
The Never-Ending Presence of Phytophthora Species in Italian Nurseries


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Phytophthora Species Diversity in Plant Nurseries
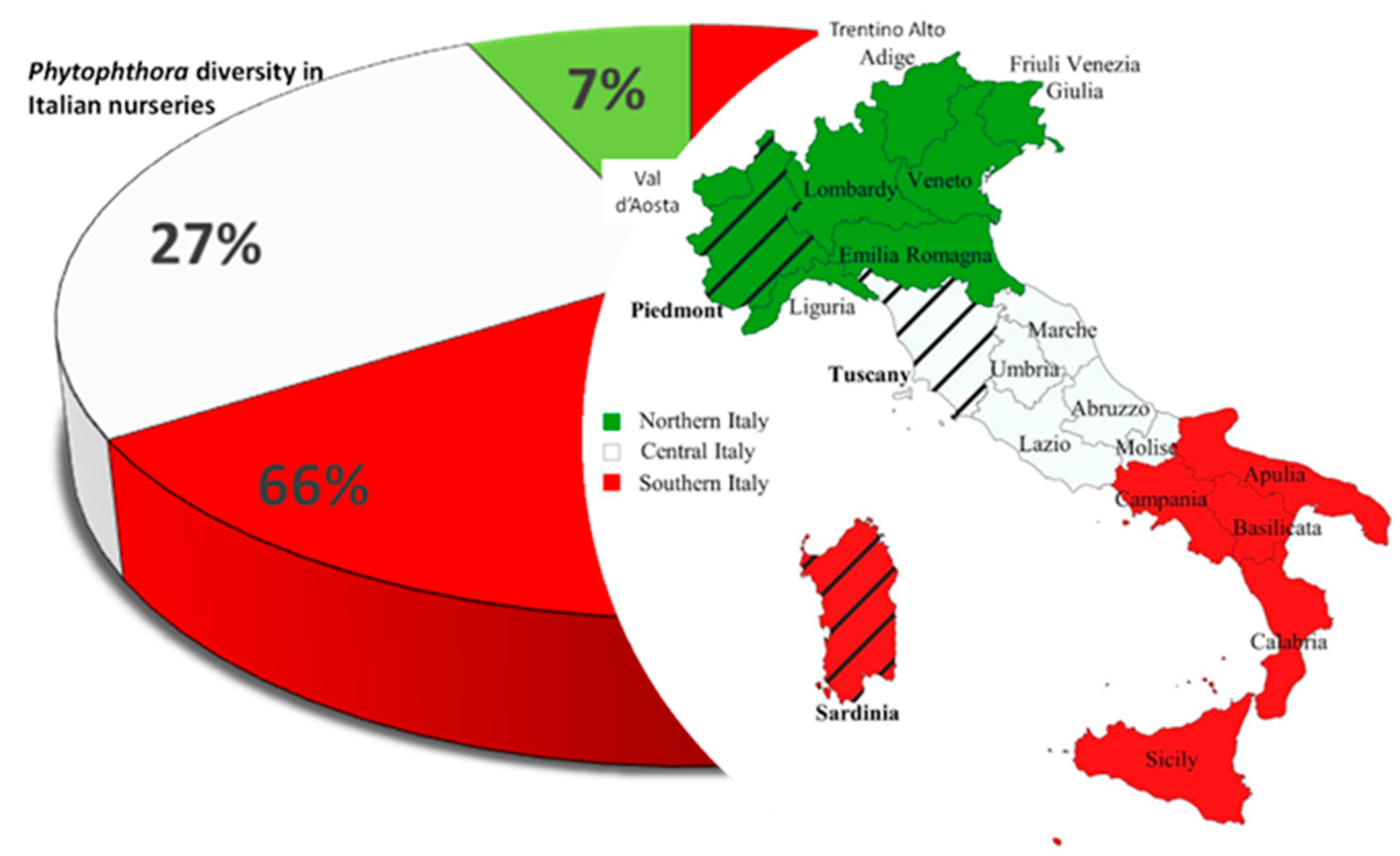
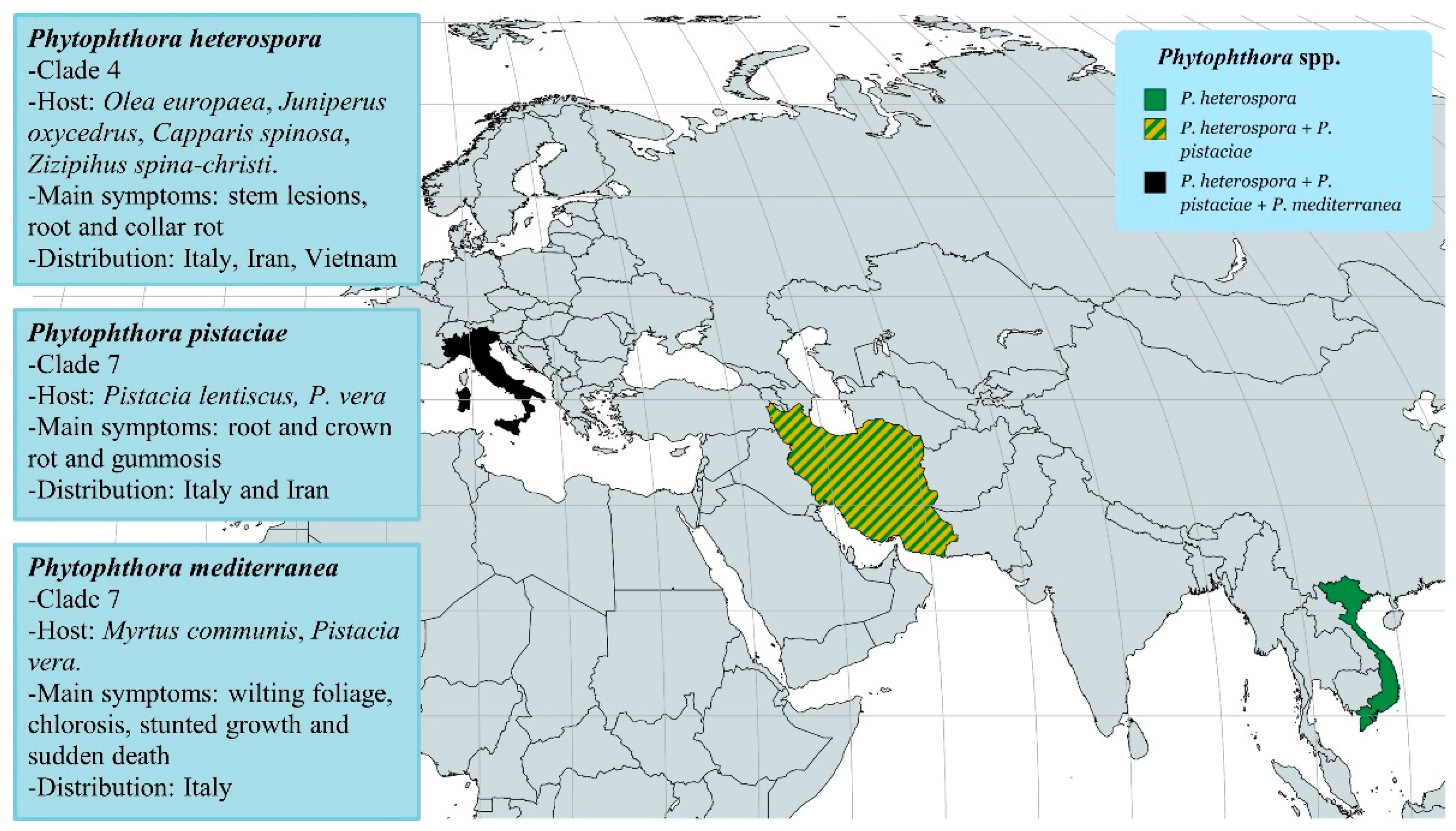
2.1. The Case of Phytophthora Species in Italian Plant Nurseries
2.2. Detection Methods for Phytophthora Species in Italian Nurseries
3. How to Tackle the Spread of Phytophthora Species
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press—American Phytopathological Society: St. Paul, MN, USA, 1996; 562p. [Google Scholar]
- Jung, T.; Colquhoun, I.J.; Hardy, G.E.S.J. New insights into the survival strategy of the invasive soilborne pathogen Phytophthora cinnamomi in different natural ecosystems in Western Australia. For. Path. 2013, 43, 266–288. [Google Scholar] [CrossRef]
- Vannini, A.; Breccia, M.; Bruni, N.; Tomassini, A.; Vettraino, A.M. Behaviour and survival of Phytophthora cambivora inoculum in soil-like substrate under different water regimes: Behavior and survival of Phytophthora cambivora. For. Path. 2012, 42, 362–370. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Tomassini, A.; Vannini, A. Use of mRNA as an indicator of the viability of Phytophthora cambivora. Acta Hortic. 2010, 866, 431–434. [Google Scholar] [CrossRef]
- Denman, S.; Kirk, S.A.; Brasier, C.M.; Barton, V.C.; Hughes, K.J.D.; Webber, J.F. Phytophthora ramorum on Quercus ilex in the United Kingdom. Plant Dis. 2005, 89, 1241. [Google Scholar] [CrossRef] [PubMed]
- Grünwald, N.J.; Goss, E.M.; Press, C.M. Phytophthora ramorum: A pathogen with a remarkably wide host range causing Sudden Oak Death on oaks and ramorum blight on woody ornamentals. Mol. Plant Pathol. 2008, 9, 729–740. [Google Scholar] [CrossRef]
- Lindqvist-Kreuze, H.; Gamboa, S.; Izarra, M.; Pérez, W.; Correa, M.Y.; Astete, A.; Särkinen, T.; Cueva, M.; Gonzáles, P. Population structure and host range of the potato late blight pathogen Phytophthora infestans in Peru Spanning Two Decades. Plant Pathol. 2020, 69, 334–346. [Google Scholar] [CrossRef] [Green Version]
- Tooley, P.W.; Kyde, K.L. Susceptibility of some eastern forest species to Phytophthora ramorum. Plant Dis. 2007, 91, 435–438. [Google Scholar] [CrossRef] [Green Version]
- Vettraino, A.M.; Hüberli, D.; Garbelotto, M. Phytophthora ramorum infection of coast live oak leaves in California forests and its capacity to sporulate in vitro. Austral. Plant Pathol. 2008, 37, 72. [Google Scholar] [CrossRef]
- Carleson, N.C.; Daniels, H.A.; Reeser, P.W.; Kanaskie, A.; Navarro, S.M.; LeBoldus, J.M.; Grünwald, N.J. Novel introductions and epidemic dynamics of the Sudden Oak Death pathogen Phytophthora ramorum in Oregon Forests. Phytopathology 2021, 111, 731–740. [Google Scholar] [CrossRef]
- Davison, E.M.; Drenth, A.; Kumar, S.; Mack, S.; Mackie, A.E.; McKirdy, S. Pathogens associated with nursery plants imported into Western Australia. Austral. Plant Pathol. 2006, 35, 473. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Brockerhoff, E.G.; Garrett, L.J.; Parke, J.L.; Britton, K.O. Live plant imports: The major pathway for forest insect and pathogen invasions of the US. Front. Ecol. Environ. 2012, 10, 135–143. [Google Scholar] [CrossRef]
- Santini, A.; Ghelardini, L.; Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographical patterns and determinants of invasion by forest pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vettraino, A.M.; Santini, A. Forest health in Italy: Learning from the Xylella incursion. Front. For. Glob. Chang. 2021, 4, 699393. [Google Scholar] [CrossRef]
- Brasier, C.; Webber, J. Sudden Larch Death. Nature 2010, 466, 824–825. [Google Scholar] [CrossRef]
- Rizzo, D.M.; Garbelotto, M.; Davidson, J.M.; Slaughter, G.W.; Koike, S.T. Phytophthora ramorum as the cause of extensive mortality of Quercus spp. and Lithocarpus densiflorus in California. Plant Dis. 2002, 86, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Goss, E.M.; Larsen, M.; Vercauteren, A.; Werres, S.; Heungens, K.; Grünwald, N.J. Phytophthora ramorum in Canada: Evidence for Migration Within North America and from Europe. Phytopathology 2011, 101, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Grünwald, N.J.; Garbelotto, M.; Goss, E.M.; Heungens, K.; Prospero, S. Emergence of the Sudden Oak Death pathogen Phytophthora ramorum. Trends Microbiol. 2012, 20, 131–138. [Google Scholar] [CrossRef]
- Mascheretti, S.; Croucher, P.J.P.; Vettraino, A.; Prospero, S.; Garbelotto, M. Reconstruction of the Sudden Oak Death Epidemic in California through microsatellite analysis of the pathogen Phytophthora ramorum: Genetic structure of P. ramorum in California. Mol. Ecol. 2008, 17, 2755–2768. [Google Scholar] [CrossRef]
- Mascheretti, S.; Croucher, P.J.P.; Kozanitas, M.; Baker, L.; Garbelotto, M. Genetic epidemiology of the Sudden Oak Death pathogen Phytophthora ramorum in California. Mol. Ecol. 2009, 18, 4577–4590. [Google Scholar] [CrossRef]
- Werres, S.; Marwitz, R.; Veld, W.A.M.I.; Cock, A.W.A.M.D.; Bonants, P.J.M.; Weerdt, M.D.; Themann, K.; Ilieva, E.; Baayen, R.P. Phytophthora ramorum sp. nov., a new pathogen on Rhododendron and Viburnum. Mycol. Res. 2001, 105, 1155–1165. [Google Scholar] [CrossRef]
- Elliott, M.; Rollins, L.; Bourret, T.; Hulbert, J.M.; Chastagner, G. Three new hosts for Phytophthora ramorum confirmed in Washington State: Salal, Oregon Grape, and Red Huckleberry. Plant Health Prog. 2021, 22, 389–391. [Google Scholar] [CrossRef]
- Hansen, E.M.; Goheen, D.J.; Jules, E.S.; Ullian, B. Managing Port-Orford-Cedar and the introduced pathogen Phytophthora lateralis. Plant Dis. 2000, 84, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbelotto, M.; Frankel, S.J.; Scanu, B. Soil- and waterborne Phytophthora species linked to recent outbreaks in Northern California restoration sites. Calif. Agr. 2018, 72, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Rooney-Latham, S.; Blomquist, C.L.; Swiecki, T.; Bernhardt, E.; Frankel, S.J. First detection in the US: New plant pathogen, Phytophthora tentaculata, in native plant nurseries and restoration sites in California. Nativ. Plants J. 2015, 16, 23–27. [Google Scholar] [CrossRef]
- Sims, L.L.; Garbelotto, M. Susceptibility to the Rare Phytophthora tentaculata and to the widespread Phytophthora cactorum is consistent with host ecology and history. For. Path. 2018, 48, e12446. [Google Scholar] [CrossRef]
- Themann, K.; Werres, S.; Lüttmann, R.; Diener, H.-A. Observations of Phytophthora spp. in water recirculation systems in commercial hardy ornamental nursery stock. Eur. J. Plant Pathol. 2002, 108, 337–343. [Google Scholar] [CrossRef]
- Eschen, R.; Rigaux, L.; Sukovata, L.; Vettraino, A.M.; Marzano, M.; Grégoire, J.-C. Phytosanitary inspection of woody plants for planting at European union entry points: A practical enquiry. Biol. Invasions 2015, 17, 2403–2413. [Google Scholar] [CrossRef] [Green Version]
- Marzano, M.; Dandy, N.; Papazova-Anakieva, I.; Avtzis, D.; Connolly, T.; Eschen, R.; Glavendekić, M.; Hurley, B.; Lindelöw, Å.; Matošević, D.; et al. Assessing awareness of tree pests and pathogens amongst tree professionals: A pan-European perspective. For. Policy Econ. 2016, 70, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Migliorini, D.; Ghelardini, L.; Tondini, E.; Luchi, N.; Santini, A. The potential of symptomless potted plants for carrying invasive soilborne plant pathogens. Divers. Distrib. 2015, 21, 1218–1229. [Google Scholar] [CrossRef] [Green Version]
- Marin, M.V.; Seijo, T.E.; Zuchelli, E.; Peres, N.A. Resistance to mefenoxam of Phytophthora cactorum and Phytophthora nicotianae causing crown and leather rot in Florida strawberry. Plant Dis. 2021, 105, 3490–3495. [Google Scholar] [CrossRef]
- Olson, H.A.; Jeffers, S.N.; Ivors, K.L.; Steddom, K.C.; Williams-Woodward, J.L.; Mmbaga, M.T.; Benson, D.M.; Hong, C.X. Diversity and mefenoxam sensitivity of Phytophthora spp. associated with the ornamental horticulture industry in the Southeastern United States. Plant Dis. 2013, 97, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasier, C.M.; Kirk, S.A.; Delcan, J.; Cooke, D.E.L.; Jung, T.; Man In’t Veld, W.A. Phytophthora alni sp. nov. and its variants: Designation of emerging heteroploid hybrid pathogens spreading on Alnus Trees. Mycol. Res. 2004, 108, 1172–1184. [Google Scholar] [CrossRef] [PubMed]
- Nirenberg, H.I.; Gerlach, W.F.; Gräfenhan, T. Phytophthora × pelgrandis, a new natural hybrid pathogenic to Pelargonium grandiflorum Hort. Mycologia 2009, 101, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Path. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.; Scanu, B.; Cooke, D.; Jung, T. Phytophthora: An ancient, historic, biologically and structurally cohesive and evolutionarily successful generic concept in need of preservation. IMA Fungus 2022, 13, 12. [Google Scholar] [CrossRef]
- Burgess, T.I.; White, D.; McDougall, K.M.; Garnas, J.; Dunstan, W.A.; Català, S.; Carnegie, A.J.; Worboys, S.; Cahill, D.; Vettraino, A.M.; et al. Distribution and diversity of Phytophthora across Australia. Pac. Conserv. Biol. 2017, 23, 150. [Google Scholar] [CrossRef] [Green Version]
- Hardy, G.E.; Sivasithamparam, K. Phytophthora spp. associated with container-grown plants in nurseries in Western Australia. Plant Dis. 1988, 72, 435. [Google Scholar] [CrossRef] [Green Version]
- Scott, P.; Williams, N. Phytophthora diseases in New Zealand Forests. NZ J. For. 2014, 59, 15. [Google Scholar]
- Simamora, A.V.; Stukely, M.J.C.; Barber, P.A.; Hardy, G.E.S.; Burgess, T.I. Age-Related susceptibility of Eucalyptus species to Phytophthora boodjera. Plant Pathol. 2017, 66, 501–512. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Barzanti, G.P.; Bianco, M.C.; Ragazzi, A.; Capretti, P.; Paoletti, E.; Luisi, N.; Anselmi, N.; Vannini, A. Occurrence of Phytophthora species in oak stands in Italy and their association with declining oak trees. For. Pathol. 2002, 32, 19–28. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Morel, O.; Perlerou, C.; Robin, C.; Diamandis, S.; Vannini, A. occurrence and distribution of Phytophthora species in European chestnut stands, and their association with ink disease and crown decline. Eur. J. Plant Pathol. 2005, 111, 169–180. [Google Scholar] [CrossRef]
- Panabieres, F.; Ali, G.S.; Allagui, M.B.; Dalio, R.J.D.; Gudmestad, N.C.; Kuhn, M.-L.; Guha Roy, S.; Schena, L.; Zampounis, A. Phytophthora nicotianae diseases worldwide: New knowledge of a long-recognized pathogen. Phytopathol. Mediterr. 2016, 55, 20–40. [Google Scholar] [CrossRef]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Bourson, P.H. Exposé analytique de diverses opinions sur les causes probables de la maladie des pommes de terre, présenté à la commission instituée près du Ministère d’Intérieur. Monit. Belg. 1845, 18, 2695–2699. [Google Scholar]
- Bourke, P.M.A. Emergence of potato blight. Nature 1964, 203, 805–808. [Google Scholar] [CrossRef]
- Reader, J. Potato: A History of the Propitious Esculent; Yale University Press: London, UK, 2009. [Google Scholar]
- Knaus, B.J.; Fieland, V.J.; Graham, K.A.; Grünwald, N.J. Diversity of foliar Phytophthora species on Rhododendron in Oregon nurseries. Plant Dis. 2015, 99, 1326–1332. [Google Scholar] [CrossRef] [Green Version]
- Burgess, T.I.; Edwards, J.; Drenth, A.; Massenbauer, T.; Cunnington, J.; Mostowfizadeh-Ghalamfarsa, R.; Dinh, Q.; Liew, E.C.Y.; White, D.; Scott, P.; et al. Current status of Phytophthora in Australia. Persoonia 2021, 47, 151–177. [Google Scholar] [CrossRef]
- Langenhoven, S.; Halleen, F.; Spies, C.F.J.; Stempien, E.; Mostert, L. Detection and quantification of black foot and crown and root rot pathogens in grapevine nursery soils in the Western Cape of South Africa. Phytopathol. Mediterr. 2018, 57, 519–537. [Google Scholar] [CrossRef]
- Simamora, A.V.; Stukely, M.J.C.; Hardy, G.E.S.; Burgess, T.I. Phytophthora boodjera sp. nov., a damping-off pathogen in production nurseries and from urban and natural landscapes, with an update on the status of P. alticola. IMA Fungus 2015, 6, 319–335. [Google Scholar] [CrossRef] [Green Version]
- Parke, J.L.; Knaus, B.J.; Fieland, V.J.; Lewis, C.; Grünwald, N.J. Phytophthora community structure analyses in Oregon nurseries inform systems approaches to disease management. Phytopathology 2014, 104, 1052–1062. [Google Scholar] [CrossRef] [Green Version]
- Rooney-Latham, S.; Blomquist, C.L.; Kosta, K.L.; Gou, Y.Y.; Woods, P.W. Phytophthora species are common on nursery stock grown for restoration and revegetation purposes in California. Plant Dis. 2019, 103, 448–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, E.M.; Streito, J.-C.; Delatour, C. First confirmation of Phytophthora lateralis in Europe. Plant Dis. 1999, 83, 587. [Google Scholar] [CrossRef] [PubMed]
- Molnar, C.; Nikolaeva, E.; Kim, S.; Olson, T.; Bily, D.; Kim, J.-E.; Kang, S. Phytophthora diversity in Pennsylvania nurseries and greenhouses inferred from clinical samples collected over four decades. Microorganisms 2020, 8, 1056. [Google Scholar] [CrossRef] [PubMed]
- USDA APHIS Home Landing Page. Available online: https://www.aphis.usda.gov/aphis/home/ (accessed on 1 September 2022).
- Mora-Sala, B.; León, M.; Pérez-Sierra, A.; Abad-Campos, P. New reports of Phytophthora species in plant nurseries in Spain. Pathogens 2022, 11, 826. [Google Scholar] [CrossRef] [PubMed]
- Mora-Sala, B.; Berbegal, M.; Abad-Campos, P. The Use of QPCR Reveals a High Frequency of Phytophthora quercina in Two Spanish Holm Oak Areas. Forests 2018, 9, 697. [Google Scholar] [CrossRef] [Green Version]
- Moralejo, E.; Pérez-Sierra, A.M.; Álvarez, L.A.; Belbahri, L.; Lefort, F.; Descals, E. Multiple alien Phytophthora taxa discovered on diseased ornamental plants in Spain. Plant Pathol. 2009, 58, 100–110. [Google Scholar] [CrossRef]
- Henricot, B.; Pérez Sierra, A.; Jung, T. Phytophthora pachypleura sp. nov., a new species causing root rot of Aucuba japonica and other ornamentals in the United Kingdom. Plant Pathol. 2014, 63, 1095–1109. [Google Scholar] [CrossRef] [Green Version]
- Gullino, C.; Garofalo, M.C.; Moretti, F.; Gianetti, G.; Mainenti, E. Rinvenimento su rododendro di Phytophthora ramoru. L’Inf. Agrar. 2003, 19, 87–89. [Google Scholar] [CrossRef]
- Ginetti, B.; Carmignani, S.; Ragazzi, A.; Werres, S.; Moricca, S. Foliar blight and shoot dieback caused by Phytophthora ramorum on Viburnum tinus in the Pistoia area, Tuscany, central Italy. Plant Dis. 2014, 98, 423. [Google Scholar] [CrossRef]
- Vannini, A.; Bruni, N.; Tomassini, A.; Franceschini, S.; Vettraino, A.M. Pyrosequencing of environmental soil samples reveals biodiversity of the Phytophthora resident community in chestnut forests. FEMS Microbiol. Ecol. 2013, 85, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Scopus Welcome to Scopus. Available online: https://www.scopus.com/home.uri (accessed on 1 October 2022).
- Web of Science—Clarivate. Available online: https://www.webofscience.com/wos/woscc/basic-search (accessed on 1 October 2022).
- Bregant, C.; Mulas, A.A.; Rossetto, G.; Deidda, A.; Maddau, L.; Piras, G.; Linaldeddu, B.T. Phytophthora mediterranea sp. nov., a new species closely related to Phytophthora cinnamomi from Nursery Plants of Myrtus communis in Italy. Forests 2021, 12, 682. [Google Scholar] [CrossRef]
- Trouillas, F.P.; Nouri, M.T.; Bourret, T.B. Identification and characterization of Phytophthora species associated with crown and root rot of pistachio trees in California. Plant Dis. 2022, 106, 197–206. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Mulas, A.A.; Bregant, C.; Piras, G.; Montecchio, L. First report of Phytophthora pistaciae causing root and collar rot on nursery plants of Pistacia lentiscus in Italy. Plant Dis. 2020, 104, 1564. [Google Scholar] [CrossRef]
- Mirsoleimani, Z.; Mostowfizadeh-Ghalamfarsa, R. Characterization of Phytophthora pistaciae, the causal agent of pistachio gummosis, based on host range, morphology, and ribosomal genome. Phytopathol. Mediterr. 2013, 52, 501–516. [Google Scholar] [CrossRef]
- Mirabolfathy, M.; Cooke, D.E.L.; Duncan, J.M.; Williams, N.A.; Ershad, D.; Alizadeh, A. Phytophthora pistaciae sp. nov. and P. melonis: The principal causes of pistachio gummosis in Iran. Mycol. Res. 2001, 105, 1166–1175. [Google Scholar] [CrossRef]
- Scanu, B.; Jung, T.; Masigol, H.; Linaldeddu, B.T.; Jung, M.H.; Brandano, A.; Mostowfizadeh-Ghalamfarsa, R.; Janoušek, J.; Riolo, M.; Cacciola, S.O. Phytophthora heterospora sp. nov., a new pseudoconidia-producing sister species of P. palmivora. J. Fungi 2021, 7, 870. [Google Scholar] [CrossRef] [PubMed]
- Babacauh, K.D. Structure des populations de Phytophthora palmivora (Butl.) Butl. emend. Bras. et Griff. Parasite du Cacaoyer (Theobroma cacao L.). Bulletin Soc. Bot. Franceance. Lett. Bot. 1983, 130, 15–25. [Google Scholar] [CrossRef]
- Statista-The Statistics Portal for Market Data, Market Research and Market Studies. Available online: https://www.utu.fi/en/news/news/statista-the-statistics-portal-for-market-data-market-research-and-market-studies (accessed on 26 October 2022).
- Vitale, S.; Luongo, L.; Galli, M.; Belisario, A. First report of Phytophthora hydropathica causing wilting and shoot dieback on viburnum in Italy. Plant Dis. 2014, 98, 1582. [Google Scholar] [CrossRef]
- Faedda, R.; Cacciola, S.O.; Pane, A.; Szigethy, A.; Bakonyi, J.; Man in’t Veld, W.A.; Martini, P.; Schena, L.; di San Lio, G.M. Phytophthora × pelgrandis causes root and collar rot of Lavandula stoechas in Italy. Plant Dis. 2013, 97, 1091–1096. [Google Scholar] [CrossRef] [Green Version]
- Szigethy, A.; Nagy, Z.Á.; Vettraino, A.M.; Józsa, A.; Cacciola, S.O.; Faedda, R.; Bakonyi, J. First report of Phytophthora × pelgrandis causing root rot and lower stem necrosis of common box, lavender and port-orford-cedar in Hungary. Plant Dis. 2013, 97, 152. [Google Scholar] [CrossRef]
- Prigigallo, M.I.; Mosca, S.; Cacciola, S.O.; Cooke, D.E.L.; Schena, L. Molecular analysis of Phytophthora diversity in nursery-grown ornamental and fruit plants. Plant Pathol. 2015, 64, 1308–1319. [Google Scholar] [CrossRef] [Green Version]
- Prigigallo, M.I.; Abdelfattah, A.; Cacciola, S.O.; Faedda, R.; Sanzani, S.M.; Cooke, D.E.L.; Schena, L. Metabarcoding analysis of phytophthora diversity using genus-specific primers and 454 pyrosequencing. Phytopathology 2016, 106, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M.; Cooke, D.E.L.; Duncan, J.M.; Hansen, E.M. Multiple New phenotypic taxa from trees and riparian ecosystems in Phytophthora gonapodyides-P. megasperma ITS Clade 6, which tend to be high-temperature tolerant and either inbreeding or sterile. Mycol. Res. 2003, 107, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M. Phytophthora biodiversity: How many Phytophthora species are there? In Proceedings of the Phytophthoras in Forests and Natural Ecosystems: Fourth Meeting of the International Union of Forest Research Organizations (IUFRO) Working Party S07.02.09, Monterey, CA, USA, 26–31 August 2007. [Google Scholar]
- Davison, E.M.; Tay, F.C.S. How many soil samples are needed to show that Phytophthora is absent from sites in the South-West of Western Australia? Austral. Plant Pathol. 2005, 34, 293. [Google Scholar] [CrossRef]
- Ho, H.H. Effect of hymexazol on growth and reproduction of low temperature Phytophthora species. Mycopathologia 1987, 98, 17–20. [Google Scholar] [CrossRef]
- Burgess, T.I.; Lopez-Villamor, A.; Paap, T.; Williams, B.; Belhaj, R.; Crone, M.; Dunstan, W.; Howard, K.; Hardy, G.E.S.J. Towards a best practice methodology for the detection of Phytophthora species in soils. Plant Pathol. 2021, 70, 604–614. [Google Scholar] [CrossRef]
- Martin, F.N.; Abad, Z.G.; Balci, Y.; Ivors, K. Identification and detection of Phytophthora: Reviewing our progress, identifying our needs. Plant Dis. 2012, 96, 1080–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Hong, C. Differential usefulness of nine commonly used genetic markers for identifying Phytophthora species. Front Microbiol. 2018, 3, 2334. [Google Scholar] [CrossRef] [Green Version]
- Robideau, G.P.; De COCK, A.W.A.M.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Désaulniers, N.; Eggertson, Q.A.; Gachon, C.M.M.; et al. DNA barcoding of oomycetes with cytochrome c oxidase subunit i and internal transcribed spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef]
- Hughes, K.J.; Tomlinson, J.A.; Griffin, R.L.; Boonham, N.; Inman, A.J.; Lane, C.R. Development of a one-step real-time polymerase chain reaction assay for diagnosis of Phytophthora ramorum. Phytopathology 2006, 96, 975–981. [Google Scholar] [CrossRef] [Green Version]
- Xiaoqing, T.; Jiayi, W.; Li, M.; Yongjun, W. Detecting Phytophthora cinnamomi associated with dieback disease on Carya cathayensis using loop-mediated isothermal amplification. PLoS ONE 2021, 16, e0257785. [Google Scholar] [CrossRef]
- Zambounis, A.; Xanthopoulou, A.; Madesis, P.; Tsaftaris, A.; Vannini, A.; Bruni, N.; Tomassini, A.; Chilosi, G.; Vettraino, A.M. HRM: A tool to assess genetic diversity of Phytophthora cambivora isolates. J. Plant Pathol. 2016, 98, 611–616. [Google Scholar] [CrossRef]
- Langrell, S.R.; Morel, O.; Robin, C. Touchdown nested multiplex PCR detection of Phytophthora cinnamom and P. cambivora from French and English chestnut grove soils. Fungal Biol. 2011, 115, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zheng, Y.; Zhang, F.; Yu, J.; Dai, T.; Wang, R.; Tian, Y.; Xu, H.; Shen, D.; Dou, D. A rapid, equipment-free method for detecting phytophthora infestans in the field using a lateral flow strip-based recombinase polymerase amplification assay. Plant Dis. 2020, 104, 2774–2778. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, J.A.; Boonham, N.; Hughes, K.J.D.; Griffen, R.L.; Barker, I. On-site DNA extraction and real-time PCR for detection of Phytophthora ramorum in the field. Appl. Environ. Microbiol. 2005, 71, 6702–6710. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.G.; Miles, T.D.; Bilodeau, G.J.; Woods, P.; Blomquist, C.; Martin, F.N.; Chilvers, M.I. Validation of a preformulated, field deployable, recombinase polymerase amplification assay for Phytophthora species. Plants 2020, 9, 466. [Google Scholar] [CrossRef] [Green Version]
- Bose, T.; Wingfield, M.J.; Roux, J.; Vivas, M.; Burgess, T.I. Community composition and distribution of Phytophthora species across adjacent native and non-native forests of South Africa. Fungal Ecol. 2018, 36, 17–25. [Google Scholar] [CrossRef] [Green Version]
- La Spada, F.; Cock, P.J.A.; Randall, E.; Pane, A.; Cooke, D.E.L.; Cacciola, S.O. DNA Metabarcoding and isolation by baiting complement each other in revealing Phytophthora diversity in anthropized and natural ecosystems. J. Fungi 2022, 8, 330. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Bonants, P.; Tomassini, A.; Bruni, N.; Vannini, A. pyrosequencing as a tool for the detection of Phytophthora species: Error rate and risk of false molecular operational taxonomic units. Lett. Appl. Microbiol. 2012, 55, 390–396. [Google Scholar] [CrossRef]
- TrendEconomy Open Data Portal. Available online: https://trendeconomy.com/data (accessed on 21 October 2022).
- Franić, I.; Prospero, S.; Hartmann, M.; Allan, E.; Auger-Rozenberg, M.-A.; Grünwald, N.J.; Kenis, M.; Roques, A.; Schneider, S.; Sniezko, R.; et al. Are traded forest tree seeds a potential source of nonnative pests? Ecol. Appl. 2019, 29, e01971. [Google Scholar] [CrossRef] [Green Version]
- Cleary, M.; Oskay, F.; Doğmuş, H.T.; Lehtijärvi, A.; Woodward, S.; Vettraino, A.M. Cryptic risks to forest biosecurity associated with the global movement of commercial seed. Forests 2019, 10, 459. [Google Scholar] [CrossRef] [Green Version]
- Hulme, P.E. Unwelcome exchange: International trade as a direct and indirect driver of biological invasions worldwide. One Earth 2021, 4, n666–n679. [Google Scholar] [CrossRef]
- Garibaldi, A.; Bertetti, D.; Poli, A.; Bizioli, L.; Gullino, M.L. First report of root rot caused by Phytophthora cinnamomi on mountain laurel (Kalmia latifolia) in Italy. Plant Dis. 2012, 96, 1381. [Google Scholar] [CrossRef] [PubMed]
- Scanu, B.; Linaldeddu, B.T.; Franceschini, A.; Anselmi, N.; Vannini, A.; Vettraino, A.M. Occurrence of Phytophthora cinnamomi in cork oak forests in Italy. For. Path. 2013, 43, 340–343. [Google Scholar] [CrossRef]
- Nechwatal, J.; Jung, T. A Study of Phytophthora spp. in declining highbush blueberry in Germany reveals that P. cinnamomi can thrive under Central European outdoor conditions. J. Plant Dis. Prot. 2021, 128, 769–774. [Google Scholar] [CrossRef]
- Vettraino, A.; Potting, R.; Raposo, R. EU Legislation on forest plant health: An overview with a focus on Fusarium circinatum. Forests 2018, 9, 568. [Google Scholar] [CrossRef] [Green Version]
- Engler, A.; Nahuelhual, L.; Cofré, G.; Barrena, J. How far from harmonization are sanitary, phytosanitary and quality-related standards? An exporter’s perception approach. Food Policy 2012, 37, 162–170. [Google Scholar] [CrossRef]
- Brasier, C.M. Preventing invasive pathogens: Deficiencies in the system. Plantsman 2005, 4, 54–57. [Google Scholar]
- Day, R.K. More trade, safer trade: Strengthening developing countries’ sanitary and phytosanitary (SPS) capacity. CABI Work. Paper 2013, 4, 33. [Google Scholar]
- Kumar, P.L.; Cuervo, M.; Kreuze, J.F.; Muller, G.; Kulkarni, G.; Kumari, S.G.; Massart, S.; Mezzalama, M.; Alakonya, A.; Muchugi, A.; et al. Phytosanitary interventions for safe global germplasm exchange and the prevention of transboundary pest spread: The role of CGIAR Germplasm Health Units. Plants 2021, 10, 328. [Google Scholar] [CrossRef]
- Roy, H.E.; Rabitsch, W.; Scalera, R.; Stewart, A.; Gallardo, B.; Genovesi, P.; Essl, F.; Adriaens, T.; Bacher, S.; Booy, O.; et al. Developing a framework of minimum standards for the risk assessment of alien species. J. Appl. Ecol. 2018, 55, 526–538. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Plant Health. Guidance on the environmental risk assessment of plant pests. EFSA J. 2011, 9, 2460. [Google Scholar] [CrossRef]
- Kenis, M.; Bacher, S.; Baker, R.H.A.; Branquart, E.; Brunel, S.; Holt, J.; Hulme, P.E.; MacLeod, A.; Pergl, J.; Petter, F.; et al. New protocols to assess the environmental impact of pests in the EPPO decision-support scheme for pest risk analysis. Bulletin OEPP EPPO Bull. Eur. Mediterr. Plant Prot. Organ. 2012, 42, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Vettraino, A.M.; Roques, A.; Yart, A.; Fan, J.T.; Sun, J.H.; Vannini, A. Sentinel trees as a tool to forecast invasions of alien plant pathogens. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef]
- Casarin, N.; Hasbroucq, S.; López-Mercadal, J.; Ángel Miranda, M.; Bragard, C.; Grégoire, J.-C. Measuring the threat from a distance: Insight into the complexity and perspectives for implementing sentinel plantation to test host range of Xylella fastidiosa. bioRiv 2022. bioRiv:07.22.500186. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Santini, A.; Nikolov, C.; Grégoire, J.C.; Tomov, R.; Orlinski, A.; Maaten, T.; Sverrisson, H.; Økland, B.; Eschen, R. A worldwide perspective of the legislation and regulations governing sentinel plants. Biol. Invasions 2020, 22, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Parke, J.L.; Grünwald, N.J. A systems approach for management of pests and pathogens of nursery crops. Plant Dis. 2012, 96, 1236–1244. [Google Scholar] [CrossRef] [Green Version]
- Weiland, J.E. The Challenges of Managing Phytophthora Root Rot in the Nursery Industry. Plant Health Prog. 2021, 22, 332–341. [Google Scholar] [CrossRef]
- Kutawa, A.B.; Ahmad, K.; Ali, A.; Hussein, M.Z.; Abdul Wahab, M.A.; Adamu, A.; Ismaila, A.A.; Gunasena, M.T.; Rahman, M.Z.; Hossain, M.I. Trends in nanotechnology and its potentialities to control plant pathogenic fungi: A review. Biology 2021, 10, 881. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Zikeli, F.; Scarascia Mugnozza, G.; Vinciguerra, V.; Tabet, D.; Romagnoli, M. Lignin nanoparticles containing essential oils for controlling Phytophthora cactorum diseases. Forest Pathol. 2022, 52, e12739. [Google Scholar] [CrossRef]
- Abd El Aty, A.A.; Zohair, M.M. Green-synthesis and optimization of an eco-friendly nanobiofungicide from Bacillus amyloliquefaciens MH046937 with antimicrobial potential against phytopathogens. Environ. Nanotechnol. Monit. Manag. 2020, 14, 100309. [Google Scholar] [CrossRef]
- Ahmad, H.; Venugopal, K.; Bhat, A.H.; Kavitha, K.; Ramanan, A.; Rajagopal, K.; Srinivasan, R.; Manikandan, E. Enhanced biosynthesis synthesis of copper oxide nanoparticles (CuO-NPs) for their antifungal activity toxicity against major phyto-pathogens of apple orchards. Pharm Res. 2020, 37, 246. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonelli, C.; Biscontri, M.; Tabet, D.; Vettraino, A.M. The Never-Ending Presence of Phytophthora Species in Italian Nurseries. Pathogens 2023, 12, 15. https://doi.org/10.3390/pathogens12010015
Antonelli C, Biscontri M, Tabet D, Vettraino AM. The Never-Ending Presence of Phytophthora Species in Italian Nurseries. Pathogens. 2023; 12(1):15. https://doi.org/10.3390/pathogens12010015
Chicago/Turabian StyleAntonelli, Chiara, Margherita Biscontri, Dania Tabet, and Anna Maria Vettraino. 2023. "The Never-Ending Presence of Phytophthora Species in Italian Nurseries" Pathogens 12, no. 1: 15. https://doi.org/10.3390/pathogens12010015