

Role of TLRs in HIV-1 Infection and Potential of TLR Agonists in HIV-1 Vaccine Development and Treatment Strategies
 ,
,
Abstract
:1. Introduction
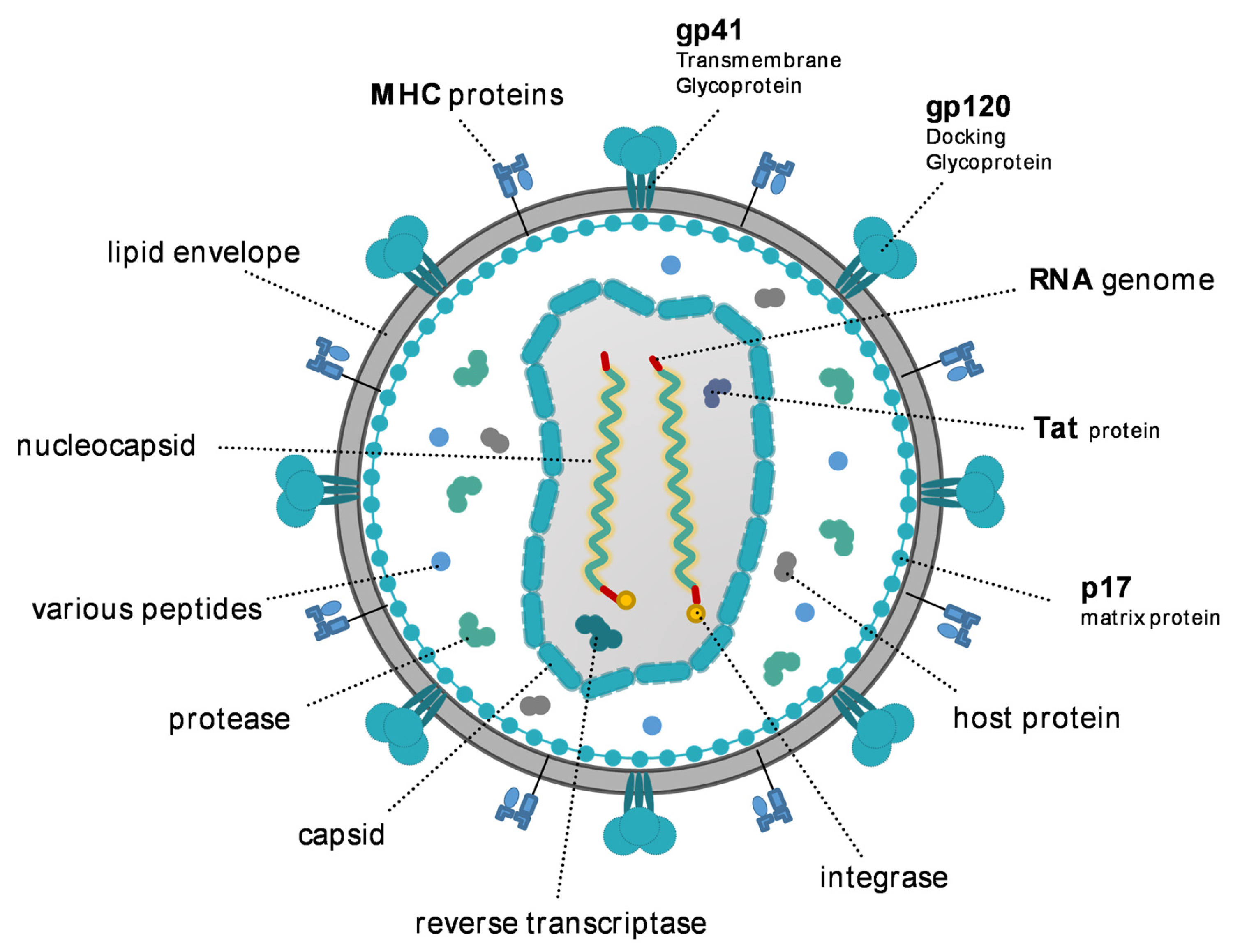
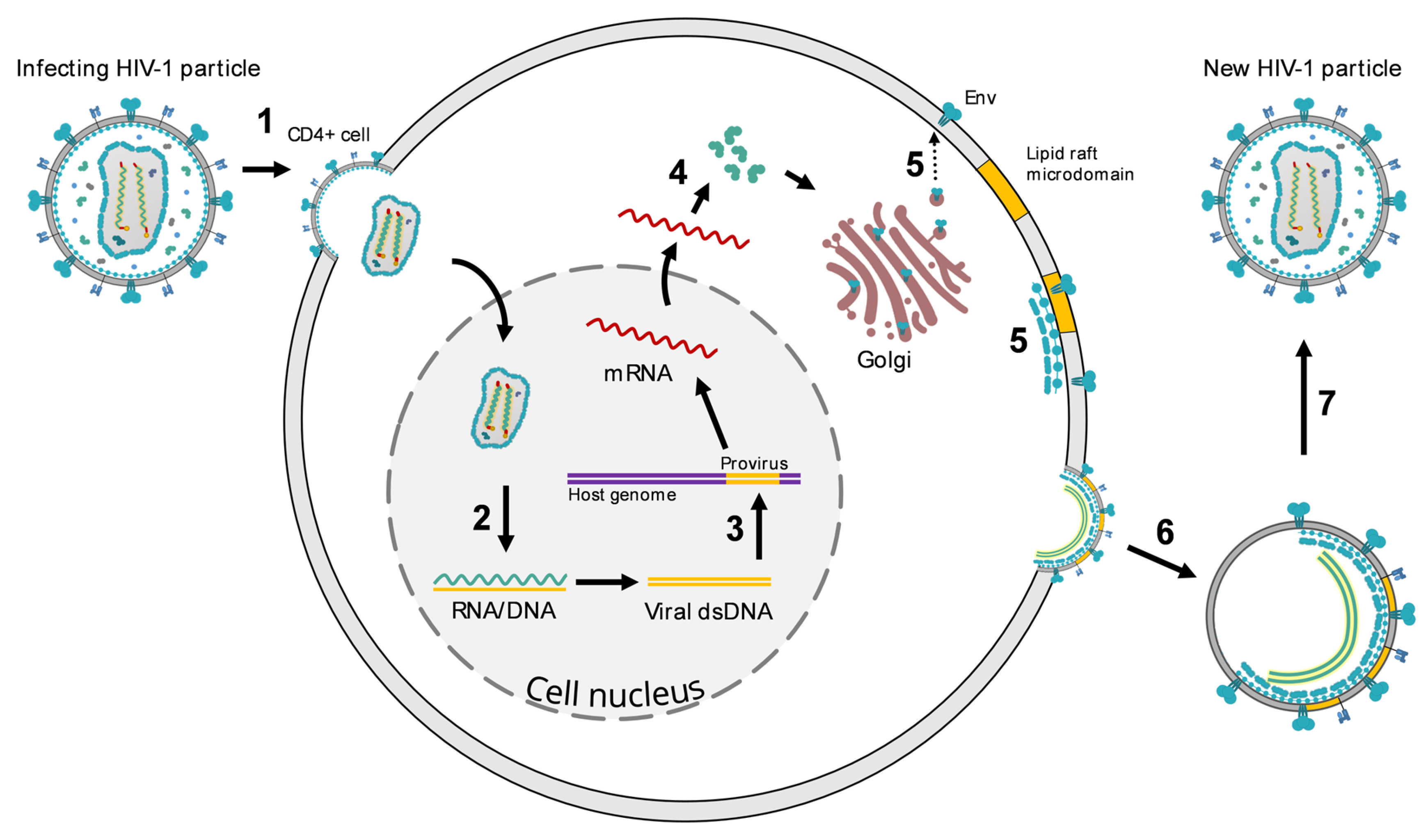
2. HIV
3. Toll-Like Receptors
3.1. Types of TLRs in Humans
3.2. Structure and Position
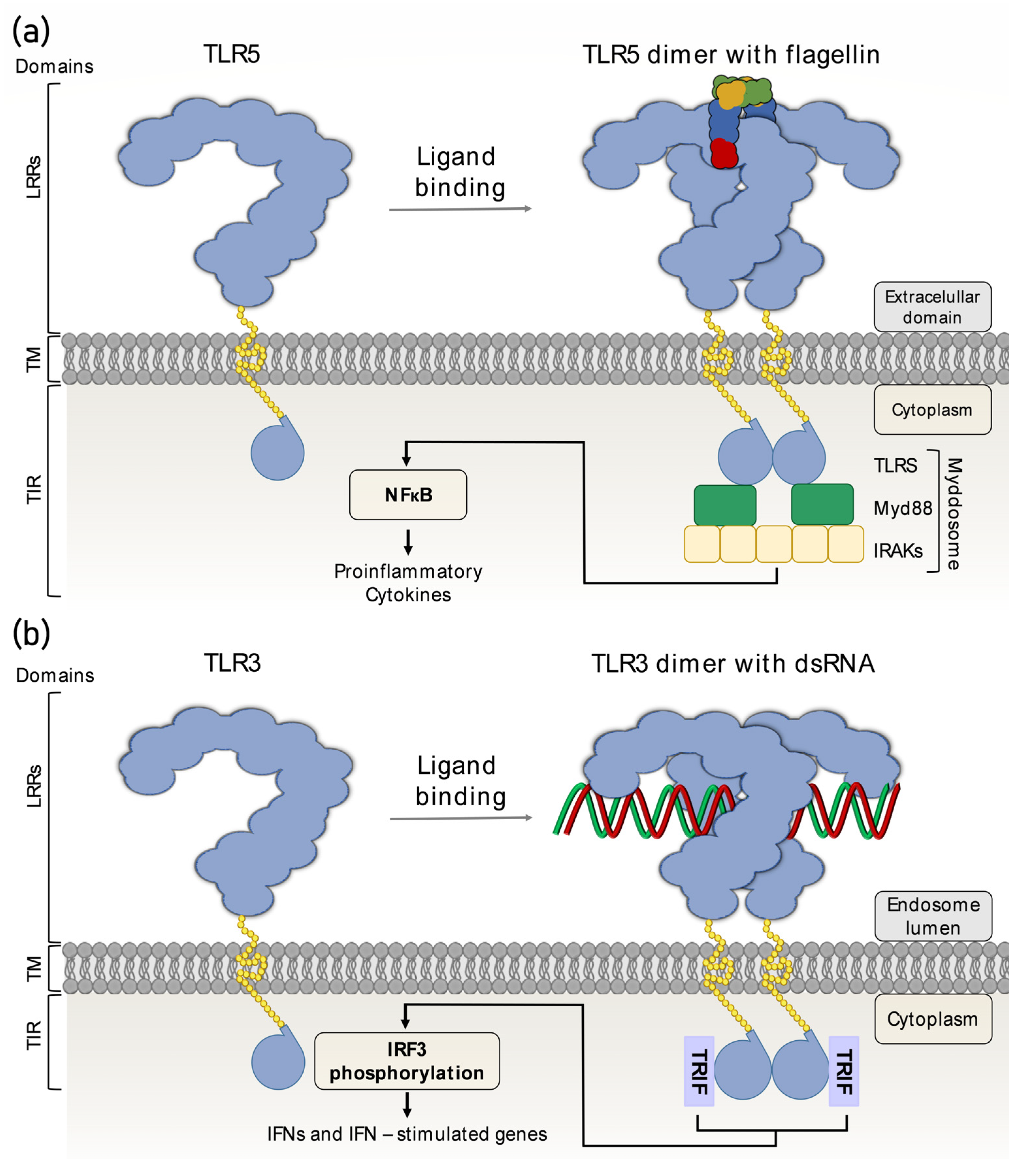
3.3. Mechanism of Action
3.4. Basic Role of TLRs in Host Immunity
Recognition of Nucleic Acids by TLRs
3.5. The Role of TLR Receptor Polymorphisms in the Biology and in the Clinical Aspects of the HIV Infection
3.5.1. TLR1 (4p14) and TLR2 (4q31.3)
3.5.2. TLR3 (4q35.1)
3.5.3. TLR4 (9q33.1)
3.5.4. TLR6 (4p14) and TLR7 (Xp22.2)
3.5.5. TLR8 (Xp22.2) and TLR9 (3p21.2)
3.5.6. Significance of TLR Polymorphisms
4. Activation of Latent HIV-1 Reservoir
TLR Agonists as Latency Reversing Agents
Studies in Non-Human Primates and Mice

{kind=link}
{kind=link}
{kind=link}
| Study | Experimental Model | Target TLR | Study Design | Main Outcomes |
|---|---|---|---|---|
| TLR agonists in humanised mice | ||||
| Cheng et al., 2018 [106] | Humanised mice (hu-mice) | TLR3 | - therapeutic vaccination of HIV-infected humanised mice with αCD40.HIV5pep vaccine (three highly conserved T-cell epitope regions of HIV Gag, Nef, and Pol fused to the C-terminus of a recombinant anti-human CD40 antibody) in combination with TLR3 agonist poly(I:C) | - poly(I:C) reactivated HIV-1 reservoirs in infected hu-mice - αCD40.HIV5pep with poly(I:C) vaccination induced HIV-specific T cell responses, reduced the concentration of proviral HIV-1 DNA in lymphoid tissues of hu-mice, and significantly delayed time to HIV-1 rebound after ART interruption |
| TLR agonists in non-human primates | ||||
| Lim et al., 2018 [100] | Macaca mulatta | TLR7 | - administration of TLR7 agonists (GS-986 and GS-9620) in 21 Indian-origin rhesus macaques that were infected with SIVmac251, treated with ART (since 65 days post infection), and experienced complete virological suppression (SIV RNA < 50 copies/mL) for about 400 days (two studies) | - TLR7 agonists induced viral replication in treated animals (up to 1000 SIV RNA copies/mL) and reduced the proviral SIV DNA reservoir (by an average of 75%) - the study failed to demonstrate a difference in time to viral rebound following ART interruption - sustained viral remission longer than 2 years in the absence of ART was observed in two animals following cessation of antiretroviral therapy |
| Del Prete et al., 2019 [101] | Macaca mulatta | TLR7 | - administration of TLR7 agonist (GS-9620) in 6 Indian-origin rhesus macaques infected with SIVmac239X, treated with ART at the early stage of infection (since day 13 post infection), and experiencing virological suppression for 75 weeks | - TLR7 agonist failed to induce a measurable increase in plasma viremia, viral RNA-to-viral DNA ratio, or decrease in viral DNA in PBMCs or tissues - SIV-specific CD8+ T-cell responses were not boosted by the TLR7 agonist |
| TLR agonists and broadly neutralizing antibodies (bNAb) in non-human primates | ||||
| Borducchi et al., 2018 [102] | Macaca mulatta | TLR7 | - a combination of TLR7 agonist (GS-9620) and V3-glycan-dependent bNAb PGT121 in 44 Indian-origin rhesus macaques infected with SIV-SF162P3 (ART initiated during acute infection) | - TLR7 agonist and bNAb combination significantly delayed viral rebound after ART discontinuation - adoptive transfer studies suggested successful targeting of latent viral reservoirs |
| Hsu et al., 2021 [75] | Macaca mulatta | TLR7 | - a combination of TLR7 agonist (GS-986) and two bNAbs (N6-LS and PGT121) in 16 Indian-origin rhesus macaques infected with SHIV-1157ipd3N4 (ART initiated at day 14 after inoculation) | - TLR agonist and bNAbs induced SHIV-specific T-cell immunity and immune activation - following ART interruption, median time to viral rebound was significantly shorter in treated vs. untreated animals (6 vs. 3 weeks, respectively) |
| Mold et al., 2022 [103] | Macaca mulatta | TLR7 | - a combination of TLR7 agonists (GS-986 and/or GS-9620) and bNAb PGT121 (either as a human IgG1, an effector enhanced IgG1, or an anti-CD3 bispecific antibody) in 33 Indian-origin rhesus macaques infected with SHIV-SF162P3 (ART initiated after 12 months of infection) | - a combination of a TLR7 agonist and all three bNAb was able to prevent viral rebound after treatment discontinuation in a subset of animals |
| Therapeutic vaccination in combination with TLR agonists in non-human primates | ||||
| Borducchi et al., 2016 [104] | Macaca mulatta | TLR7 | - therapeutic vaccination with an Ad26 vector expressing SIVsmE546 Gag/Pol/Env as prime vaccination and an MVA-based vaccine as a booster in combination with the TLR7 agonist GS-986 - study included 36 Indian-origin rhesus macaques infected with SIVmac251 | - significant reduction of median setpoint plasma SIV RNA levels and 2.5-fold delay in time to viral rebound following treatment discontinuation at week 72 - successful virological control to undetectable setpoint viral loads in 3/9 animals |
| Walker-Sperling et al., 2022 [105] | Macaca mulatta | TLR7 | - therapeutic vaccination with Ad26-based (expressing SIVsmE543 gag-pol, HIV-1 mosaic-1 env, and HIV-1 mosaic-2 env) and MVA-based (expressing SIVsmE543 gag-pol, HIV-1 mosaic-1 env-gag-pol, and HIV-1 mosaic-2 env-gag-pol) vaccines with bNAb PGT101 and TLR7 agonist GS-9620 - study included Indian-origin rhesus macaques infected with SHIV-SF162P3 (ART initiation at day 9 following infection) | - only 6/10 animals receiving combined active and passive immunisation (vaccine, bNAb, and TLR7 agonist) rebounded and three animals achieved virological control following discontinuation of ART |
| ClinicalTrials.gov Identifier | Study Coordinators | Clinical Trial | Trial Phase | Status (as of 11 November 2022) |
|---|---|---|---|---|
| TLR agonists and broadly neutralizing monoclonal antibodies | ||||
| NCT05281510 | Gilead Sciences | A Phase 2a Study to Evaluate the Safety and Tolerability of a Regimen of Dual Anti-HIV Envelope Antibodies, VRC07-523LS and CAP256V2LS, in a Sequential Regimen With a TLR Agonist, Vesatolimod, in Early Antiretroviral-Treated HIV-1 Clade C-Infected Women | Phase 2a | Recruiting |
| NCT03837756 | University of Aarhus | Combining a TLR Agonist With Broadly Neutralizing Antibodies for Reservoir Reduction and Immunological Control of HIV Infection: An Investigator-initiated Randomised, Placebo-controlled, Phase IIa Trial. | Phase 2a | Active, not yet recruiting |
| Therapeutic vaccination and TLR agonist | ||||
| NCT04364035 | Aelix Therapeutics | Phase IIa Randomised, Double-blind, Placebo-controlled Study of HIV-1 Vaccines MVA.HTI and ChAdOx1.HTI With TLR7 Agonist Vesatolimod (GS-9620) in Early Treated HIV-1 Infection | Phase 2a | Active, not yet recruiting |
| NCT04177355 | National Institute of Allergy and Infectious Diseases (NIAID) | A Phase 1 Clinical Trial to Evaluate the Safety and Immunogenicity of HIV-1 BG505 SOSIP.664 gp140 With TLR Agonist and/or Alum Adjuvants in Healthy, HIV-uninfected Adults | Phase 1 | recruiting |
| NCT04301154 | Henry M. Jackson Foundation for the Advancement of Military Medicine | Phase I, Proof of Concept, Open-Label, Randomized Clinical Trial to Evaluate the Safety and Effects of Using Prime-boost HIVIS DNA and MVA-CMDR Vaccine Regimens With or Without Toll-like Receptor 4 Agonist on HIV Reservoirs in Perinatally HIV Infected Children and Youth | Phase 1 | Recruiting |
| Active and passive immunisation with TLR agonist | ||||
| NCT04357821 | University of California, San Francisco | Combinatorial Therapy With a Therapeutic Conserved Element DNA Vaccine, MVA Vaccine Boost, TLR9 Agonist and Broadly Neutralizing Antibodies: a Proof-of-concept Study Aimed at Inducing an HIV Remission | Phase 1/phase 2 | Active, not yet recruiting |
5. Current Clinical Trials and Future Research
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- United Nations Programme on HIV/AIDS. UNAIDS Data 2021; UNAIDS: Geneva, Switzerland, 2021; pp. 4–38. [Google Scholar]
- Barré-Sinoussi, F.; Chermann, J.C.; Rey, F.; Nugeyre, M.T.; Chamaret, S.; Gruest, J.; Dauguet, C.; Axler-Blin, C.; Vézinet-Brun, F.; Rouzioux, C.; et al. Isolation of a T-Lymphotropic Retrovirus from a Patient at Risk for Acquired Immune Deficiency Syndrome (AIDS) Isolation of a T-Lymphotropic Retrovirus from a Patient at Risk for Acquired Immune Deficiency Syndrome (AIDS). Science 1983, 220, 868–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wensing, A.M.; Calvez, V.; Ceccherini, F. 2022 Update of the Drug Resistance Mutations in HIV-1. Top. Antivir. Med. 2022, 30, 559–574. [Google Scholar] [PubMed]
- Hütter, G.; Nowak, D.; Mossner, M. Long-Term Control of HIV by CCR5 Delta32/Delta32 Stem-Cell Transplantation. N. Engl. J. Med. 2009, 360, 692–698. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Peppa, D.; Hill, A.L.; Gálvez, C.; Salgado, M.; Pace, M.; McCoy, L.E.; Griffith, S.A.; Thornhill, J.; Alrubayyi, A.; et al. Evidence for HIV-1 cure after CCR5Δ32/Δ32 allogeneic haemopoietic stem-cell transplantation 30 months post analytical treatment interruption: A case report. Lancet HIV 2020, 7, e340–e347. [Google Scholar] [CrossRef] [Green Version]
- Bryson, Y.; Brown, T.R. HIV-1 remission with CCR5∆32∆32 haplo-cord transplant in a U.S. woman: IMPAACT P1107 Cases of HIV-1 Cure. In Proceedings of the CROI 2022, Virtual, 12–16 February 2022. [Google Scholar]
- Kim, Y.; Anderson, J.L.; Lewin, S.R. Getting the “Kill” into “Shock and Kill”: Strategies to Eliminate Latent HIV. Cell Host Microbe 2018, 23, 14–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoye, A.A.; Fromentin, R.; Takata, H.; Brehm, J.H.; Fukazawa, Y.; Randall, B.; Pardons, M.; Tai, V.; Tang, J.; Smedley, J.; et al. The ingenol-based protein kinase C agonist GSK445A is a potent inducer of HIV and SIV RNA transcription. PLoS Pathog. 2022, 18, e1010245. [Google Scholar] [CrossRef]
- Duan, T.; Du, Y.; Xing, C.; Wang, H.Y.; Wang, R.-F. Toll-like Receptor Signaling and Its Role in Cell-Mediated Immunity. Front. Immunol. 2022, 13, 812774. [Google Scholar] [CrossRef] [PubMed]
- Coffin, J.; Blomberg, J.; Fan, H.; Gifford, R.; Hatziioannou, T.; Lindemann, D.; Mayer, J.; Stoye, J.; Tristem, M.; Johnson, W.; et al. ICTV Virus Taxonomy Profile: Retroviridae 2021. J. Gen. Virol. 2021, 102, 001712. [Google Scholar] [CrossRef]
- Briggs, J.A.G.; Wilk, T.; Welker, R.; Kräusslich, H.-G.; Fuller, S.D. Structural organization of authentic, mature HIV-1 virions and cores. EMBO J. 2003, 22, 1707–1715. [Google Scholar] [CrossRef] [Green Version]
- PTroyano-Hernáez, P.; Reinosa, R.; Holguín, Á. HIV Capsid Protein Genetic Diversity Across HIV-1 Variants and Impact on New Capsid-Inhibitor Lenacapavir. Front. Microbiol. 2022, 13, 854974. [Google Scholar] [CrossRef]
- Watts, J.M.; Dang, K.K.; Gorelick, R.J.; Leonard, C.W.; Bess, J.W., Jr.; Swanstrom, R.; Burch, C.L.; Weeks, K.M. Architecture and secondary structure of an entire HIV-1 RNA genome. Nature 2009, 460, 711–716. [Google Scholar] [CrossRef] [Green Version]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-Length Human Immunodeficiency Virus Type 1 Genomes from Subtype C-Infected Seroconverters in India, with Evidence of Intersubtype Recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Engelman, A.N.; Kvaratskhelia, M. Multimodal Functionalities of HIV-1 Integrase. Viruses 2022, 14, 926. [Google Scholar] [CrossRef]
- Kleinpeter, A.B.; Freed, E.O. HIV-1 Maturation: Lessons Learned from Inhibitors. Viruses 2020, 12, 940. [Google Scholar] [CrossRef]
- Volchenkov, R.; Sprater, F.; Vogelsang, P.; Appel, S. The 2011 Nobel Prize in Physiology or Medicine. Scand. J. Immunol. 2012, 75, 1–4. [Google Scholar] [CrossRef]
- Vidya, M.K.; Kumar, V.G.; Sejian, V.; Bagath, M.; Krishnan, G.; Bhatta, R. Toll-like receptors: Significance, ligands, signaling pathways, and functions in mammals. Int. Rev. Immunol. 2018, 37, 20–36. [Google Scholar] [CrossRef]
- El-Zayat, S.R.; Sibaii, H.; Mannaa, F.A. Toll-like receptors activation, signaling, and targeting: An overview. Bull. Natl. Res. Cent. 2019, 43, 187. [Google Scholar] [CrossRef] [Green Version]
- Lind, N.A.; Rael, V.E.; Pestal, K.; Liu, B.; Barton, G.M. Regulation of the nucleic acid-sensing Toll-like receptors. Nat. Rev. Immunol. 2022, 22, 224–235. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like Receptor Signaling Pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Martinsen, J.T.; Gunst, J.D.; Højen, J.F.; Tolstrup, M.; Søgaard, O.S. The Use of Toll-Like Receptor Agonists in HIV-1 Cure Strategies. Front. Immunol. 2020, 11, 1112. [Google Scholar] [CrossRef]
- Shokri, M.; Tappeh, K.H.; Meshkini, E.; Aminpour, A. Evaluation of Toll-Like Receptor 11 Agonist Adjuvant Activity in Immunization of BALB/c Mice with Total Lysate Antigens of Toxoplasma gondii RH Strain. Iran. J. Parasitol. 2020, 15, 349–356. [Google Scholar] [CrossRef]
- Gonzalez, R.M.S.; Shehata, H.; O’Connell, M.J.; Yang, Y.; Moreno-Fernandez, M.E.; Chougnet, C.A.; Aliberti, J. Toxoplasma gondii-Derived Profilin Triggers Human Toll-like Receptor 5-Dependent Cytokine Production. J. Innate Immun. 2014, 6, 685–694. [Google Scholar] [CrossRef]
- Motoi, Y.; Shibata, T.; Takahashi, K.; Kanno, A.; Murakami, Y.; Li, X.; Kasahara, T.; Miyake, K. Lipopeptides are signaled by Toll-like receptor 1, 2 and 6 in endolysosomes. Int. Immunol. 2014, 26, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Sepehri, Z.; Kiani, Z.; Nasiri, A.A.; Kohan, F. Toll-like receptor 2 and type 2 diabetes. Cell Mol. Biol. Lett. 2016, 21, 2. [Google Scholar] [CrossRef] [Green Version]
- Romero, C.D.; Varma, T.K.; Hobbs, J.B.; Reyes, A.; Driver, B.; Sherwood, E.R. The Toll-like Receptor 4 Agonist Monophosphoryl Lipid A Augments Innate Host Resistance to Systemic Bacterial Infection. Infect. Immun. 2011, 79, 3576–3587. [Google Scholar] [CrossRef] [Green Version]
- Hajam, I.A.; Dar, P.; Shahnawaz, I.; Jaume, J.C.; Lee, J.H. Bacterial flagellin—A potent immunomodulatory agent. Exp. Mol. Med. 2017, 49, e373. [Google Scholar] [CrossRef] [Green Version]
- Bagheri-Hosseinabadi, Z.; Zarandi, E.R.; Mirabzadeh, M.; Amiri, A.; Abbasifard, M. mRNA expression of toll-like receptors 3, 7, 8, and 9 in the nasopharyngeal epithelial cells of coronavirus disease 2019 patients. BMC Infect. Dis. 2022, 22, 448. [Google Scholar] [CrossRef]
- Henrick, B.M.; Yao, X.-D.; Zahoor, M.; Abimiku, A.; Osawe, S.; Rosenthal, K.L. TLR10 Senses HIV-1 Proteins and Significantly Enhances HIV-1 Infection. Front. Immunol. 2019, 10, 482. [Google Scholar] [CrossRef]
- Oosting, M.; Cheng, S.-C.; Bolscher, J.M.; Vestering-Stenger, R.; Plantinga, T.S.; Verschueren, I.C.; Arts, P.; Garritsen, A.; van Eenennaam, H.; Sturm, P.; et al. Human TLR10 is an anti-inflammatory pattern-recognition receptor. Proc. Natl. Acad. Sci. USA 2014, 111, E4478–E4484. [Google Scholar] [CrossRef] [Green Version]
- Sousa, C.; Fernandes, S.A.; Cardoso, J.C.R.; Wang, Y.; Zhai, W.; Guerreiro, P.M.; Chen, L.; Canário, A.V.M.; Power, D.M. Toll-like Receptor Evolution: Does Temperature Matter? Front. Immunol. 2022, 13, 812890. [Google Scholar] [CrossRef]
- Asami, J.; Shimizu, T. Structural and functional understanding of the toll-like receptors. Protein Sci. 2021, 30, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Tanji, H.; Ohto, U.; Motoi, Y.; Shibata, T.; Miyake, K.; Shimizu, T. Autoinhibition and relief mechanism by the proteolytic processing of Toll-like receptor 8. Proc. Natl. Acad. Sci. USA 2016, 113, 3012–3017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, K.; Akira, S. TLR7 Structure: Cut in Z-Loop. Immunity 2016, 45, 705–707. [Google Scholar] [CrossRef] [Green Version]
- Sameer, A.S.; Nissar, S. Toll-like Receptors (TLRs): Structure, Functions, Signaling, and Role of Their Polymorphisms in Colorectal Cancer Susceptibility. BioMed. Res. Int. 2021, 2021, 1157023. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Su, L.; Morin, M.D.; Jones, B.T.; Whitby, L.R.; Surakattula, M.M.R.P.; Huang, H.; Shi, H.; Choi, J.H.; Wang, K.-W.; et al. TLR4/MD-2 activation by a synthetic agonist with no similarity to LPS. Proc. Natl. Acad. Sci. USA 2016, 113, E884–E893. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Chen, J.; Chu, F.; Zhu, J.; Jin, T. Inflammatory Role of TLR-MyD88 Signaling in Multiple Sclerosis. Front. Mol. Neurosci. 2020, 12, 314. [Google Scholar] [CrossRef]
- Han, K.-J.; Su, X.; Xu, L.-G.; Bin, L.-H.; Zhang, J.; Shu, H.-B. Mechanisms of the TRIF-induced Interferon-stimulated Response Element and NF-κB Activation and Apoptosis Pathways. J. Biol. Chem. 2004, 279, 15652–15661. [Google Scholar] [CrossRef] [Green Version]
- Nouri, Y.; Weinkove, R.; Perret, R. T-cell intrinsic Toll-like receptor signaling: Implications for cancer immunotherapy and CAR T-cells. J. Immunother. Cancer 2021, 9, e003065. [Google Scholar] [CrossRef]
- Dalod, M.; Chelbi, R.; Malissen, B.; Lawrence, T. Dendritic cell maturation: Functional specialization through signaling specificity and transcriptional programming. EMBO J. 2014, 33, 1104–1116. [Google Scholar] [CrossRef] [Green Version]
- López, C.B.; Moltedo, B.; Alexopoulou, L.; Bonifaz, L.; Flavell, R.A.; Moran, T.M. TLR-Independent Induction of Dendritic Cell Maturation and Adaptive Immunity by Negative-Strand RNA Viruses. J. Immunol. 2022, 173, 6882–6889. [Google Scholar] [CrossRef]
- Komai-Koma, M.; Jones, L.; Ogg, G.S.; Xu, D.; Liew, F.Y. TLR2 is expressed on activated T cells as a costimulatory receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 3029–3034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchta, C.M.; Bishop, G.A. Toll-like receptors and B cells: Functions and mechanisms. Immunol. Res. 2014, 59, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Hou, B. TLR signaling in B-cell development and activation. Cell Mol. Immunol. 2013, 10, 103–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- YLu, Y.; Li, Z.; Ma, C.; Wang, H.; Zheng, J.; Cui, L.; He, W. The interaction of influenza H5N1 viral hemagglutinin with sialic acid receptors leads to the activation of human γδ T cells. Cell Mol. Immunol. 2013, 10, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.D.; Heeke, D.S.; Rao, E.; Maynard, S.K.; Hornigold, D.; McCrae, C.; Fraser, N.; Tovchigrechko, A.; Yu, L.; Williams, N.; et al. A Novel Class of Small Molecule Agonists with Preference for Human over Mouse TLR4 Activation. PLoS ONE 2016, 11, e0164632. [Google Scholar] [CrossRef] [Green Version]
- Funchal, G.A.; Jaeger, N.; Czepielewski, R.S.; Machado, M.S.; Muraro, S.P.; Stein, R.T.; Bonorino, C.B.C.; Porto, B.N. Respiratory Syncytial Virus Fusion Protein Promotes TLR-4–Dependent Neutrophil Extracellular Trap Formation by Human Neutrophils. PLoS ONE 2015, 10, e0124082. [Google Scholar] [CrossRef] [Green Version]
- Klammer, M.G.; Dzaye, O.; Wallach, T.; Krüger, C.; Gaessler, D.; Buonfiglioli, A.; Derkow, K.; Kettenmann, H.; Brinkmann, M.M.; Lehnardt, S. UNC93B1 Is Widely Expressed in the Murine CNS and Is Required for Neuroinflammation and Neuronal Injury Induced by MicroRNA let-7b. Front. Immunol. 2021, 12, 715774. [Google Scholar] [CrossRef]
- Pelka, K.; Bertheloot, D.; Reimer, E.; Phulphagar, K.; Schmidt, S.V.; Christ, A.; Stahl, R.; Watson, N.; Miyake, K.; Hacohen, N.; et al. The Chaperone UNC93B1 Regulates Toll-like Receptor Stability Independently of Endosomal TLR Transport. Immunity 2018, 48, 911–922.e7. [Google Scholar] [CrossRef]
- Miyake, K.; Shibata, T.; Fukui, R.; Sato, R.; Saitoh, S.-I.; Murakami, Y. Nucleic Acid Sensing by Toll-Like Receptors in the Endosomal Compartment. Front. Immunol. 2022, 13, 941931. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, J.; Zhao, Y.; Ma, X.; Yi, H. Toll-like receptor 3 (TLR3) regulation mechanisms and roles in antiviral innate immune responses. J. Zhejiang Univ. B 2021, 22, 609–632. [Google Scholar] [CrossRef]
- Okude, H.; Ori, D.; Kawai, T. Signaling Through Nucleic Acid Sensors and Their Roles in Inflammatory Diseases. Front. Immunol. 2021, 11, 625833. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; He, H.; Sun, C.; Fu, J.; Ghosh, D.; Deng, C.; Sheng, Y. Association of toll-like receptor polymorphisms with acquisition of HIV infection and clinical findings. Medicine 2020, 99, e23663. [Google Scholar] [CrossRef] [PubMed]
- Willie, B.M.; Hall, N.B.; Stein, C.M.; Jurevic, R.J.; Weinberg, A.M.; Mehlotra, R.K.; A Zimmerman, P. Association of Toll-like receptor polymorphisms with HIV status in North Americans. Genes Immun. 2014, 15, 569–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidyant, S.; Chatterjee, A.; Agarwal, V.; Dhole, T.N. Susceptibility to HIV-1 infection is influenced by toll like receptor-2 (−196 to −174) polymorphism in a north Indian population. J. Gene Med. 2017, 19, e2971. [Google Scholar] [CrossRef]
- Royo, J.L.; Alarcón-Martín, E.; Díaz-Fuentes, J.; Colmenero, J.D.; Bravo, M.J. Discordance in TLR2 (−196 to −174) polymorphism effect on HIV infection risk. J. Gene Med. 2018, 20, e3051. [Google Scholar] [CrossRef]
- Laplana, M.; Bravo, M.J.; Fernández-Fuertes, M.; Ruiz-Garcia, C.; Alarcón-Martin, E.; Colmenero, J.D.D.; Caruz, A.; Fibla, J.; Real, L.M.; Royo, J.L. Toll-Like Receptor 2 Promoter −196 to −174 Deletion Affects CD4 Levels Along Human Immunodeficiency Virus Infection Progression. J. Infect. Dis. 2020, 222, 2007–2011. [Google Scholar] [CrossRef]
- Kaushik, G.; Vashishtha, R.; Tripathi, H.; Yadav, R.N. Genetic Polymorphism of Toll-like Receptors in HIV-I Infected Patients with and without Tuberculosis Co-Infection. Int. J. Mycobacteriol. 2022, 6, 239–245. [Google Scholar] [CrossRef]
- Onyishi, C.U.; May, R.C. Human immune polymorphisms associated with the risk of cryptococcal disease. Immunology 2022, 165, 143–157. [Google Scholar] [CrossRef]
- Huik, K.; Avi, R.; Pauskar, M.; Kallas, E.; Jõgeda, E.-L.; Karki, T.; Marsh, K.; Jarlais, D.D.; Uusküla, A.; Lutsar, I. Association between TLR3 rs3775291 and resistance to HIV among highly exposed Caucasian intravenous drug users. Infect. Genet. Evol. 2013, 20, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Vidyant, S.; Chatterjee, A.; Dhole, T. A single-nucleotide polymorphism in TLR4 is linked with the risk of HIV-1 infection. Br. J. Biomed. Sci. 2018, 76, 59–63. [Google Scholar] [CrossRef]
- Habibabadi, H.M.; Parsania, M.; Pourfathollah, A.A.; Haghighat, S.; Sharifi, Z. Association of TLR3 single nucleotide polymorphisms with susceptibility to HTLV-1 infection in Iranian asymptomatic blood donors. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200026. [Google Scholar] [CrossRef]
- Kim, Y.-C.; Jeong, B.-H. Strong Association of the rs4986790 Single Nucleotide Polymorphism (SNP) of the Toll-like Receptor 4 (TLR4) Gene with Human Immunodeficiency Virus (HIV) Infection: A Meta-Analysis. Genes 2021, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Mungmunpuntipantip, R.; Wiwanitkit, V. Genetic polymorphism of toll-like receptors in HIV-I infected patient and tuberculosis Co-Infection. Int. J. Mycobacteriol. 2022, 11, 225. [Google Scholar] [CrossRef] [PubMed]
- Jabłońska, A.; Studzińska, M.; Szenborn, L.; Wiśniewska-Ligier, M.; Karlikowska-Skwarnik, M.; Gęsicki, T.; Paradowska, E. TLR4 896A/G and TLR9 1174G/A polymorphisms are associated with the risk of infectious mononucleosis. Sci. Rep. 2020, 10, 13154. [Google Scholar] [CrossRef] [PubMed]
- Tarancon-Diez, L.; de Pablo-Bernal, R.S.; Jiménez, J.L.; Álvarez-Ríos, A.I.; Genebat, M.; Rosado-Sánchez, I.; Muñoz-Fernández, M.; Ruiz-Mateos, E.; Leal, M. Role of toll-like receptor 4 Asp299Gly polymorphism in the development of cardiovascular diseases in HIV-infected patients. AIDS 2018, 32, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Said, E.; Al-Yafei, F.; Zadjali, F.; Hasson, S.; Al-Balushi, M.; Al-Mahruqi, S.; Koh, C.; Al-Naamani, K.; Al-Busaidi, J.; Idris, M.; et al. Association of single-nucleotide polymorphisms in TLR7 (Gln11Leu) and TLR9 (1635A/G) with a higher CD4T cell count during HIV infection. Immunol. Lett. 2014, 160, 58–64. [Google Scholar] [CrossRef]
- Anokhin, V.V.; Bakhteeva, L.B.; Khasanova, G.R.; Khaiboullina, S.F.; Martynova, E.V.; Tillett, R.L.; Schlauch, K.A.; Lombardi, V.C.; Rizvanov, A.A. Previously Unidentified Single Nucleotide Polymorphisms in HIV/AIDS Cases Associate with Clinical Parameters and Disease Progression. BioMed. Res. Int. 2016, 2016, 2742648. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhu, J.; Su, B.; Cao, L.; Li, Z.; Wei, H.; Huang, X.; Zheng, K.; Li, A.; Chen, N.; et al. Effects of TLR7 Polymorphisms on the Susceptibility and Progression of HIV-1 Infection in Chinese MSM Population. Front. Immunol. 2020, 11, 589010. [Google Scholar] [CrossRef]
- Shaikh, N.N.; Nirmalkar, A.; Thakar, M. Polymorphisms in Toll-Like Receptors (TLRs)-7 and 9 Genes in Indian Population with Progressive and Nonprogressive HIV-1 Infection. AIDS Res. Hum. Retrovir. 2019, 35, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.-Y.; Baumann, K.; Hamouda, O.; Eckert, J.K.; Neumann, K.; Kücherer, C.; Bartmeyer, B.; Poggensee, G.; Oh, N.; Pruss, A.; et al. A frequent functional toll-like receptor 7 polymorphism is associated with accelerated HIV-1 disease progression. AIDS 2009, 23, 297–307. [Google Scholar] [CrossRef]
- Singh, H.; Samani, D.; Aggarwal, S. TLR7 Polymorphism (rs179008 and rs179009) in HIV-Infected Individual Naïve to ART. Mediat. Inflamm. 2020, 2020, 6702169. [Google Scholar] [CrossRef] [PubMed]
- Beima-Sofie, K.M.; Bighma, A.W.; Lingappa, J.; Wamalwa, D.; Mackelprang, R.D.; Bamshad, M.J.; Maleche-Obimbo, E.; Richardson, B.A.; John-Stewart, G.C. Toll-like Receptor (TLR) variants are associated with infant HIV-1 acquisition and peak plasma HIV-1 RNA level. AIDS 2013, 23, 2431–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, D.C.; Schuetz, A.; Imerbsin, R.; Silsorn, D.; Pegu, A.; Inthawong, D.; Sopanaporn, J.; Visudhiphan, P.; Chuenarom, W.; Keawboon, B.; et al. TLR7 agonist, N6-LS and PGT121 delayed viral rebound in SHIV-infected macaques after antiretroviral therapy interruption. PLoS Pathog. 2021, 17, e1009339. [Google Scholar] [CrossRef]
- Joshi, A.; Punke, E.B.; Mehmetoglu-Gurbuz, T.; Peralta, D.P.; Garg, H. TLR9 polymorphism correlates with immune activation, CD4 decline and plasma IP10 levels in HIV patients. BMC Infect. Dis. 2019, 19, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallejo, A.; Molina-Pinelo, S.; de Felipe, B.; Abad-Fernández, M.; González-Escribano, M.F.; Leal, M.; Soriano-Sarabia, N. Brief Report: Toll-like Receptor 9-1635A/G Polymorphism Is Associated with HIV-1 Rebound after Four Weeks of Interruption of Antiretroviral Therapy. J. Acquir. Immune Defic. Syndr. 2020, 85, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Bochud, P.-Y.; Hersberger, M.; Taffé, P.; Bochud, M.; Stein, C.; Rodrigues, S.D.; Calandra, T.; Francioli, P.; Telenti, A.; Speck, R.; et al. Polymorphisms in Toll-like receptor 9 influence the clinical course of HIV-1 infection. AIDS 2007, 21, 441–446. [Google Scholar] [CrossRef]
- Kulmann-Leal, B.; Ellwanger, J.H.; Valverde-Villegas, J.M.; Simon, D.; Marangon, C.G.; Mattevi, V.S.; Lazzaretti, R.K.; Kuhmmer, R.; Sprinz, E.; Chies, J.A.B. TLR9 2848 G/A Gene Polymorphism in HCV+, HIV+, and HCV+/HIV+ Individuals. Genet. Test. Mol. Biomark. 2022, 26, 263–269. [Google Scholar] [CrossRef]
- Valverde-Villegas, J.M.; dos Santos, B.P.; de Medeiros, R.M.; Mattevi, V.S.; Lazzaretti, R.K.; Sprinz, E.; Kuhmmer, R.; Chies, J.A.B. Endosomal toll-like receptor gene polymorphisms and susceptibility to HIV and HCV co-infection—Differential influence in individuals with distinct ethnic background. Hum. Immunol. 2017, 78, 221–226. [Google Scholar] [CrossRef]
- Jabłońska, A.; Jabłonowska, E.; Studzińska, M.; Kamerys, J.; Paradowska, E. The TLR9 2848C/T Polymorphism Is Associated with the CMV DNAemia among HIV/CMV Co-Infected Patients. Cells 2021, 10, 2360. [Google Scholar] [CrossRef]
- Varshney, D.; Singh, S.; Sinha, E.; Mohanty, K.K.; Kumar, S.; Barik, S.K.; Patil, S.A.; Katara, P. Systematic review and meta-analysis of human Toll-like receptors genetic polymorphisms for susceptibility to tuberculosis infection. Cytokine 2022, 152, 155791. [Google Scholar] [CrossRef]
- Washizaki, A.; Murata, M.; Seki, Y.; Kikumori, M.; Tang, Y.; Tan, W.; Wardani, N.P.; Irie, K.; Akari, H. The Novel PKC Activator 10-Methyl-Aplog-1 Combined with JQ1 Induced Strong and Synergistic HIV Reactivation with Tolerable Global T Cell Activation. Viruses 2021, 13, 2037. [Google Scholar] [CrossRef] [PubMed]
- Elsheikh, M.M.; Tang, Y.; Li, D.; Jiang, G. Deep latency: A new insight into a functional HIV cure. eBioMedicine 2019, 45, 624–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- French, A.J.; Natesampillai, S.; Krogman, A.; Correia, C.; Peterson, K.L.; Alto, A.; Chandrasekar, A.P.; Misra, A.; Li, Y.; Kaufmann, S.H.; et al. Reactivating latent HIV with PKC agonists induces resistance to apoptosis and is associated with phosphorylation and activation of BCL2. PLoS Pathog. 2020, 16, e1008906. [Google Scholar] [CrossRef]
- Eckschlager, T.; Plch, J.; Stiborova, M.; Hrabeta, J. Histone Deacetylase Inhibitors as Anticancer Drugs. Int. J. Mol. Sci. 2017, 18, 1414. [Google Scholar] [CrossRef] [Green Version]
- Ait-Ammar, A.; Kula, A.; Darcis, G.; Verdikt, R.; De Wit, S.; Gautier, V.; Mallon, P.W.G.; Marcello, A.; Rohr, O.; Van Lint, C. Current Status of Latency Reversing Agents Facing the Heterogeneity of HIV-1 Cellular and Tissue Reservoirs. Front. Microbiol. 2020, 10, 3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.K.L.; Greene, W.C. NF-kB roles in latency. Curr. Opin. HIV AIDS 2012, 6, 12–18. [Google Scholar] [CrossRef]
- Bosque, A.; Nilson, K.A.; Macedo, A.B.; Spivak, A.M.; Archin, N.M.; Van Wagoner, R.M.; Martins, L.J.; Novis, C.L.; Szaniawski, M.A.; Ireland, C.M. HHS Public Access. Cell Rep. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Krämer, O.H.; Moriggl, R. Acetylation and sumoylation control STAT5 activation antagonistically. JAK-STAT 2012, 1, 203–207. [Google Scholar] [CrossRef] [Green Version]
- Vrendenbarg, S. The Role of TLR7 and/or TLR8 Agonists in HIV Cure. Master’s Thesis, Utrecht University, Utrecht, The Netherlands, 2021. [Google Scholar]
- Macedo, A.B.; Novis, C.L.; Bosque, A. Targeting Cellular and Tissue HIV Reservoirs with Toll-like Receptor Agonists. Front. Immunol. 2019, 10, 2450. [Google Scholar] [CrossRef] [Green Version]
- Lisziewicz, J.; Sun, D.; Weichold, F.F.; Thierry, A.R.; Lusso, P.; Tang, J.; Gallo, R.C.; Agrawal, S. Antisense oligodeoxynucleotide phosphorothioate complementary to Gag mRNA blocks replication of human immunodeficiency virus virus type 1 in human peripheral blood cells. Proc. Natl. Acad. Sci. USA 1994, 91, 7942–7946. [Google Scholar] [CrossRef]
- Duan, S.; Xu, X.; Wang, J.; Huang, L.; Peng, J.; Yu, T.; Zhou, Y.; Cheng, K.; Liu, S. TLR1/2 Agonist Enhances Reversal of HIV-1 Latency and Promotes NK Cell-Induced Suppression of HIV-1-Infected Autologous CD4+ T Cells. J. Virol. 2021, 95, JVI0081621. [Google Scholar] [CrossRef] [PubMed]
- Tsai, A.; Irrinki, A.; Kaur, J.; Cihlar, T.; Kukolj, G.; Sloan, D.D.; Murry, J.P. Toll-like receptor 7 agonist GS-9620 induces HIV expression and HIV-specific immunity in cells from HIV-infected individuals on suppressive antiretroviral therapy. J. Virol. 2017, 91, e02166-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanford, R.E.; Guerra, B.; Chavez, D.; Giavedoni, L.; Hodara, V.L.; Brasky, K.M.; Fosdick, A.; Frey, C.R.; Zheng, J.; Wolfgang, G.; et al. GS-9620, an Oral Agonist of Toll-like Receptor-7, Induces Prolonged Suppression of Hepatitis B Virus in Chronically Infected Chimpanzees. Gastroenterology 2013, 144, 1508–1517.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlaepfer, E.; Speck, R.F. TLR8 Activates HIV from Latently Infected Cells of Myeloid-Monocytic Origin Directly via the MAPK Pathway and from Latently Infected CD4+ T Cells Indirectly via TNF-α. J. Immunol. 2016, 186, 4314–4324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Offersen, R.; Nissen, S.K.; Rasmussen, T.A.; Østergaard, L.; Denton, P.W.; Søgaard, O.S.; Tolstrup, M. Transcription and NK Cell-Mediated Inhibition of HIV-1-Infected. J. Virol. 2016, 90, 4441–4453. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Wang, Q.; Li, G.; Banga, R.; Ma, J.; Yu, H.; Yasui, F.; Zhang, Z.; Pantaleo, G.; Perreau, M.; et al. TLR3 agonist and CD40-targeting vaccination induces immune responses and reduces HIV-1 reservoirs. J. Clin. Investig. 2018, 128, 4387–4396. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.Y.; Osuna, C.E.; Hraber, P.T.; Hesselgesser, J.; Gerold, J.M.; Barnes, T.L.; Sanisetty, S.; Seaman, M.S.; Lewis, M.G.; Geleziunas, R.; et al. TLR7 agonists induce transient viremia and reduce the viral reservoir in SIV-infected rhesus macaques on antiretroviral therapy. Sci. Transl. Med. 2018, 10, eaao4521. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, G.Q.; Alvord, W.G.; Li, Y.; Deleage, C.; Nag, M.; Oswald, K.; Thomas, J.A.; Pyle, C.; Bosche, W.J.; Coalter, V.; et al. TLR7 agonist administration to SIV-infected macaques receiving early initiated cART does not induce plasma viremia. JCI Insight 2019, 4, e127717. [Google Scholar] [CrossRef] [Green Version]
- Borducchi, E.N.; Liu, J.; Nkolola, J.P.; Cadena, A.M.; Yu, W.-H.; Fischinger, S.; Broge, T.; Abbink, P.; Mercado, N.B.; Chandrashekar, A.; et al. Antibody and TLR7 agonist delay viral rebound in SHIV-infected monkeys. Nature 2018, 563, 360–364. [Google Scholar] [CrossRef]
- Moldt, B.; Chandrashekar, A.; Borducchi, E.N.; Nkolola, J.P.; Stephenson, H.; Nagel, M.; Hung, M.; Goldsmith, J.; Pace, C.S.; Carr, B.; et al. HIV envelope antibodies and TLR7 agonist partially prevent viral rebound in chronically SHIV-infected monkeys. PLoS Pathog. 2022, 18, e1010467. [Google Scholar] [CrossRef]
- Borducchi, E.N.; Cabral, C.; Stephenson, K.; Liu, J.; Abbink, P.; Ng’Ang’A, D.; Nkolola, J.P.; Brinkman, A.L.; Peter, L.; Lee, B.C.; et al. Ad26/MVA therapeutic vaccination with TLR7 stimulation in SIV-infected rhesus monkeys. Nature 2016, 540, 284–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker-Sperling, V.E.K.; Mercado, N.B.; Chandrashekar, A.; Borducchi, E.N.; Liu, J.; Nkolola, J.P.; Lewis, M.; Murry, J.P.; Yang, Y.; Geleziunas, R.; et al. Therapeutic efficacy of combined active and passive immunization in ART-suppressed, SHIV-infected rhesus macaques. Nat. Commun. 2022, 13, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Tang, X.; He, Y.; Bin Ju, B.; Wang, H. A Δ42PD1 fusion-expressing DNA vaccine elicits enhanced adaptive immune response to HIV-1: The key role of TLR4. Virol. J. 2022, 19, 174. [Google Scholar] [CrossRef] [PubMed]
- SenGupta, D.; Brinson, C.; DeJesus, E.; Mills, A.; Shalit, P.; Guo, S.; Cai, Y.; Wallin, J.J.; Zhang, L.; Humeniuk, R.; et al. The TLR7 agonist vesatomolid induced a modest delay in viral rebound in HIV controllers after cessation of antiretroviral therapy. Sci. Transl. Med. 2021, 13, 599. [Google Scholar] [CrossRef]
- A Riddler, S.; Para, M.; A Benson, C.; Mills, A.; Ramgopal, M.; DeJesus, E.; Brinson, C.; Cyktor, J.; Jacobs, J.; Koontz, D.; et al. Vesatolimod, a Toll-like Receptor 7 Agonist, Induces Immune Activation in Virally Suppressed Adults Living with Human Immunodeficiency Virus–1. Clin. Infect. Dis. 2021, 72, e815–e824. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rozman, M.; Zidovec-Lepej, S.; Jambrosic, K.; Babić, M.; Drmić Hofman, I. Role of TLRs in HIV-1 Infection and Potential of TLR Agonists in HIV-1 Vaccine Development and Treatment Strategies. Pathogens 2023, 12, 92. https://doi.org/10.3390/pathogens12010092
Rozman M, Zidovec-Lepej S, Jambrosic K, Babić M, Drmić Hofman I. Role of TLRs in HIV-1 Infection and Potential of TLR Agonists in HIV-1 Vaccine Development and Treatment Strategies. Pathogens. 2023; 12(1):92. https://doi.org/10.3390/pathogens12010092
Chicago/Turabian StyleRozman, Marija, Snjezana Zidovec-Lepej, Karlo Jambrosic, Maja Babić, and Irena Drmić Hofman. 2023. "Role of TLRs in HIV-1 Infection and Potential of TLR Agonists in HIV-1 Vaccine Development and Treatment Strategies" Pathogens 12, no. 1: 92. https://doi.org/10.3390/pathogens12010092