
Systematic Review and Modelling of Age-Dependent Prevalence of Toxoplasma gondii in Livestock, Wildlife and Felids in Europe

 , , , , , , , , , , , , ,
, , , , , , , , , , , , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
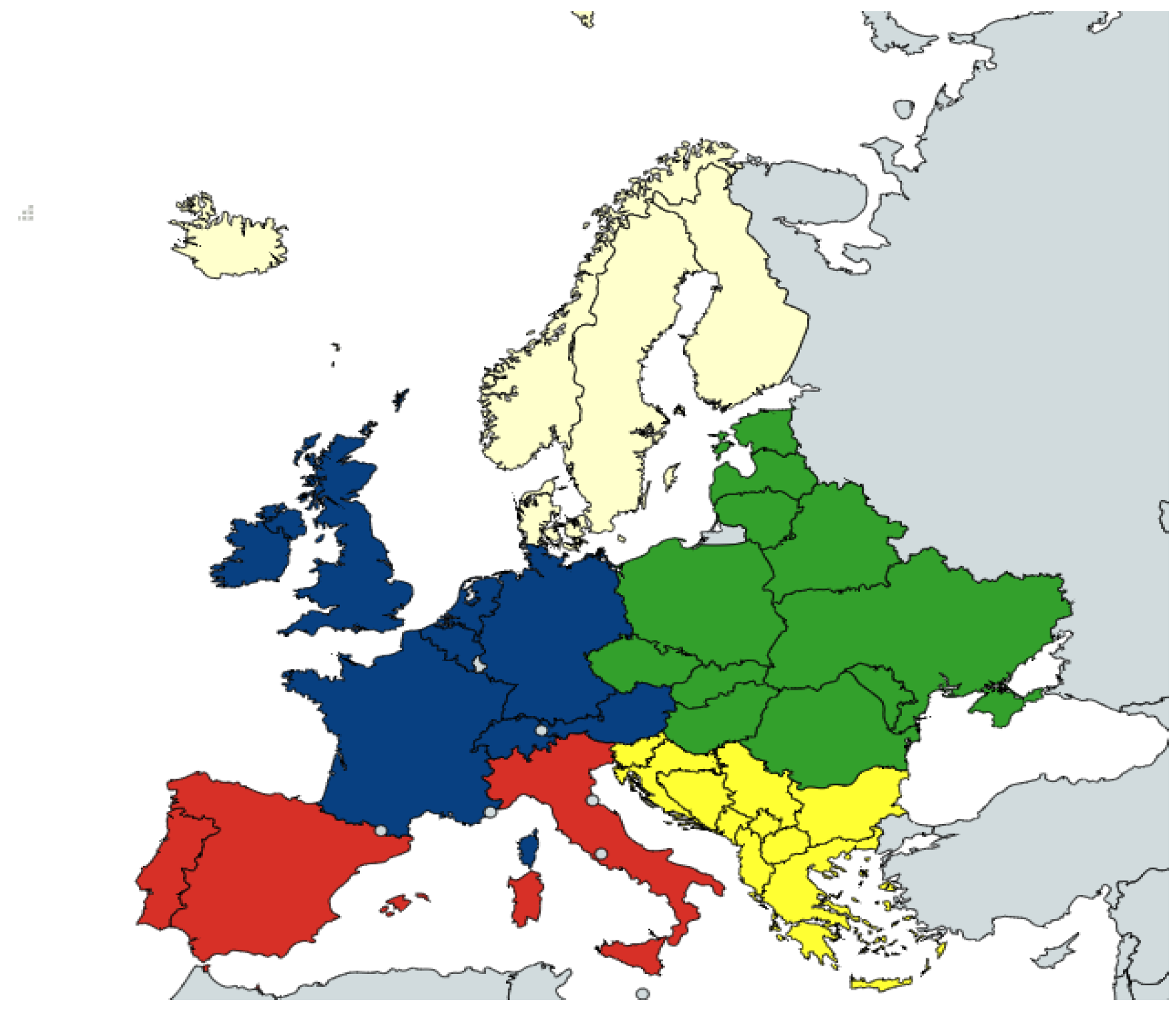
2.1. Data
2.2. Analysis of Direct Detection Data
2.3. Analysis of Indirect Detection Data
2.3.1. Age-Structured Model
2.3.2. Bayesian Hierarchical Model
3. Results
3.1. Data Collection
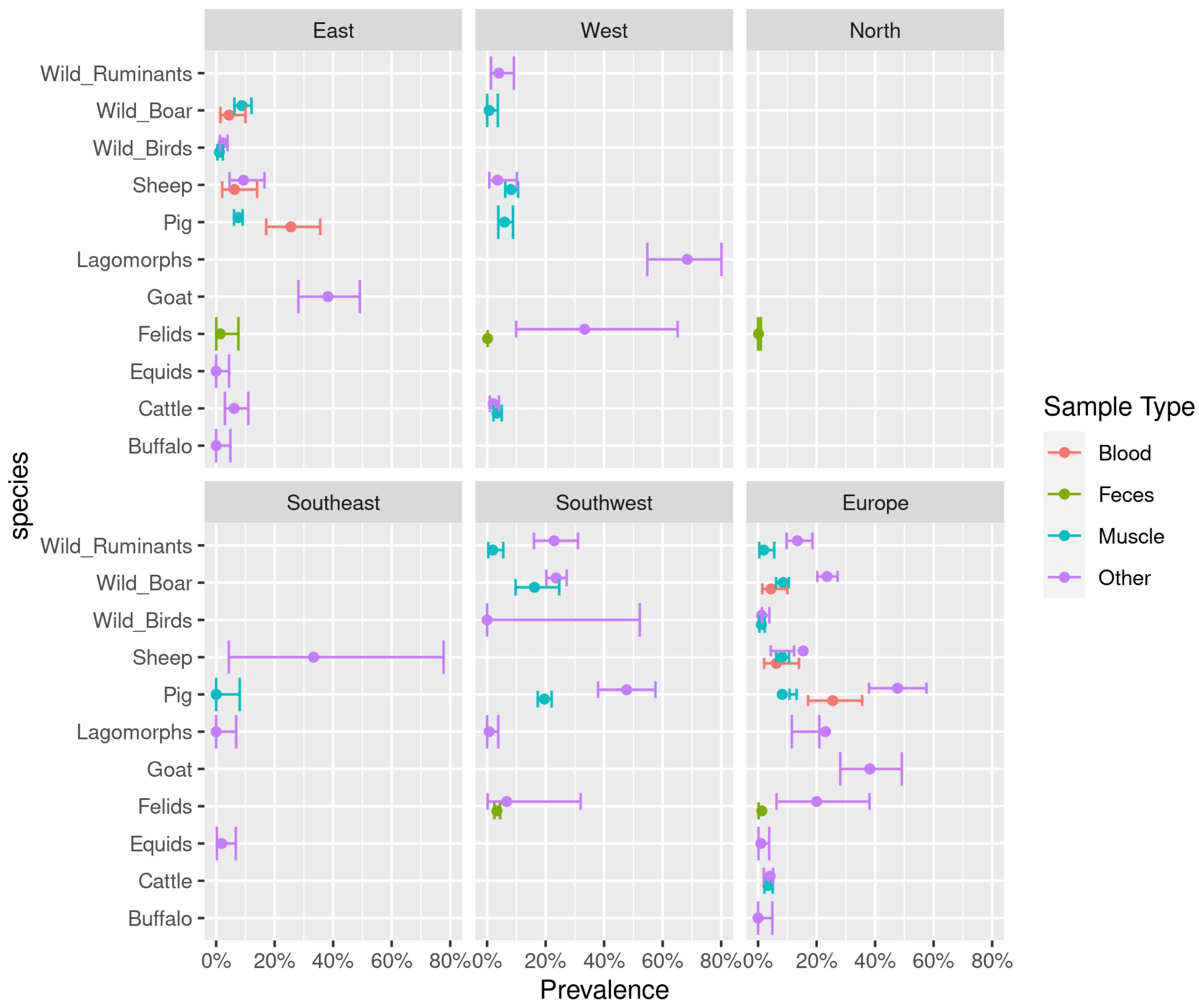
3.2. Animal Prevalence Results of Direct Methods
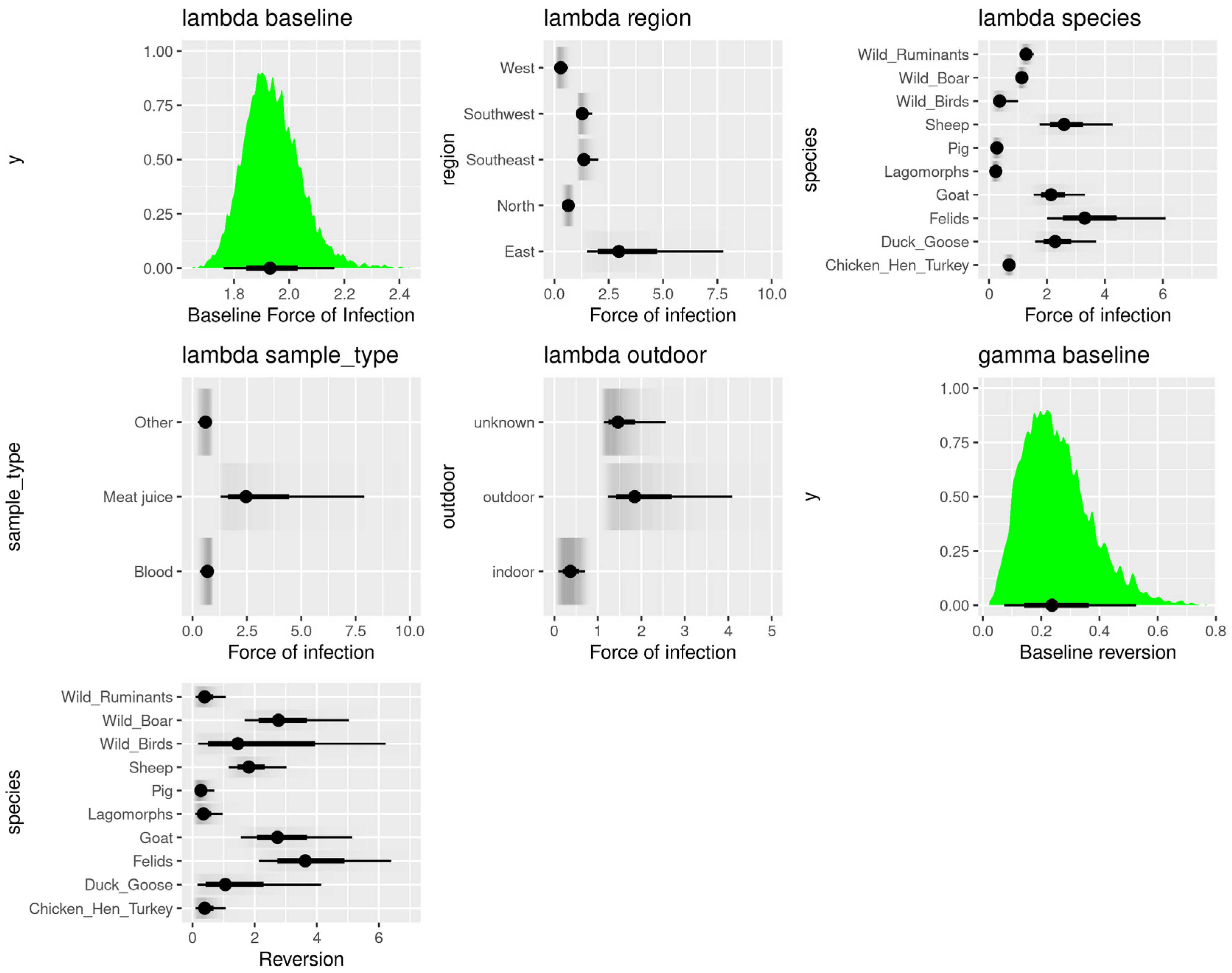
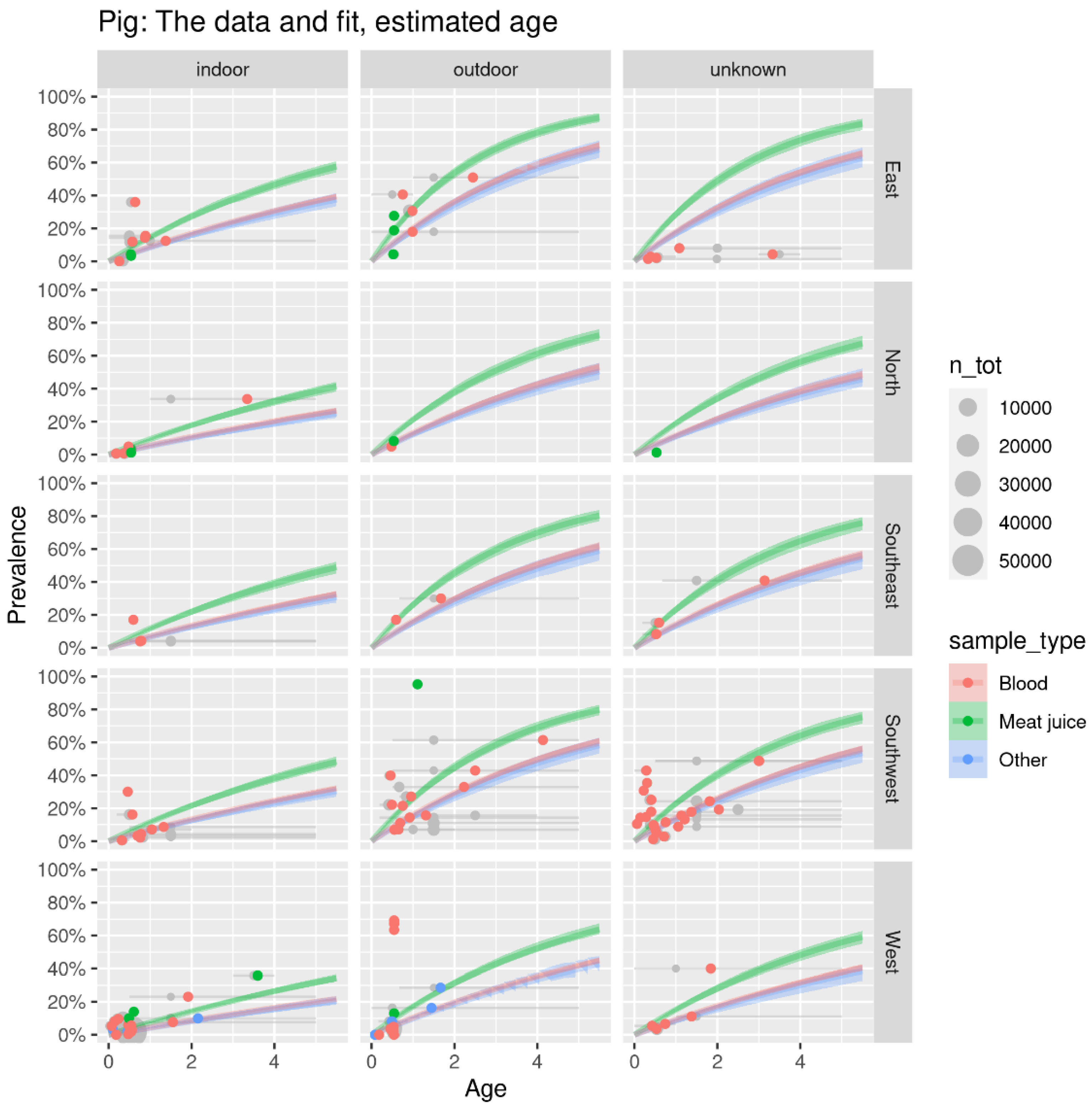
3.3. Age-Dependent Animal Seroprevalence Results Using the Bayesian Hierarchical Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P. The History of Toxoplasma gondii—The First 100 Years. J. Eukaryot. Microbiol. 2008, 55, 467–475. [Google Scholar] [CrossRef]
- Dubey, J.P. Advances in the life cycle of Toxoplasma gondii. Int. J. Parasitol. 1998, 28, 1019–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchison, W.M.; Dunachie, J.F.; Siim, J.C.; Work, K. Life cycle of Toxoplasma gondii. BMJ 1969, 4, 806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frenkel, J.K.; Dubey, J.P.; Miller, N.L. Toxoplasma gondii: Fecal forms separated from eggs of the nematode Toxocara cati. Science 1969, 164, 432–433. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, W.J.; Jeffers, V. Mechanisms of Toxoplasma gondii persistence and latency. FEMS Microbiol. Rev. 2012, 36, 717–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAuley, J.B. Congenital Toxoplasmosis. J. Pediatr. Infect. Dis. Soc. 2014, 3 (Suppl. S1), S30–S35. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ); Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; et al. Public health risks associated with food-borne parasites. EFSA J. 2018, 16, e05495. Available online: https://data.europa.eu/doi/10.2903/j.efsa.2018.5495 (accessed on 29 November 2022).
- Torgerson, P.R.; Mastroiacovo, P. The global burden of congenital toxoplasmosis: A systematic review. Bull. World Health Organ. 2013, 91, 501–508. [Google Scholar] [CrossRef]
- Cook, A.J.; Gilbert, R.E.; Buffolano, W.; Zufferey, J.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of toxoplasma infection in pregnant women: European multicentre case-control study. European Research Network on Congenital Toxoplasmosis. BMJ 2000, 321, 142–147. [Google Scholar] [CrossRef] [Green Version]
- Plant, J.W.; Richardson, N.; Moyle, G.G. Toxoplasma infection and abortion in sheep associated with feeding of grain contaminated with cat faeces. Aust. Vet. J. 1974, 50, 19–21. [Google Scholar] [CrossRef]
- Faull, W.B.; Clarkson, M.J.; Winter, A.C. Toxoplasmosis in a flock of sheep: Some investigations into its source and control. Vet. Rec. 1986, 119, 491–493. [Google Scholar] [CrossRef] [PubMed]
- Skjerve, E.; Waldeland, H.; Nesbakken, T.; Kapperud, G. Risk factors for the presence of antibodies to Toxoplasma gondii in Norwegian slaughter lambs. Prev. Vet. Med. 1998, 35, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, K.; Bahia-Oliveira, L.; Dixon, B.; Dumètre, A.; de Wit, L.A.; VanWormer, E.; Villena, I. Environmental transmission of Toxoplasma gondii: Oocysts in water, soil and food. Food Waterborne Parasitol. 2019, 15, e00049. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, S.; Basso, W.; Silván, J.B.; Ortega-Mora, L.; Maksimov, P.; Gethmann, J.; Conraths, F.; Schares, G. Toxoplasma gondii infection and toxoplasmosis in farm animals: Risk factors and economic impact. Food Waterborne Parasitol. 2019, 15, e00037. [Google Scholar] [CrossRef]
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef] [Green Version]
- Djokic, V.; Blaga, R.; Aubert, D.; Durand, B.; Perret, C.; Geers, R.; Ducry, T.; Vallee, I.; Djakovic, O.D.; Mzabi, A.; et al. Toxoplasma gondii infection in pork produced in France. Parasitology 2016, 143, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Montoya, J.G. Laboratory Diagnosis of Toxoplasma gondii Infection and Toxoplasmosis. J. Infect. Dis. 2002, 185, S73–S82. [Google Scholar] [CrossRef] [Green Version]
- Robert-Gangneux, F.; Dardé, M.L. Epidemiology of and Diagnostic Strategies for Toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef] [Green Version]
- Ybañez, R.H.D.; Ybañez, A.P.; Nishikawa, Y. Review on the Current Trends of Toxoplasmosis Serodiagnosis in Humans. Front. Cell. Infect. Microbiol. 2020, 10, 204. [Google Scholar] [CrossRef]
- Opsteegh, M. Relationship between seroprevalence in the main livestock species and presence of Toxoplasma gondii in meat (GP/EFSA/BIOHAZ/2013/01) An extensive literature review. Final report. EFSA Support. Publ. 2016, EN-996, 294. [Google Scholar] [CrossRef] [Green Version]
- Opsteegh, M.; Schares, G.; Blaga, R.; van der Giessen, J. Experimental studies on Toxoplasma gondii in the main livestock species (GP/EFSA/BIOHAZ/2013/01) Final report. EFSA Support. Publ. 2016, 13. Available online: https://data.europa.eu/doi/10.2903/sp.efsa.2016.EN-995 (accessed on 29 November 2022). [CrossRef]
- Opsteegh, M.; Spano, F.; Aubert, D.; Balea, A.; Burrells, A.; Cherchi, S.; Cornelissen, J.; Dam-Deisz, C.; Guitian, J.; Györke, A.; et al. The relationship between the presence of antibodies and direct detection of Toxoplasma gondii in slaughtered calves and cattle in four European countries. Int. J. Parasitol. 2019, 49, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- The EFSA Comprehensive European Food Consumption Database—Data Europa EU. Available online: https://data.europa.eu/data/datasets/the-efsa-comprehensive-european-food-consumption-database?locale=en (accessed on 29 November 2022).
- Kohl, C.; McIntosh, E.J.; Unger, S.; Haddaway, N.R.; Kecke, S.; Schiemann, J.; Wilhelm, R. Online tools supporting the conduct and reporting of systematic reviews and systematic maps: A case study on CADIMA and review of existing tools. Environ. Evid. 2018, 7, 8, Erratum in Environ. Evid. 2018, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Bouwknegt, M.; Devleesschauwer, B.; Graham, H.; Robertson, L.J.; van der Giessen, J.W.; The Euro-Fbp Workshop Participants. Prioritisation of food-borne parasites in Europe, 2016. Eurosurveillance 2018, 23, 17–00161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stan Development Team. Stan Modeling Language Users Guide and Reference Manual, Version 2.30. Stan-Dev.Github.Io. 2022. Available online: https://mc-stan.org/ (accessed on 29 November 2022).
- Posit. RStudio: Integrated Development for R. RStudio, PBC, Boston, MA URL. Available online: https://www.posit.co/ (accessed on 29 November 2022).
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Rubin, D.B. Bayesian Data Analysis. 1st Ed. Chapman and Hall/CRC. 1995. Available online: https://www.taylorfrancis.com/books/9781135439415 (accessed on 29 November 2022).
- Guo, M.; Mishra, A.; Buchanan, R.L.; Dubey, J.P.; Hill, D.E.; Gamble, H.R.; Jones, J.L.; Pradhan, A.K. A Systematic Meta-Analysis of Toxoplasma gondii Prevalence in Food Animals in the United States. Foodborne Pathog. Dis. 2016, 13, 109–118. [Google Scholar] [CrossRef]
- Deng, H.; Devleesschauwer, B.; Liu, M.; Li, J.; Wu, Y.; van der Giessen, J.W.B.; Opsteegh, M. Seroprevalence of Toxoplasma gondii in pregnant women and livestock in the mainland of China: A systematic review and hierarchical meta-analysis. Sci. Rep. 2018, 8, 6218. [Google Scholar] [CrossRef]
- Rostami, A.; Riahi, S.M.; Fakhri, Y.; Saber, V.; Hanifehpour, H.; Valizadeh, S.; Gholizadeh, M.; Pouya, R.H.; Gamble, H. The global seroprevalence of Toxoplasma gondii among wild boars: A systematic review and meta-analysis. Vet. Parasitol. 2017, 244, 12–20. [Google Scholar] [CrossRef]
- Foroutan, M.; Fakhri, Y.; Riahi, S.M.; Ebrahimpour, S.; Namroodi, S.; Taghipour, A.; Spotin, A.; Gamble, H.R.; Rostami, A. The global seroprevalence of Toxoplasma gondii in pigs: A systematic review and meta-analysis. Vet. Parasitol. 2019, 269, 42–52. [Google Scholar] [CrossRef]
- Montazeri, M.; Galeh, T.M.; Moosazadeh, M.; Sarvi, S.; Dodangeh, S.; Javidnia, J.; Sharif, M.; Daryani, A. The global serological prevalence of Toxoplasma gondii in felids during the last five decades (1967–2017): A systematic review and meta-analysis. Parasites Vectors 2020, 13, 82. [Google Scholar] [CrossRef]
- Hajimohammadi, B.; Ahmadian, S.; Firoozi, Z.; Askari, M.; Mohammadi, M.; Eslami, G.; Askari, V.; Loni, E.; Barzegar-Bafrouei, R.; Boozhmehrani, M.J. A Meta-Analysis of the Prevalence of Toxoplasmosis in Livestock and Poultry Worldwide. EcoHealth 2022, 19, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Belluco, S.; Mancin, M.; Conficoni, D.; Simonato, G.; Pietrobelli, M.; Ricci, A. Investigating the Determinants of Toxoplasma gondii Prevalence in Meat: A Systematic Review and Meta-Regression. PLoS ONE 2016, 11, e0153856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotteland, C.; McFerrin, B.M.; Zhao, X.; Gilot-Fromont, E.; Lélu, M. Agricultural landscape and spatial distribution of Toxoplasma gondii in rural environment: An agent-based model. Int. J. Health Geogr. 2014, 13, 45. [Google Scholar] [CrossRef] [Green Version]
- Teng, K.T.-Y.; Martinez Avilés, M.; Ugarte-Ruiz, M.; Barcena, C.; de la Torre, A.; Lopez, G.; Moreno, M.A.; Dominguez, L.; Alvarez, J. Spatial Trends in Salmonella Infection in Pigs in Spain. Front. Vet. Sci. 2020, 7, 345. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.; Berg, R.; Tagel, M.; Must, K.; Deksne, G.; Enemark, H.L.; Alban, L.; Johansen, M.V.; Nielsen, H.V.; Sandberg, M.; et al. Seroprevalence of Toxoplasma gondii in domestic pigs, sheep, cattle, wild boars, and moose in the Nordic-Baltic region: A systematic review and meta-analysis. Parasite Epidemiol. Control 2019, 5, e00100. [Google Scholar] [CrossRef] [PubMed]
- Opsteegh, M.; Swart, A.; Fonville, M.; Dekkers, L.; van der Giessen, J. Age-related Toxoplasma gondii seroprevalence in Dutch wild boar inconsistent with lifelong persistence of antibodies. PLoS ONE 2011, 6, e16240. [Google Scholar] [CrossRef]
- Opsteegh, M.; Haveman, R.; Swart, A.; Mensink-Beerepoot, M.; Hofhuis, A.; Langelaar, M.; van der Giessen, J. Seroprevalence and risk factors for Toxoplasma gondii infection in domestic cats in The Netherlands. Prev. Vet. Med. 2012, 104, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Dubey, J.P.; Beattie, C.P. Toxoplasmosis of Animals and Man; CRC Press: Boca Raton, FL, USA, 1988; 220p, ISBN 0849346185. [Google Scholar]
- Anderson, R.M.; May, R.M. The population dynamics of microparasites and their invertebrate hosts. Philos. Trans. R. Soc. B 1981, 291, 451–524. [Google Scholar]
- Restif, O.; Koella, J.C. Concurrent Evolution of Resistance and Tolerance to Pathogens. Am. Nat. 2004, 164, E90–E102. [Google Scholar] [CrossRef]
- Hutchinson, W.M.; Bradley, M.; Cheyne, W.M.; Wells, B.W.; Hay, J. Behavioural abnormalities in Toxoplasma-infected mice. Ann. Trop. Med. Parasitol. 1980, 74, 337–345. [Google Scholar] [CrossRef]
- Dubey, J.P.; Weigel, R.M.; Siegel, A.M.; Thulliez, P.; Kitron, U.D.; Mitchell, M.A.; Mannelli, A.; Mateus-Pinilla, N.; Shen, S.K.; Kwok, O.C.H.; et al. Sources and reservoirs of Toxoplasma gondii infection on 47 swine farms in Illinois. J. Parasitol. 1995, 81, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Viana, C.; Sereno, M.J.; Bersot, L.D.S.; Kich, J.D.; Nero, L.A. Comparison of Meat Juice Serology and Bacteriology for Surveillance of Salmonella in the Brazilian Pork Production Chain. Foodborne Pathog. Dis. 2020, 17, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Wallander, C.; Frössling, J.; Vågsholm, I.; Burrells, A.; Lundén, A. «Meat juice» is not a homogeneous serological matrix. Foodborne Pathog. Dis. 2015, 12, 280–288. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Animal Species Included | Testing Methods |
|---|---|---|
| Buffalo | Bubalus bubalis | Direct |
| Cattle | Bos taurus | Direct |
| Duck/Goose | Anas platyrhynchos, Anser anser, Anser cygnoides | Indirect |
| Equids | Equus caballus, Equus asinus and their cross-breeds | Direct |
| Felids | Felis catus, Felis silvestris, Lynx lynx, Lynx pardinus | Direct and Indirect |
| Goat | Capra hircus | Direct and Indirect |
| Lagomorphs | Oryctolagus cuniculus, Lepus europaeus, Lepus granatensis, Lepus timidus | Direct and Indirect |
| Poultry | Galus galus, Meleagris gallopavo | Indirect |
| Pig | Sus scrofa | Direct and Indirect |
| Sheep | Ovis aries | Direct and Indirect |
| Wild birds | Anas crecca, Aythya ferina, Anas penelope, Anas strepera, Anas platyrhynchos (feral), Anas acuta, Anas clypeata, Phasianus colchicus, Columbidae (family), Anas platyrhynchos (feral) | Direct and Indirect |
| Wild boar | Sus scrofa (feral) | Direct and Indirect |
| Wild ruminants | Rupicapra rupicapra, Cervidae (family), Dama dama, Alces alces, Ovis aries musimon, Ovis gmelini musimon, Ovis musimon, Ovis orientalis musimon, Ovis aries, Ovis ammon, Cervus elaphus, Rangifer tarandus, Rangifer tarandus platyrhynchus, Capreolus capreolus, Cervus nippon, Capra pyrenaica hispanica, Capra pyrenaica victoriae, Capra pyrenaica, Odocoileus virginianus | Direct and Indirect |
| Variable | Values |
|---|---|
| species[i] | Buffalo; Felids; Cattle; Duck, Goose; Goat; Equids; Pig; Chicken, Hen, Turkey a; Lagomorphs; Sheep; Wild Birds; Wild Boar; Wild Ruminants |
| region[i] | East, North, Southeast, Southwest, West |
| pop[i] | A unique identifier for a population |
| test[i] | Direct, Indirect |
| outdoors[i] | Outdoor, Indoor, Unknown |
| sample_type[i] | Blood, Meat juice, Other b |
| ntot[i] | Total number of animals tested |
| npos[i] | Total number of animals found positive |
| age[i] | Best estimate of average age range |
| agemin[i] | Lower bound of the age range |
| agemax[i] | Upper threshold of the age range |
| agemean[i] | The most probable age at sampling |
| Species | Holding Conditions | East | North | Southeast | Southwest | West | Europe |
|---|---|---|---|---|---|---|---|
| Chicken_Hen_Turkey | indoor | 10.3% (9.2%, 11.6%) [0.6] | 10.4% (9.2%, 11.8%) [0.9 *] | 13.0% (11.5%, 14.7%) [0.9 *] | 1.6% (1.4%, 1.8%) [0.1] | 3.4% (3.1%, 3.9%) [0.4] | 7.8% (1.5%, 14.0%) |
| outdoor | 23.8% (21.5%, 26.3%) [0.6] | 23.5% (21.1%, 26.2%) [0.9 *] | 28.8% (25.9%, 32.0%) [0.9 *] | 6.3% (5.6%, 7.0%) [0.2] | 27.4% (25.1%, 30.0%) [1.4] | 22.0% (5.9%, 30.7%) | |
| Duck_Goose | indoor | 10.2% (8.8%, 11.8%) [0.2] | 11.4% (9.7%, 13.4%) [0.4 *] | 14.3% (12.1%, 16.7%) [0.4 *] | 14.0% (11.9%, 16.2%) [0.4 *] | 5.1% (4.4%, 5.9%) [0.2] | 11.0% (4.7%, 15.9%) |
| outdoor | 25.6% (22.3%, 29.3%) [0.2] | 25.5% (21.9%, 29.6%) [0.4 *] | 31.2% (26.9%, 35.9%) [0.4 *] | 30.7% (26.7%, 35.1%) [0.4 *] | 31.0% (26.2%, 35.7%) [0.6] | 28.8% (22.8%, 35.1%) | |
| Felids | indoor | 48.0% (45.2%, 50.9%) [7.3] | 27.2% (24.7%, 29.6%) [4.6 *] | 32.9% (30.1%, 35.7%) [4.6 *] | 24.7% (22.3%, 27.1%) [3.0] | 26.5% (24.8%, 28.2%) [6.2] | 31.9% (23.3%, 49.6%) |
| outdoor | 73.8% (70.9%, 76.6%) [7.5] | 41.2% (37.6%, 44.7%) [2.9] | 49.9% (46.1%, 53.6%) [3.0] | 54.0% (50.9%, 56.9%) [3.7] | 51.5% (49.3%, 53.8%) [6.6] | 54.1% (39.1%, 75.5%) | |
| Goat | indoor | 26.5% (24.1%, 28.9%) [2.8 *] | 17.4% (15.7%, 19.1%) [2.8 *] | 21.6% (19.7%, 23.5%) [2.8 *] | 21.2% (19.2%, 23.1%) [2.8 *] | 16.0% (14.5%, 17.5%) [3.3] | 20.5% (15.1%, 27.9%) |
| outdoor | 47.5% (44.2%, 50.6%) [2.4] | 46.0% (42.9%, 49.1%) [4.0] | 44.0% (41.1%, 46.6%) [2.8] | 48.2% (45.2%, 51.0%) [3.3] | 30.6% (28.2%, 32.9%) [2.8 *] | 43.3% (29.2%, 50.2%) | |
| Lagomorphs | indoor | 2.0% (1.7%, 2.4%) [0.2] | 5.4% (4.6%, 6.3%) [1.1 *] | 6.8% (5.7%, 8.0%) [1.1 *] | 5.4% (4.6%, 6.3%) [0.9] | 4.3% (3.7%, 5.0%) [1.1 *] | 4.8% (1.8%, 7.5%) |
| outdoor | 18.8% (16.3%, 21.6%) [1.1] | 16.8% (14.4%, 19.4%) [1.5] | 20.8% (17.9%, 24.0%) [1.5] | 16.0% (13.9%, 18.3%) [1.1] | 12.1% (10.5%, 13.9%) [1.3] | 16.9% (11.1%, 22.7%) | |
| Pig | indoor | 5.2% (4.9%, 5.4%) [0.6] | 3.6% (3.3%, 3.8%) [0.6] | 8.3% (7.7%, 8.9%) [1.2] | 6.6% (6.3%, 6.9%) [1.0] | 2.4% (2.4%, 2.5%) [0.5] | 5.2% (2.4%, 8.7%) |
| outdoor | 22.0% (21.1%, 23.0%) [1.1] | 6.5% (6.0%, 7.0%) [0.5] | 16.9% (15.8%, 18.1%) [1.0] | 18.5% (17.7%, 19.3%) [1.2] | 5.5% (5.3%, 5.7%) [0.5] | 13.9% (5.4%, 22.6%) | |
| Sheep | indoor | 43.9% (41.8%, 46.1%) [3.2 *] | 30.3% (28.4%, 32.3%) [3.2 *] | 36.7% (34.5%, 38.9%) [3.2 *] | 36.1% (34.5%, 37.7%) [3.2 *] | 24.8% (23.7%, 26.0%) [3.2 *] | 34.4% (24.1%, 45.1%) |
| outdoor | 78.5% (77.0%, 79.8%) [3.7] | 61.0% (58.7%, 63.3%) [3.5] | 63.2% (61.0%, 65.4%) [2.9] | 60.0% (58.9%, 61.1%) [2.7] | 53.8% (52.5%, 55.1%) [3.6] | 63.3% (53.0%, 79.3%) | |
| Wild_Birds | indoor | 5.4% (3.2%, 9.0%) [3.8 *] | 3.4% (2.0%, 5.7%) [3.8 *] | 4.3% (2.5%, 7.1%) [3.8 *] | 4.2% (2.5%, 7.0%) [3.8 *] | 2.7% (1.6%, 4.5%) [3.8 *] | 4.0% (1.9%, 7.4%) |
| outdoor | 12.5% (7.6%, 20.3%) [3.8 *] | 8.0% (4.7%, 13.3%) [3.8 *] | 9.5% (5.7%, 15.6%) [3.5] | 9.8% (5.9%, 16.1%) [3.8] | 6.7% (4.0%, 11.1%) [4.0] | 9.3% (4.7%, 17.0%) | |
| Wild_Boar | indoor | 19.1% (17.6%, 20.7%) [2.3 *] | 12.4% (11.3%, 13.5%) [2.3 *] | 15.4% (14.0%, 16.9%) [2.3 *] | 15.1% (14.0%, 16.3%) [2.3 *] | 9.9% (9.2%, 10.6%) [2.3 *] | 14.4% (9.5%, 20.0%) |
| outdoor | 44.3% (42.1%, 46.7%) [2.8] | 28.7% (26.7%, 30.6%) [2.5] | 33.2% (30.8%, 35.6%) [2.3 *] | 33.5% (31.8%, 35.3%) [2.4] | 19.9% (18.7%, 21.1%) [2.0] | 31.9% (19.2%, 45.7%) | |
| Wild_Ruminants | indoor | 22.0% (20.3%, 23.8%) [4.8 *] | 14.3% (13.1%, 15.5%) [4.8 *] | 17.7% (16.2%, 19.5%) [4.8 *] | 17.4% (16.2%, 18.8%) [4.8 *] | 11.4% (10.6%, 12.3%) [4.8 *] | 16.6% (10.9%, 23.0%) |
| outdoor | 51.6% (49.0%, 54.3%) [5.8] | 27.7% (25.9%, 29.5%) [4.2] | 37.9% (35.3%, 40.6%) [4.8 *] | 39.1% (37.1%, 41.1%) [5.1] | 24.6% (23.2%, 26.1%) [4.6] | 36.2% (23.7%, 53.2%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dámek, F.; Swart, A.; Waap, H.; Jokelainen, P.; Le Roux, D.; Deksne, G.; Deng, H.; Schares, G.; Lundén, A.; Álvarez-García, G.; et al. Systematic Review and Modelling of Age-Dependent Prevalence of Toxoplasma gondii in Livestock, Wildlife and Felids in Europe. Pathogens 2023, 12, 97. https://doi.org/10.3390/pathogens12010097
Dámek F, Swart A, Waap H, Jokelainen P, Le Roux D, Deksne G, Deng H, Schares G, Lundén A, Álvarez-García G, et al. Systematic Review and Modelling of Age-Dependent Prevalence of Toxoplasma gondii in Livestock, Wildlife and Felids in Europe. Pathogens. 2023; 12(1):97. https://doi.org/10.3390/pathogens12010097
Chicago/Turabian StyleDámek, Filip, Arno Swart, Helga Waap, Pikka Jokelainen, Delphine Le Roux, Gunita Deksne, Huifang Deng, Gereon Schares, Anna Lundén, Gema Álvarez-García, and et al. 2023. "Systematic Review and Modelling of Age-Dependent Prevalence of Toxoplasma gondii in Livestock, Wildlife and Felids in Europe" Pathogens 12, no. 1: 97. https://doi.org/10.3390/pathogens12010097