasit. Dis. 2006, 30, 138–141. [Google Scholar]
- Tinoco-Gracia, L.; Quiroz-Romero, H.; Quintero-Martínez, M.T.; Rentería-Evangelista, T.B.; González-Medina, Y.; Barreras-Serrano, A.; Hori-Oshima, M.H.; Moro, J.V. Prevalence of Rhipicephalus sanguineus ticks on dogs in a region on the Mexico-USA border. Vet. Rec. 2009, 164, 59. [Google Scholar] [CrossRef] [PubMed]
- Sahu, A.; Mohanty, B.; Panda, M.R.; Sardar, K.K.; Dehuri, M. Prevalence of tick infestation in dogs in and around Bhubaneswar. Vet. World 2013, 6, 982. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.; Godara, R.; Yadav, A.; Katoch, R. Prevalence of ticks in sheep and goats of Jammu region. Ind. J. Small Rum. 2018, 24, 183–186. [Google Scholar] [CrossRef]
- Lüssenhop, J.; Bäumer, W.; Kietzmann, M.; Schnieder, T.; Wolken, S. Dynamics of distribution and efficacy of different spot-on permethrin formulations in dogs artificially infested with Dermacentor reticulatus. Parasites Vectors 2011, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Széll, Z.; Sréter-Lancz, Z.; Márialigeti, K.; Sréter, T. Temporal distribution of Ixodes ricinus, Dermacentor reticulatus and Haemaphysalis concinna in Hungary. Vet. Parasitol. 2006, 141, 377–379. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Yazid Abdad, M.; Stenos, J.; Bitam, I.; Fournier, P.E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [Green Version]
- Randolph, S.E.; Storey, K. Impact of microclimate on immature tick-rodent host interactions (Acari: Ixodidae): Implications for parasite transmission. J. Med. Entomol. 1999, 36, 741–748. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Otranto, D. Rhipicephalus sanguineus sl (Latreille, 1806) (Figs. 127–129). In Ticks of Europe and North Africa; Springer: Berlin/Heidelberg, Germany, 2017; pp. 323–327. [Google Scholar]
- Senbill, H.; Tanaka, T.; Karawia, D.; Rahman, S.; Zeb, J.; Sparagano, O.; Baruah, A. Morphological identification and molecular characterization of economically important ticks (Acari: Ixodidae) from North and North–Western Egypt. Acta Trop. 2022, 231, 106438. [Google Scholar] [CrossRef]
- Khan, S.S.; Ahmed, H.; Afzal, M.S.; Khan, M.R.; Birtles, R.J.; Oliver, J.D. Epidemiology, distribution and identification of ticks on livestock in Pakistan. Int. J. Environ. Res. Public Health 2022, 19, 3024. [Google Scholar] [CrossRef]
- Moraes-Filho, J.; Marcili, A.; Nieri-Bastos, F.A.; Richtzenhain, L.J.; Labruna, M.B. Genetic analysis of ticks belonging to the Rhipicephalus sanguineus group in Latin America. Acta Trop. 2011, 117, 51–55. [Google Scholar] [CrossRef]
- Nava, S.; Mastropaolo, M.; Venzal, J.M.; Mangold, A.J.; Guglielmone, A.A. Mitochondrial DNA analysis of Rhipicephalus sanguineus sensu lato (Acari: Ixodidae) in the Southern Cone of South America. Vet. Parasitol. 2012, 190, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Latrofa, M.S.; Ramos, R.A.N.; Lia, R.P.; Capelli, G.; Parisi, A.; Porretta, D.; Urbanelli, S.; Otranto, D. Biological compatibility between two temperate lineages of brown dog ticks, Rhipicephalus sanguineus (sensu lato). Parasit Vectors 2018, 11, 398. [Google Scholar] [CrossRef] [PubMed]
- Hoogstraal, H.; Wassef, H.; Büttiker, W. Ticks (Acarina) of Saudi Arabia. Fam. Argasidae, Ixodidae. Fauna Saudi Arab. 1981, 3, 25–110. [Google Scholar]
- Apanaskevich, D. Host-parasite relationships of the genus Hyalomma Koch, 1844 (Acari, Ixodidae) and their connection with microevolutionary process. Parazitologiia 2004, 38, 515–523. [Google Scholar]
- Bakheit, M.A.; Latif, A.A.; Vatansever, Z.; Seitzer, U.; Ahmed, J. The huge risks due to Hyalomma ticks. Arthropods as Vectors of Emerging Diseases; Springer: Berlin/Heidelberg, Germany, 2012; pp. 167–194. [Google Scholar]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa: A Guide to Species Identification; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Perveen, N.; Muzaffar, S.B.; Al-Deeb, M.A. Prevalence, Distribution, and Molecular Record of Four Hard Ticks from Livestock in the United Arab Emirates. Insects 2021, 12, 1016. [Google Scholar] [CrossRef]
- Kaiser, M.; Hoogstraal, H. The Hyalomma ticks (Ixodoidea, Ixodidae) of Pakistan, India, and Ceylon, with keys to subgenera and species. Acarologia 1964, 6, 257–286. [Google Scholar]
- Begum, F.; Wisseman Jr., C.; Casals, J. Tick-borne viruses of West Pakistan: Ii. Hazara virus, a new agent isolated from Ixodes redikorzevi ticks from the Kaghan valley, W. Pakistan. Am. J. Epidemiol. 1970, 92, 192–194. [Google Scholar] [CrossRef]
- Choubdar, N.; Karimian, F.; Koosha, M.; Nejati, J.; Oshaghi, M.A. Hyalomma spp. ticks and associated Anaplasma spp. and Ehrlichia spp. on the Iran-Pakistan border. Parasites Vectors 2021, 14, 1–8. [Google Scholar] [CrossRef]
- Salih, D.; Hassan, S.; El Hussein, A.; Jongejan, F. Preliminary survey of ticks (Acari: Ixodidae) on cattle in northern Sudan. Onderstepoort J. Vet. Res. 2004, 71, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apanaskevich, D.A.; Schuster, A.L.; Horak, I.G. The genus Hyalomma: VII. Redescription of all parasitic stages of H. (Euhyalomma) dromedarii and H. (E.) schulzei (Acari: Ixodidae). J. Med. Entomol. 2008, 45, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Shemshad, K.; Rafinejad, J.; Kamali, K.; Piazak, N.; Sedaghat, M.M.; Shemshad, M.; Biglarian, A.; Nourolahi, F.; Valad Beigi, E.; Ali Enayati, A. Species diversity and geographic distribution of hard ticks (Acari: Ixodoidea: Ixodidae) infesting domestic ruminants, in Qazvin Province, Iran. Parasitol. Res. 2012, 110, 373–380. [Google Scholar] [CrossRef]
- Provine, W.B. Ernst Mayr: Genetics and speciation. Genetics 2004, 167, 1041–1046. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Primer Name | Primer Sequence (5′–3′) | Target Gene | Product Size (bp) | Reference |
|---|---|---|---|---|---|
| Ticks | LCOI490 | GGTCAACAAATCATAAAGATATTG | cox1 | ~710 | [31] |
| HCO2198 | TAAACTTCAGGGTGACCAAAAAATCA | ||||
| Ticks | 16S+1 | CTGCTCAATGATTTTTTAAATTGCTGTGG | 16S rRNA | ~460 | [32] |
| 16S-1 | CCGGTCTGAACTCAGATCAAGT |
| Demographic Variable | Category | Study Area (Districts) | Total n (%) | ||||
|---|---|---|---|---|---|---|---|
| Charsadda n (%) | Mardan n (%) | Malakand n (%) | Peshawar n (%) | Swabi n (%) | |||
| Age (year) | Puppy (<1) | 21 (12.5) | 31 (25.2) | 24 (19.5) | 28 (22.8) | 19 (15.4) | 123 (41) |
| Juvenile (1–3) | 17 (17.2) | 23 (23.2) | 19 (19.2) | 25 (25.3) | 15 (15.2) | 99 (33) | |
| Adult (>3) | 14 (18.0) | 12 (15.4) | 19 (24.4) | 18 (23.0) | 15 (19.2) | 78 (26) | |
| Gender | Female | 38 (20.7) | 28 (15.2) | 39 (21.2) | 40 (21.7) | 39 (21.2) | 184 (61) |
| Male | 22 (19) | 23 (19.8) | 24 (20.7) | 26 (22.4) | 21 (18.1) | 116 (38.6) | |
| Breed | Short-haired | 33 (18.1) | 36 (11.5) | 38 (20.9) | 41 (22.5) | 34 (18.7) | 182 (60.6) |
| Long-haired | 23 (19.5) | 22 (18.6) | 25 (21.2) | 27 (22.9) | 21 (17.8) | 118 (39.3) | |
| Dog roaming range | Free-roaming | 41 (20.4) | 31 (15.4) | 42(20.8) | 44 (22.0) | 43 (21.4) | 201 (67.0) |
| Non-roaming | 19 (19.1) | 20 (20.2) | 20 (20.2) | 22 (22.4) | 18 (18.1) | 99 (33.0) | |
| Host | Study Area (Districts) | Total Prevalence n (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Dog | Charsadda | Mardan | Malakand | Peshawar | Swabi | ||||||
| NHE | NHTI (%) | NHE | NHTI (%) | NHE | NHTI (%) | NHE | NHTI (%) | NHE | NHTI (%) | ||
| 52 | 33 (11.0) | 66 | 44 (14.7) | 62 | 31 (10.3) | 71 | 39 (13.0) | 49 | 36 (12.0) | 183/300 (61%) | |
| Host Demographic Variable | Category | Examined Host n (%) | Tick-Infested Host n (%) | 95% CI | Chi (χ2) Statistic | p-value |
|---|---|---|---|---|---|---|
| Age (year) | Puppy (<1) | 123 (41.0) | 104 (34.7) | 29.3–40.1 | 12.210 | 0.001 |
| Juvenile (1–3) | 99 (33.0) | 54 (18.0) | 13.6–22.3 | |||
| Adult (>3) | 78 (26.0) | 25 (8.3) | 5.1–11.4 | |||
| Gender | Female | 184 (61.0) | 127 (42.3) | 36.7–47.9 | 6.115 | 0.01 |
| Male | 116 (38.6) | 56 (18.7) | 14.3–22.1 | |||
| Breed | Short-haired | 182 (60.6) | 126 (42.0) | 36.4–47.5 | 4.232 | 0.05 |
| Long-haired | 118 (39.3) | 57 (19.0) | 14.5–23.4 |
| Host Management Variable | Category | Examined Host n (%) | Tick-Infested Host n (%) | 95% CI | Chi (χ2) Statistic | p-value |
|---|---|---|---|---|---|---|
| Acaricides | No use | 168 (56.0) | 130 (43.3) | 37.7–48.9 | 17.989 | 0.001 |
| Irregular use | 85 (28.3) | 38 (12.7) | 8.9–16.5 | |||
| Regular use | 47 (15.7) | 15 (5.0) | 2.5–7.5 | |||
| Dog bathing | No | 214 (71.3) | 143 (47.7) | 42.0–53.3 | 2.391 | 0.10 |
| Yes | 86 (28.7) | 40 (13.3) | 9.5–17.1 | |||
| Previous tick infestation | Yes | 238 (79.3) | 169 (56.3) | 50.7–61.9 | 5.110 | 0.10 |
| No | 62 (20.7) | 14 (4.7) | 2.3–7.1 | |||
| Dog roaming range | Free-roaming | 201 (67.0) | 143 (47.7) | 39.1–50.3 | 6.351 | 0.01 |
| Non-roaming | 99 (33.0) | 40 (14.7) | 10.7–18.7 |
| Demographic/Host Management Associated Variable | Tick-Infested Host n (%) | Univariate Logistic Regression Analysis | Multivariate Logistic Regression Analysis | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| β | OR | (95% CI) | p-value | β | OR | (95% CI) | p-value | ||||
| LL | UL | LL | UL | ||||||||
| Age (year) | |||||||||||
| Puppy (< 1) | 104 (34.7) | 1.78 | 3.41 | 2.45 | 4.83 | 0.001 | 1.66 | 4.49 | 2.23 | 9.18 | 0.001 |
| Juvenile (1–3) | 54 (18.0) | ||||||||||
| Adult (> 3) | 25 (8.3) | ||||||||||
| Gender | |||||||||||
| Female | 127 (42.3) | 1.25 | 2.38 | 1.48 | 3.87 | 0.001 | −0.66 | 0.26 | 0.05 | 1.10 | 0.08 |
| Male | 56 (18.7) | ||||||||||
| Breed | |||||||||||
| Short-haired | 126 (42.0) | 1.43 | 2.41 | 1.49 | 3.90 | 0.001 | −0.09 | 0.08 | 0.01 | 0.48 | 0.01 |
| Long-haired | 57 (19.0) | ||||||||||
| Acaricides | |||||||||||
| No use | 130 (43.3) | 1.88 | 2.97 | 2.12 | 4.24 | 0.001 | 1.02 | 2.36 | 1.32 | 4.41 | 0.004 |
| Irregular | 38 (12.7) | ||||||||||
| Regular | 15 (5.0) | ||||||||||
| Dog bathing | |||||||||||
| Yes | 143 (47.7) | 1.14 | 2.07 | 1.24 | 3.48 | 0.005 | 0.69 | 0.90 | 0.37 | 2.13 | 0.11 |
| No | 40 (13.3) | ||||||||||
| Previous tick infestation | |||||||||||
| Yes | 169 (56.3) | 3.40 | 8.69 | 4.61 | 17.32 | 0.001 | 1.01 | 3.15 | 1.13 | 9.19 | 0.03 |
| No | 14 (4.7) | ||||||||||
| Dog roaming range | |||||||||||
| Free-roaming | 143 (47.7) | 1.92 | 3.63 | 2.21 | 6.06 | 0.001 | 1.27 | 3.7 | 0.61 | 33.6 | 0.18 |
| Non-roaming | 40 (14.7) | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeb, J.; Song, B.; Senbill, H.; Aziz, M.U.; Hussain, S.; Khan, M.A.; Qadri, I.; Cabezas-Cruz, A.; de la Fuente, J.; Sparagano, O.A. Ticks Infesting Dogs in Khyber Pakhtunkhwa, Pakistan: Detailed Epidemiological and Molecular Report. Pathogens 2023, 12, 98. https://doi.org/10.3390/pathogens12010098
Zeb J, Song B, Senbill H, Aziz MU, Hussain S, Khan MA, Qadri I, Cabezas-Cruz A, de la Fuente J, Sparagano OA. Ticks Infesting Dogs in Khyber Pakhtunkhwa, Pakistan: Detailed Epidemiological and Molecular Report. Pathogens. 2023; 12(1):98. https://doi.org/10.3390/pathogens12010098
Chicago/Turabian StyleZeb, Jehan, Baolin Song, Haytham Senbill, Muhammad Umair Aziz, Sabir Hussain, Munsif Ali Khan, Ishtiaq Qadri, Alejandro Cabezas-Cruz, José de la Fuente, and Olivier Andre Sparagano. 2023. "Ticks Infesting Dogs in Khyber Pakhtunkhwa, Pakistan: Detailed Epidemiological and Molecular Report" Pathogens 12, no. 1: 98. https://doi.org/10.3390/pathogens12010098