
Validating Immunomodulatory Responses of r-LdODC Protein and Its Derived HLA-DRB1 Restricted Epitopes against Visceral Leishmaniasis in BALB/c Mice

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Parasites
2.2. Soluble Leishmania Antigen (SLA)
2.3. Protein and Peptide Synthesis
2.4. Immunization of Mice
2.5. Splenic Mononuclear Cell Isolation
2.6. Quantification of Secretory IFN-γ in Protein and Peptides Cocktail Immunized Mice Group
2.7. Monitoring the T Cell Proliferation in Different Immunized Mice Groups
2.8. Measurement of Intracellular Cytokines Production and I-AD/I-ED Expression in Protein and Peptide Cocktail Immunized Mice
2.9. Monitoring the Parasite Load in the Macrophage
2.10. Statistical Analysis
3. Results
3.1. The r-LdODC Protein and Its Derived Synthetic Peptides Characterization
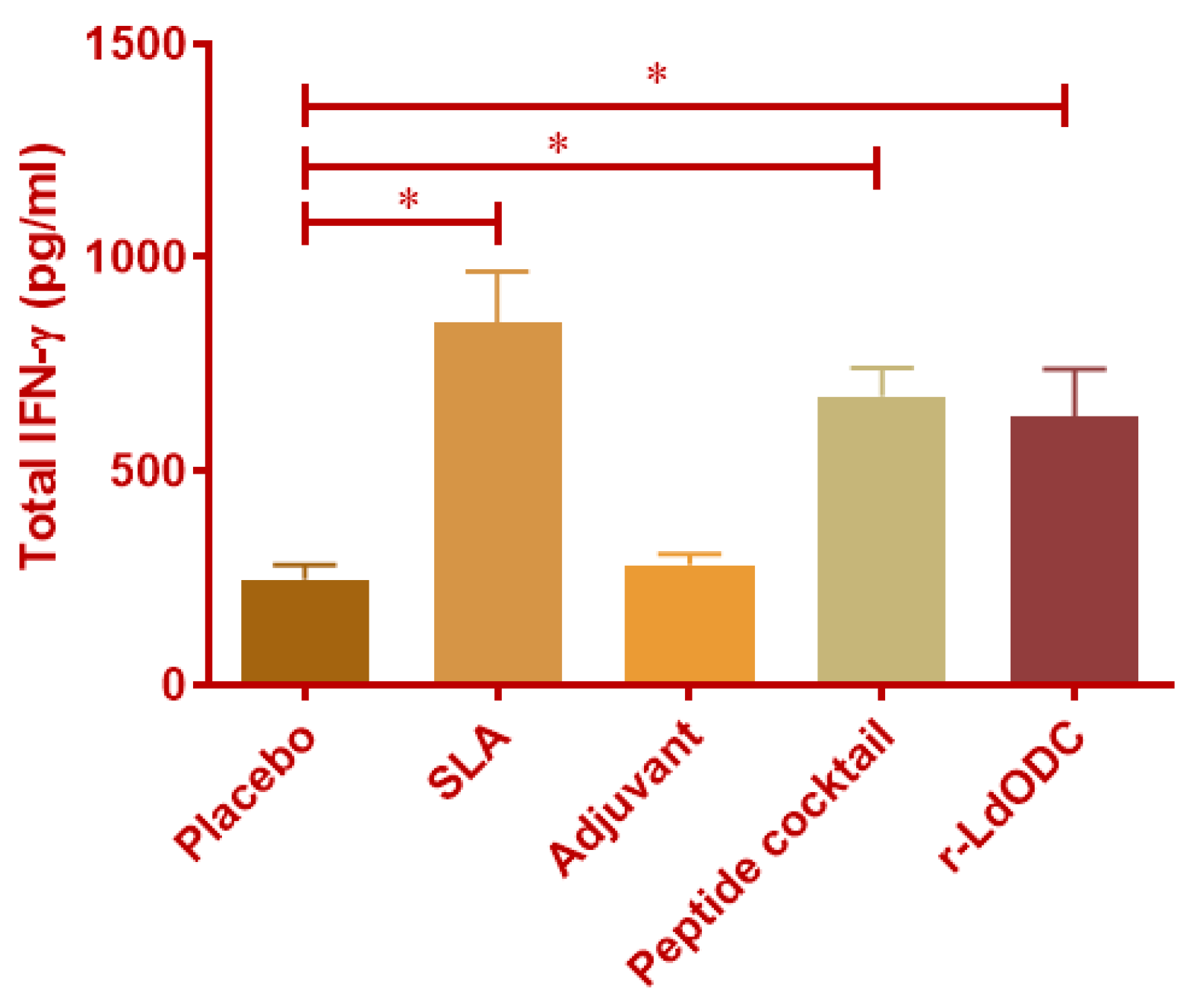
3.2. Quantification of IFN-γ by ELISA
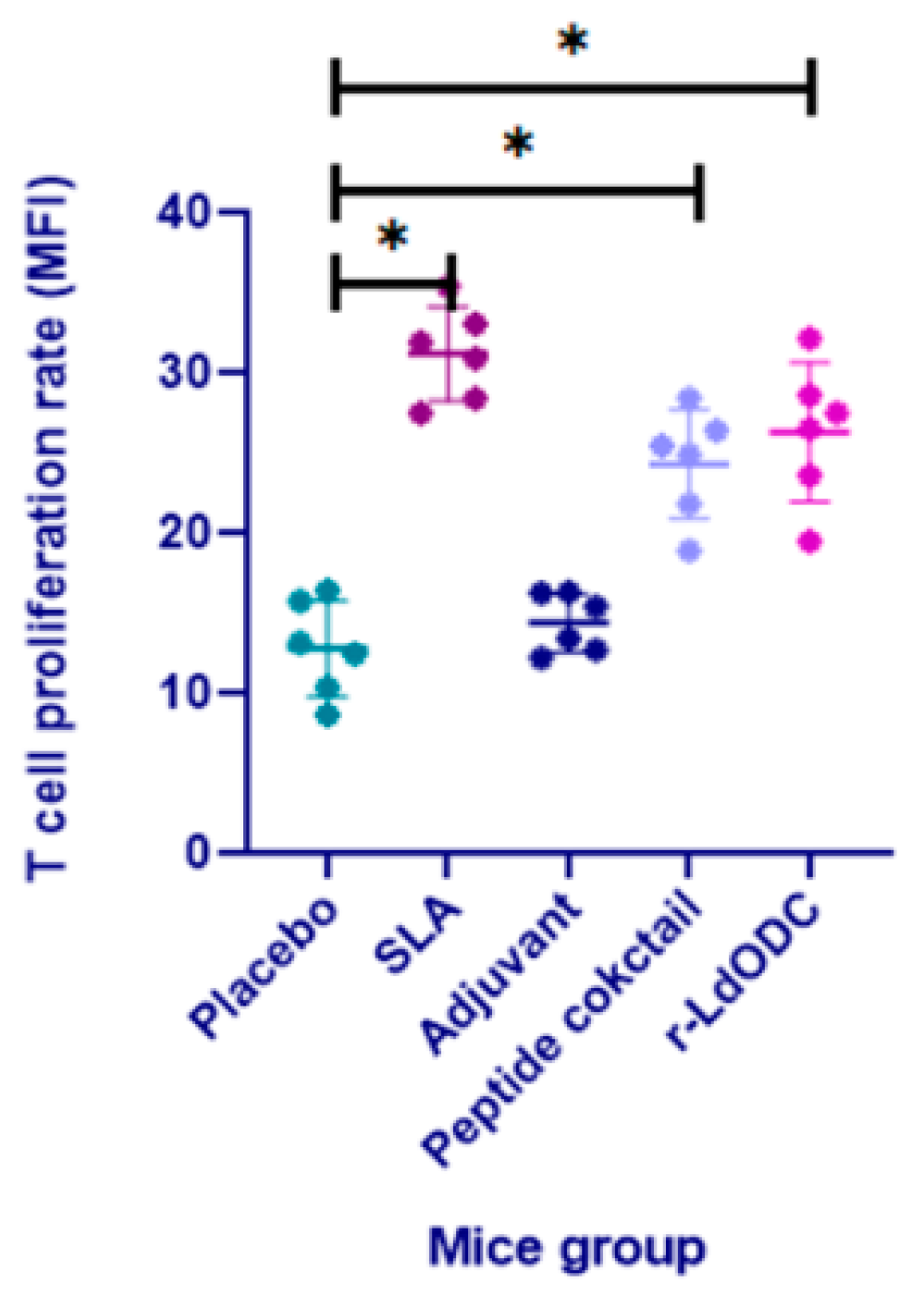
3.3. Measurement of T Cell Proliferation
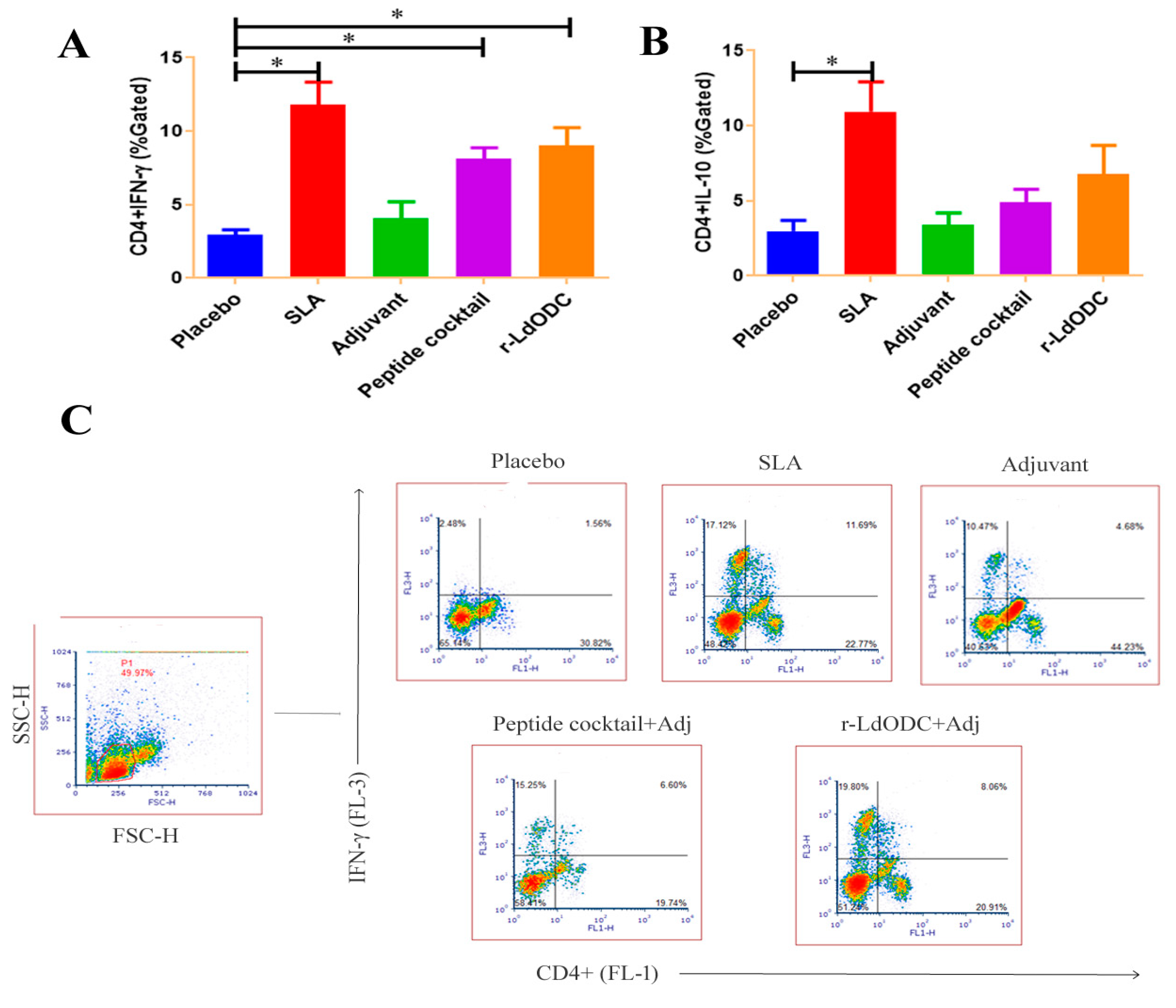
3.4. Assessment of Intracellular Cytokine Production against Protein and Peptides Cocktail
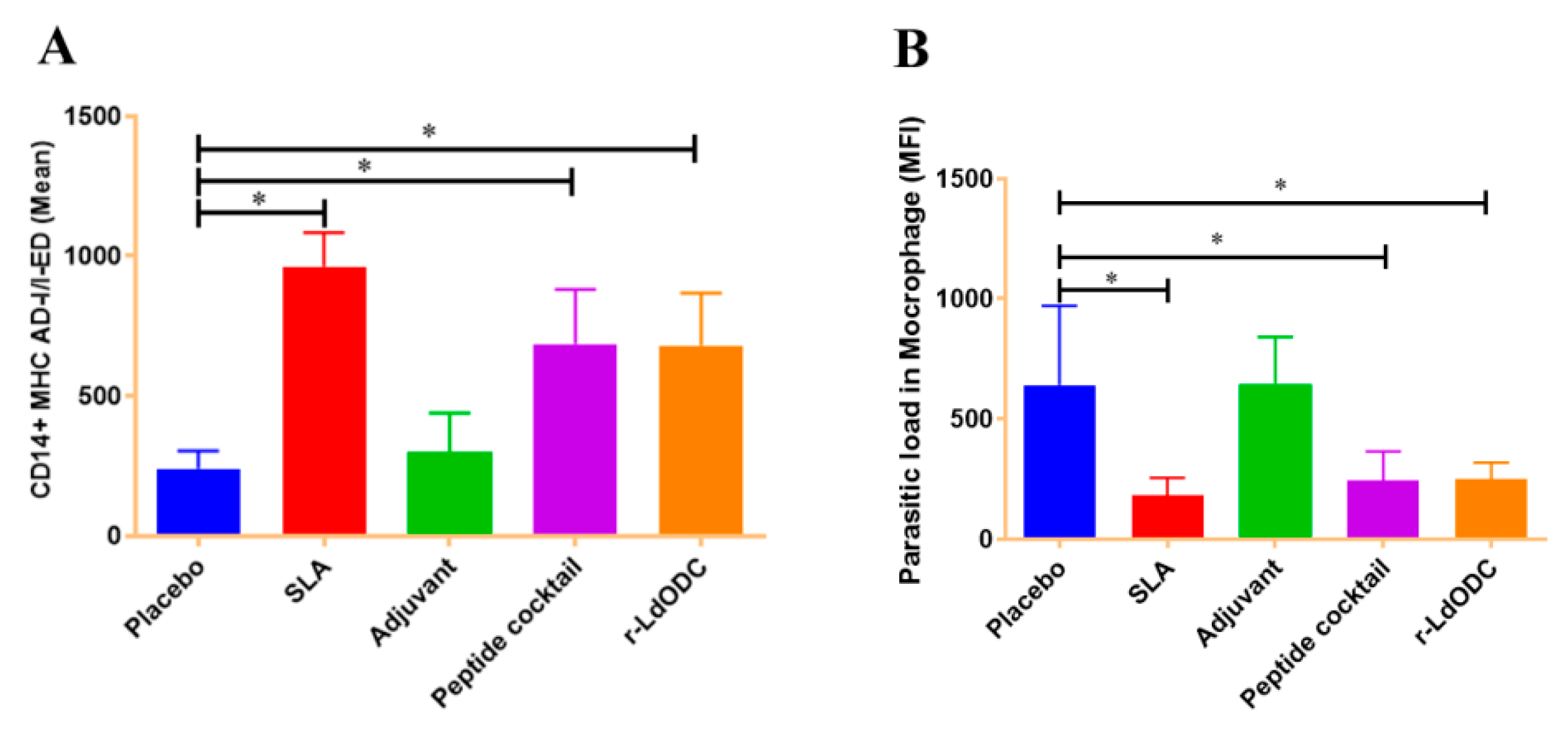
3.5. Measurement of Parasite Load in Macrophage
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Okwor, I.; Uzonna, J. Social and economic burden of human leishmaniasis. Am. J. Trop. Med. Hyg. 2016, 94, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Hirve, S.; Boelaert, M.; Matlashewski, G.; Mondal, D.; Arana, B.; Kroeger, A.; Olliaro, P. Transmission dynamics of visceral leishmaniasis in the indian subcontinent–a systematic literature review. PLoS Negl. Trop. Dis. 2016, 10, e0004896. [Google Scholar] [CrossRef] [Green Version]
- Rijal, S.; Sundar, S.; Mondal, D.; Das, P.; Alvar, J.; Boelaert, M. Eliminating visceral leishmaniasis in South Asia: The road ahead. BMJ 2019, 364, k5224. [Google Scholar] [CrossRef] [Green Version]
- Bhunia, G.S.; Kesari, S.; Chatterjee, N.; Kumar, V.; Das, P. The burden of visceral leishmaniasis in India: Challenges in using remote sensing and GIS to understand and control. ISRN Infect. Dis. 2012, 2013, 675846. [Google Scholar] [CrossRef] [Green Version]
- McCall, L.I.; Zhang, W.W.; Matlashewski, G. Determinants for the development of visceral leishmaniasis disease. PLoS Pathog. 2013, 9, e1003053. [Google Scholar] [CrossRef] [Green Version]
- Amit, A.; Dikhit, M.R.; Pandey, R.K.; Singh, K.; Mishra, R.; Das, V.R.; Das, P.; Bimal, S. Elevated serum ADA activity as a marker for diagnosis and prognosis of visceral leishmaniasis and post kala-azar dermal leishmaniasis in indian patients. PLoS ONE 2016, 11, e0154117. [Google Scholar]
- Moore, E.M.; Lockwood, D.N. Treatment of visceral leishmaniasis. J. Glob. Infect. Dis. 2010, 2, 151. [Google Scholar]
- Sundar, S.; Singh, A. Recent developments and future prospects in the treatment of visceral leishmaniasis. Ther. Adv. Infect. Dis. 2016, 3, 98–109. [Google Scholar] [CrossRef]
- Evans, K.J.; Kedzierski, L. Development of vaccines against visceral leishmaniasis. J. Trop. Med. 2012, 2012, 892817. [Google Scholar] [CrossRef]
- Costa, C.H.N.; Peters, N.C.; Maruyama, S.R.; de Brito, E.C., Jr.; de Miranda Santos, I.K.F.; Working Group on Research Priorities for Development of Leishmaniasis Vaccines. Vaccines for the leishmaniases: Proposals for a research agenda. PLoS Negl. Trop. Dis. 2011, 5, e943. [Google Scholar]
- Lodge, R.; Descoteaux, A. Leishmania invasion and phagosome biogenesis. In Molecular Mechanisms of Parasite Invasion; Springer: New York, NY, USA, 2008; pp. 174–181. [Google Scholar]
- Pakpour, N.; Zaph, C.; Scott, P. The central memory CD4+T cell population generated during Leishmania major infection requires IL-12 to produce IFN-γ. J. Immunol. 2008, 180, 8299–8305. [Google Scholar] [CrossRef] [Green Version]
- Pearson, R.D.; Romito, R.O.S.E.A.N.N.; Symes, P.H.; Harcus, J.L. Interaction of Leishmania donovani promastigotes with human monocyte-derived macrophages: Parasite entry, intracellular survival, and multiplication. Infect. Immun. 1981, 32, 1249–1253. [Google Scholar] [CrossRef] [Green Version]
- Aderem, A.; Underhill, D.M. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef]
- Dayakar, A.; Chandrasekaran, S.; Kuchipudi, S.V.; Kalangi, S.K. Cytokines: Key determinants of resistance or disease progression in visceral leishmaniasis: Opportunities for novel diagnostics and immunotherapy. Front. Immunol. 2019, 10, 670. [Google Scholar] [CrossRef] [Green Version]
- Pandey, R.; Dikhit, M.R.; Kumar, A.; Dehury, B.; Pandey, K.; Topno, R.K.; Das, P.; Bimal, S. Evaluating the immunomodulatory responses of LdODC-derived MHC Class-II restricted peptides against VL. Parasite Immunol. 2020, 42, e12699. [Google Scholar] [CrossRef]
- Andersen, M.H.; Tan, L.; Søndergaard, I.; Zeuthen, J.; Elliott, T.; Haurum, J.S. Poor correspondence between predicted and experimental binding of peptides to class I MHC molecules. Tissue Antigens 2000, 55, 519–531. [Google Scholar] [CrossRef]
- Joshi, S.; Rawat, K.; Yadav, N.K.; Kumar, V.; Siddiqi, M.I.; Dube, A. Visceral leishmaniasis: Advancements in vaccine development via classical and molecular approaches. Front. Immunol. 2014, 5, 380. [Google Scholar] [CrossRef] [Green Version]
- Skeiky, Y.A.; Coler, R.N.; Brannon, M.; Stromberg, E.; Greeson, K.; Crane, R.T.; Campos-Neto, A.; Reed, S.G. Protective efficacy of a tandemly linked, multi-subunit recombinant leishmanial vaccine (Leish-111f) formulated in MPL® adjuvant. Vaccine 2002, 20, 3292–3303. [Google Scholar] [CrossRef]
- Raman, V.S.; Reed, S.G.; Duthie, M.S.; Fox, C.B.; Matlashewski, G. Adjuvants for Leishmania vaccines: From models to clinical application. Front. Immunol. 2012, 3, 144. [Google Scholar] [CrossRef] [Green Version]
- Bimal, S.; Singh, S.K.; Sinha, S.; Pandey, K.; Sinha, P.K.; Ranjan, A.; Bhattacharya, S.K.; Das, P. Leishmania donovani: Role of CD2 on CD4+T-cell function in Visceral Leishmaniasis. Exp. Parasitol. 2008, 118, 238–246. [Google Scholar] [CrossRef]
- Killeen, N.; Stuart, S.G.; Littman, D.R. Development and function of T cells in mice with a disrupted CD2 gene. EMBO J. 1992, 11, 4329–4336. [Google Scholar] [CrossRef] [PubMed]
- Légaré, D.; Richard, D.; Mukhopadhyay, R.; Stierhof, Y.D.; Rosen, B.P.; Haimeur, A.; Papadopoulou, B.; Ouellette, M. The Leishmania ATP-binding cassette protein PGPA is an intracellular metal-thiol transporter ATPase. J. Biol. Chem. 2001, 276, 26301–26307. [Google Scholar] [CrossRef] [Green Version]
- Dey, S.; Ouellette, M.; Lightbody, J.; Papadopoulou, B.; Rosen, B.P. An ATP-dependent As (III)-glutathione transport system in membrane vesicles of Leishmania tarentolae. Proc. Natl. Acad. Sci. USA 1996, 93, 2192–2197. [Google Scholar] [CrossRef] [PubMed]
- Haimeur, A.; Brochu, C.; Genest, P.A.; Papadopoulou, B.; Ouellette, M. Amplification of the ABC transporter gene PGPA and increased trypanothione levels in potassium antimonyl tartrate (SbIII) resistant Leishmania tarentolae. Mol. Biochem. Parasitol. 2000, 108, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Amit, A.; Chaudhary, R.; Chandel, A.S.; Mahantesh, V.; Suman, S.S.; Singh, S.K.; Dikhit, M.R.; Ali, V.; Rabidas, V.; et al. Leishmania donovani: Impairment of the cellular immune response against recombinant ornithine decarboxylase protein as a possible evasion strategy of Leishmania in visceral leishmaniasis. Int. J. Parasitol. 2015, 45, 33–42. [Google Scholar] [CrossRef]
- Kumar, A.; Dikhit, M.R.; Amit, A.; Zaidi, A.; Pandey, R.K.; kumar Singh, A.; Suman, S.S.; Ali, V.; Das, V.N.R.; Pandey, K.; et al. Immunomodulation induced through ornithine decarboxylase DNA immunization in Balb/c mice infected with Leishmania donovani. Mol. Immunol. 2018, 97, 33–44. [Google Scholar] [CrossRef]
- Pandey, R.K.; Dikhit, M.R.; Lokhande, K.B.; Pandey, K.; Das, P.; Bimal, S. An immunoprophylactic evaluation of Ld-ODC derived HLA-A0201 restricted peptides against visceral leishmaniasis. J. Biomol. Struct. Dyn. 2021, 40, 1–11. [Google Scholar] [CrossRef]
- Morozov, A.V.; Morozov, V.A.; Astakhova, T.M.; Timofeev, A.V.; Karpov, V.L. DNA vaccine encoding α-fetoprotein fused with the ornithine decarboxylase degradation signal significantly suppresses the hepatocellular carcinoma growth in mice. Mol. Biol. 2012, 46, 391–406. [Google Scholar] [CrossRef]
- Dikhit, M.R.; Kumar, A.; Das, S.; Dehury, B.; Rout, A.K.; Jamal, F.; Sahoo, G.C.; Topno, R.K.; Pandey, K.; Das, V.N.R.; et al. Identification of potential MHC Class-II-restricted epitopes derived from Leishmania donovani antigens by reverse vaccinology and evaluation of their CD4+T-cell responsiveness against visceral leishmaniasis. Front. Immunol. 2017, 8, 1763. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Engwerda, C. Vaccines to prevent leishmaniasis. Clin. Transl. Immunol. 2014, 3, e13. [Google Scholar] [CrossRef]
- Wang, P.; Sidney, J.; Kim, Y.; Sette, A.; Lund, O.; Nielsen, M.; Peters, B. Peptide binding predictions for HLA DR, DP and DQ molecules. BMC Bioinform. 2010, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sospedra, M.; Muraro, P.A.; Stefanová, I.; Zhao, Y.; Chung, K.; Li, Y.; Giulianotti, M.; Simon, R.; Mariuzza, R.; Pinilla, C.; et al. Redundancy in antigen-presenting function of the HLA-DR and-DQ molecules in the multiple sclerosis-associated HLA-DR2 haplotype. J. Immunol. 2006, 176, 1951–1961. [Google Scholar] [CrossRef] [Green Version]
- Jucaud, V. The immunogenicity of HLA class II mismatches: The predicted presentation of nonself allo-HLA-derived peptide by the HLA-DR phenotype of the recipient is associated with the formation of DSA. J. Immunol. Res. 2017, 2017, 2748614. [Google Scholar] [CrossRef]
- Reiner, S.L.; Zheng, S.; Wang, Z.E.; Stowring, L.; Locksley, R.M. Leishmania promastigotes evade interleukin 12 (IL-12) induction by macrophages and stimulate a broad range of cytokines from CD4+T cells during initiation of infection. J. Exp. Med. 1994, 179, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Kemp, K.; Kemp, M.; Kharazmi, A.; Ismail, A.; Kurtzhals, J.A.L.; Hviid, L.; Theander, T.G. Leishmania-specific T cells expressing interferon-gamma (IFN-γ) and IL-10 upon activation are expanded in individuals cured of visceral leishmaniasis. Clin. Exp. Immunol. 1999, 116, 500. [Google Scholar] [CrossRef]
- Kane, M.M.; Mosser, D.M. The role of IL-10 in promoting disease progression in leishmaniasis. J. Immunol. 2001, 166, 1141–1147. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Singh, N.; Gautam, S.; Singh, O.P.; Gidwani, K.; Rai, M.; Sacks, D.; Sundar, S.; Nylén, S. Leishmania specific CD4 T cells release IFNγ that limits parasite replication in patients with visceral leishmaniasis. PLoS Negl. Trop. Dis. 2014, 8, e3198. [Google Scholar] [CrossRef]
- Andargie, T.E.; Ejara, E.D. Pro-and anti-inflammatory cytokines in visceral leishmaniasis. J. Cell Sci. Ther. 2015, 6, 1. [Google Scholar] [CrossRef]
- Lage, D.P.; Ribeiro, P.A.; Dias, D.S.; Mendonça, D.V.; Ramos, F.F.; Carvalho, L.M.; de Oliveira, D.; Steiner, B.T.; Martins, V.T.; Perin, L.; et al. A candidate vaccine for human visceral leishmaniasis based on a specific T cell epitope-containing chimeric protein protects mice against Leishmania infantum infection. NPJ Vaccines 2020, 5, 1–13. [Google Scholar] [CrossRef]
- Roy, K.; Naskar, K.; Ghosh, M.; Roy, S. Class II MHC/peptide interaction in Leishmania donovani infection: Implications in vaccine design. J. Immunol. 2014, 192, 5873–5880. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.; Fasel, N. How to master the host immune system? Leishmania parasites have the solutions! Int. Immunol. 2018, 30, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Oghumu, S.; Satoskar, A.R. Mechanisms of immune evasion in leishmaniasis. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 2013; Volume 82, pp. 155–184. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandey, R.; Gautam, R.K.; Sharma, S.; Tedla, M.G.; Mahantesh, V.; Dikhit, M.R.; Kumar, A.; Pandey, K.; Bimal, S. Validating Immunomodulatory Responses of r-LdODC Protein and Its Derived HLA-DRB1 Restricted Epitopes against Visceral Leishmaniasis in BALB/c Mice. Pathogens 2023, 12, 16. https://doi.org/10.3390/pathogens12010016
Pandey R, Gautam RK, Sharma S, Tedla MG, Mahantesh V, Dikhit MR, Kumar A, Pandey K, Bimal S. Validating Immunomodulatory Responses of r-LdODC Protein and Its Derived HLA-DRB1 Restricted Epitopes against Visceral Leishmaniasis in BALB/c Mice. Pathogens. 2023; 12(1):16. https://doi.org/10.3390/pathogens12010016
Chicago/Turabian StylePandey, Rajkishor, Rohit Kumar Gautam, Simran Sharma, Mebrahtu G. Tedla, Vijay Mahantesh, Manas Ranjan Dikhit, Akhilesh Kumar, Krishna Pandey, and Sanjiva Bimal. 2023. "Validating Immunomodulatory Responses of r-LdODC Protein and Its Derived HLA-DRB1 Restricted Epitopes against Visceral Leishmaniasis in BALB/c Mice" Pathogens 12, no. 1: 16. https://doi.org/10.3390/pathogens12010016