

The Investigation of Giardiasis (Foodborne and Waterborne Diseases) in Buffaloes in Van Region, Türkiye: First Molecular Report of Giardia duodenalis Assemblage B from Buffaloes
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Area
2.2. Animal Material and Sample Collection
2.3. Method
2.3.1. Microscopic Analysis
2.3.2. DNA Extraction and Nested PCR
2.3.3. Sequence Analyzes
3. Results
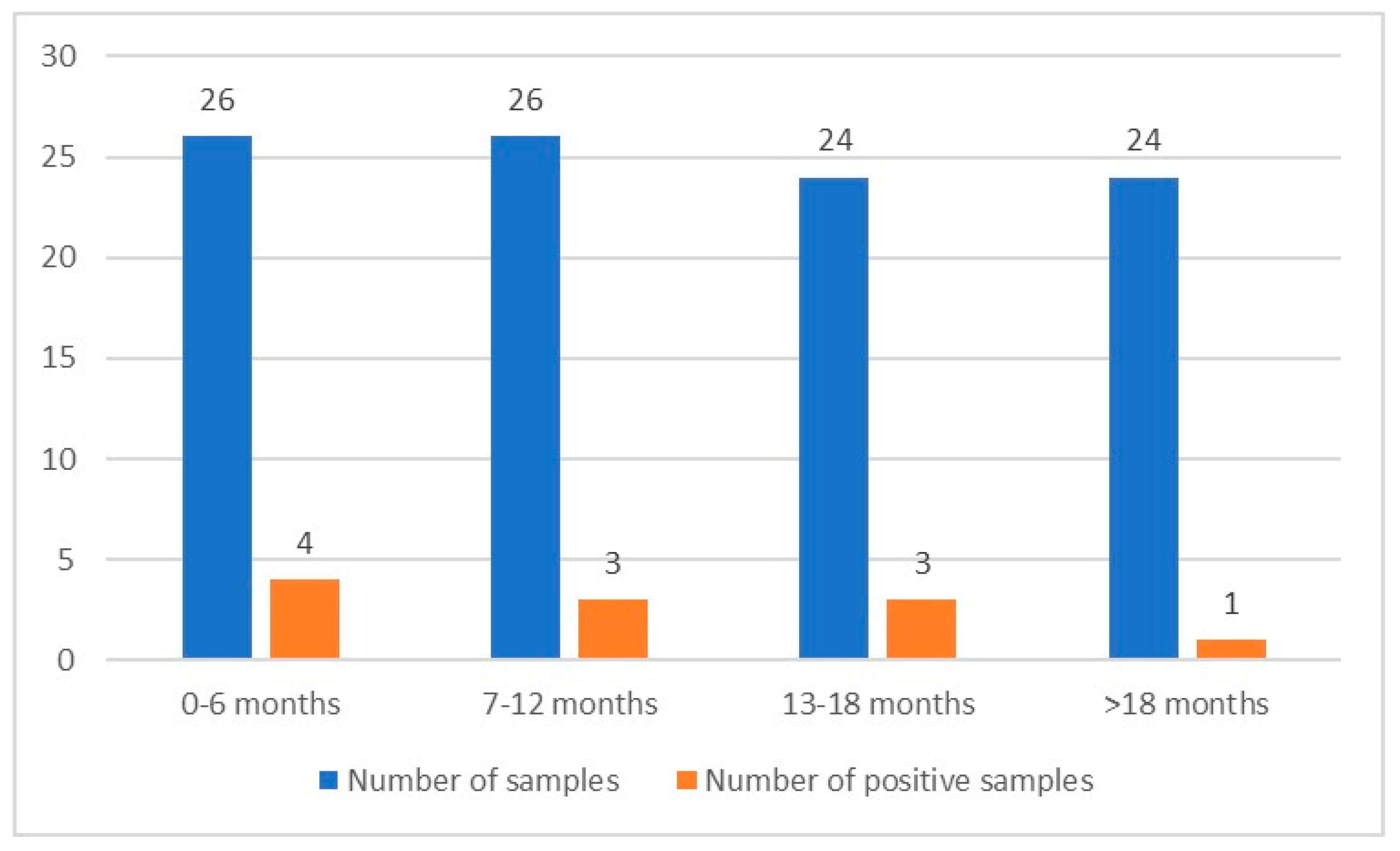
3.1. Native Lugol and Nested PCR Analysis Results
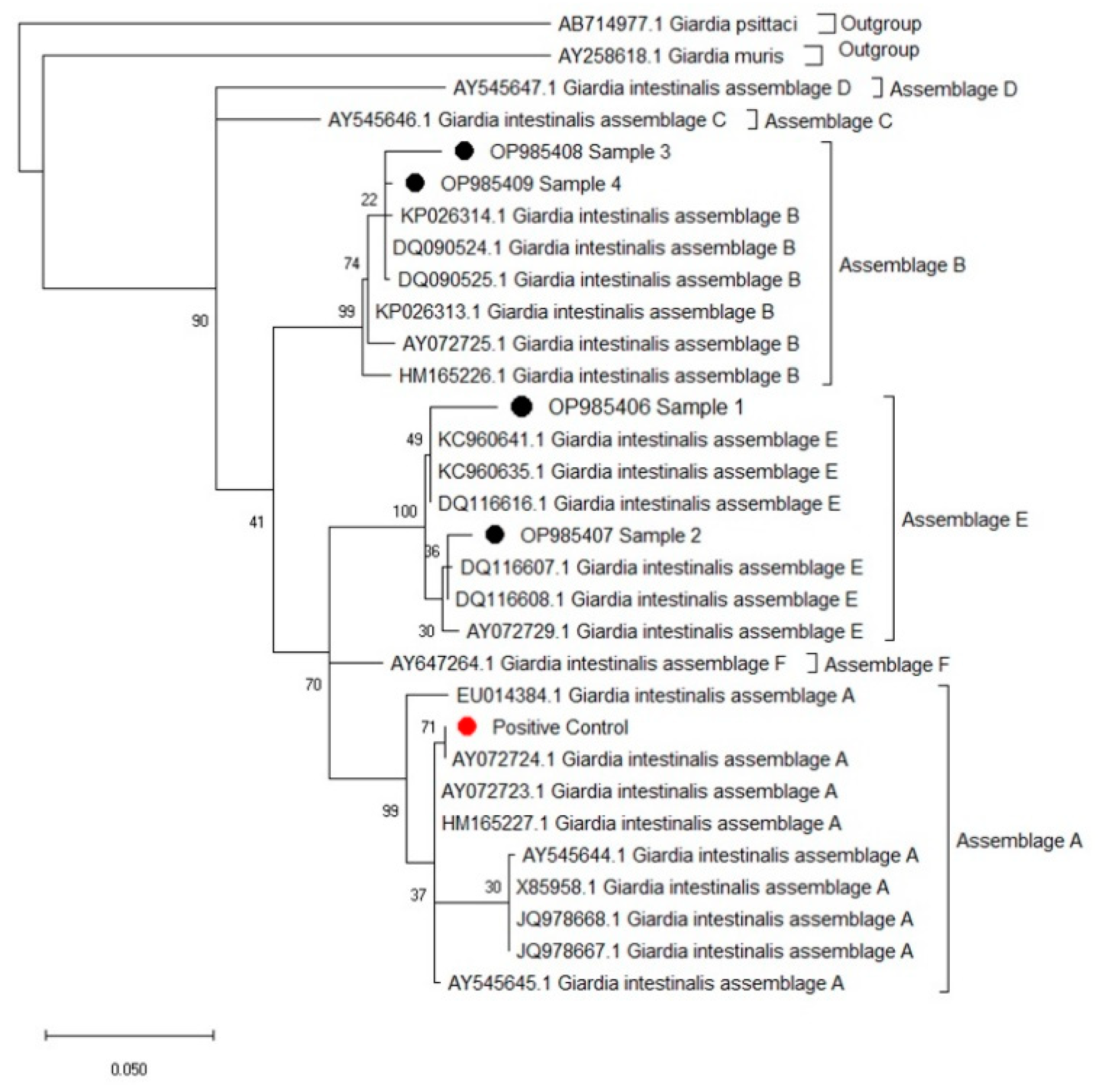
3.2. Sequence Analyses and NCBI Basic Local Alignment Search
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Estimates of Foodborne Ilness in the United States. Available online: //www.cdc.gov/foodborneburden/2011-foodborne-estimates.html. (accessed on 11 October 2022).
- WHO. Estimating The Burden of Foodborne Diseases. Available online: https://www.who.int/activities/estimating-the-burden-of-foodborne-diseases (accessed on 11 October 2022).
- Ansell, B.R.E.; McConville, M.J.; Ma’ayeh, S.Y.; Dagley, M.J.; Gasser, R.B.; Svärd, S.G.; Jex, A.R. Drug resistance in Giardia duodenalis. Biotechnol. Adv. 2015, 33, 888–901. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Hijjawi, N.; Feng, Y.; Xiao, L. Giardia: An Under-Reported Foodborne Parasite. Int. J. Parasitol 2019, 49, 1–11. [Google Scholar] [CrossRef] [PubMed]
- WHO. World Health Statistics; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- FAO/WHO. Multicriteria-based ranking for risk management of food-borne parasites. Microbiological Risk Assessment Series No. 23. Food and Agriculture Organization of the United Nations/World Health Organization, 2014. Available online: http://apps.who.int/iris/bitstream/10665/112672/1/9789241564700_eng.pdf (accessed on 27 September 2022).
- Robertson, L.J. Giardia as a Foodborne Pathogen. In Springer Briefs in Food, Health and Nutrition; Springer: New York, NY, USA; Heidelberg, Germany; Dordrecht, The Netherlands; London, UK, 2013. [Google Scholar]
- Dixon, B.R. Transmission dynamics of foodborne parasites on fresh produce. In Foodborne Parasites in the Food Supply Web; Gajadhar, A.A., Ed.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Oxford UK, 2015; pp. 317–353. [Google Scholar]
- Squire, S.A.; Yang, R.; Robertson, I.; Ayi, I.; Ryan, U. Molecular characterization of Cryptosporidium and Giardia in farmers and their ruminant livestock from the Coastal Savannah zone of Ghana. Infect. Genet. Evol. 2017, 55, 236–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillin, F.D.; Eckmann, L. Encyclopedia of Gastroenterology; Johnson, L.R., Ed.; Elsevier: London, UK, 2003; Chapter 13; pp. 233–234. [Google Scholar]
- Robertson, L.J. Giardia and Cryptosporidium infections in sheep and goats: A review of the potential for transmission to humans via environmental contamination. Epidemiol. Infect. 2009, 137, 913–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efstratiou, A.; Ongerth, J.; Karanis, P. Evolution of monitoring for Giardia and Cryptosporidium in water. Water Res. 2017, 123, 96–112. [Google Scholar] [CrossRef] [PubMed]
- Fantinatti, M.; Bello, A.R.; Fernandes, O.; Da-Cruz, A.M. Identification of Giardia lamblia assemblage E in humans points to a new anthropozoonotic cycle. J. Infect. Dis. 2016, 214, 1256–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, M.E.; O’Handley, R.M.; Ralston, B.J.; McAllister, T.A.; Thompson, R.A. Update on Cryptosporidium and Giardia infections in cattle. Trends Parasitol. 2004, 20, 185–191. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, G.; Chen, G.; Jian, F.; Zhang, S.; Feng, C.; Wang, R.; Zhu, J.; Dong, H.; Hua, J. Multilocus genotyping of Giardia duodenalis in dairy cattle in Henan, China. PLoS ONE 2014, 9, e100453. [Google Scholar] [CrossRef] [Green Version]
- Jian, Y.; Zhang, X.; Li, X.; Karanis, G.; Ma, L.; Karanis, P. Prevalence and molecular characterization of Giardia duodenalis in cattle and sheep from the Qinghai-Tibetan Plateau Area (QTPA), northwestern China. Vet. Parasitol. 2018, 250, 40–44. [Google Scholar] [CrossRef]
- Soysal, M. Anatolian Water Buffaloes Husbandry in Turkey. Buffalo Bull. 2013, 32, 293–309. [Google Scholar]
- Cacciò, S.M.; Rinaldi, L.; Cringoli, G.; Condoleo, R.; Pozio, E. Molecular identification of Cryptosporidium parvum and Giardia duodenalis in the Italian water buffalo (Bubalus bubalis). Vet. Parasitol. 2007, 150, 146–149. [Google Scholar] [CrossRef]
- Abeywardena, H.; Jex, A.R.; Koehler, A.V.; Rajapakse, R.; Udayawarna, K.; Haydon, S.R.; Stevens, M.A.; Gasser, R.B. First molecular characterization of Cryptosporidium and Giardia from bovines (Bos taurus and Bubalus bubalis) in Sri Lanka: Unexpected absence of C. parvum from pre-weaned calves. Parasites Vectors 2014, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Helmy, Y.A.; Klotz, C.; Wilking, H.; Krücken, J.; Nöckler, K.; Samson-Himmelstjerna, V.; Zessin, K.-H.; Aebischer, T. Epidemiology of Giardia duodenalis infection in ruminant livestock and children in the Ismailia province of Egypt: Insights by genetic characterization. Parasites Vectors 2014, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Caccio, S.; De Giacomo, M.; Pozio, E. Sequence analysis of the β-Giardin gene and development of a PCR-RFLP assay to genotype Giardia duodenalis cysts from human faecal samples. Int. J. Parasitol. 2002, 32, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Lalle, M.; Pozio, E.; Capelli, G.; Bruschi, F.; Crotti, D.; Cacciò, S.M. Genetic heterogeneity at the β-giardin locus among human and animal isolates of Giardia duodenalis and identification of potentially zoonotic subgenotypes. Int. J. Parasitol. 2005, 35, 207–213. [Google Scholar] [CrossRef]
- Ayan, A.; Ural, D.A.; Erdogan, H.; Kilinc, O.O.; Gültekin, M.; Ural, K. Prevalance and molecular characterization of Giardia duodenalis in livestock in Van, Turkey. Int. J. Ecosyst. Ecol. Sci. 2019, 9, 289–296. [Google Scholar] [CrossRef]
- Siwila, J. Giardiasis: Livestock and companion animals. Curr. Top. Giardiasis 2017, 39–49. [Google Scholar]
- Feng, Y.; Xiao, L. Zoonotic potential and molecular epidemiology of Giardia species and giardiasis. Clin. Microbiol. Rev. 2011, 24, 110–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mark-Carew, M.P.; Wade, S.E.; Chang, Y.-F.; Schaaf, S.; Mohammed, H.O. Prevalence of Giardia duodenalis assemblages among dairy herds in the New York City Watershed. Vet. Parasitol. 2012, 185, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.; Power, M.; Ens, E. Cryptosporidium and Giardia in feral water buffalo (Bubalus bubalis) in the South East arnhem land indigenous protected area, Australia. Parasitol. Res. 2020, 119, 2149–2157. [Google Scholar] [CrossRef]
- de Aquino, M.C.C.; Harvey, T.V.; Inácio, S.V.; Nagata, W.B.; Ferrari, E.D.; Oliveira, B.C.M.; Albuquerque, G.R.; Widmer, G.; Meireles, M.V.; Bresciani, K.D.S. First description of Giardia duodenalis in buffalo calves (Bubalus bubalis) in southwest region of São Paulo State, Brazil. Food Waterborne Parasitol. 2019, 16, e00062. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, L.; Condoleo, R.; Condoleo, R.; Saralli, G.; Bruni, G.; Cringoli, G. Cryptosporidium and Giardia in water buffaloes (Bubalus bubalis) of the Italian Mediterranean bred. Vet. Res. Commun. 2007, 31, 253–255. [Google Scholar] [CrossRef]
- Obayes, H.H.; Al-Rubaie, H.M.; Zayer, A.A.J.; Radhy, A.M. Detection of the prevalence of some gastrointestinal protozoa in buffaloes of Babylon Governorate. Basrah J. Vet. Res. 2016, 15, 294–303. [Google Scholar]
- Ozdal, N.; Tanritanir, P.; Göz, Y.; Deger, S.; Kozat, S. Parasitic Protozoans (Eimeria, Giardia, and Cryptosporidium) in lambs. Bull Vet Inst Pulawy 2009, 53, 47–51. [Google Scholar]
- Değerli, S.; Çeliksöz, A.; Kalkan, K.; Özçelik, S. Prevalence of Cryptosporidium spp. and Giardia spp. in cows and calves in Sivas. Turk. J. Vet. Anim. Sci. 2005, 29, 995–999. [Google Scholar]
- Ayan, A.; Ural, K.; Aysul, N.; Gültekin, M.; Erdoğan, H.; Balikçi, C.; Toplu, S.; Toros, G. Natural Cyst Shedding in Calves Infected with Giardia duodenalis. J. Adv. VetBio Sci. Tech. 2016, 1, 14–19. [Google Scholar]
- Abeywardena, H.; Jex, A.R.; von Samson-Himmelstjerna, G.; Haydon, S.R.; Stevens, M.A.; Gasser, R.B. First molecular characterisation of Cryptosporidium and Giardia from Bubalus bubalis (water buffalo) in Victoria, Australia. Infect. Genet. Evol. 2013, 20, 96–102. [Google Scholar] [CrossRef]
- Utaaker, K.S.; Chaudhary, S.; Bajwa, R.S.; Robertson, L.J. Prevalence and zoonotic potential of intestinal protozoans in bovines in Northern India. Vet. Parasitol. Reg. Stud. Rep. 2018, 13, 92–97. [Google Scholar] [CrossRef]
- Taş, C.Z.; Yılmaz, H.; Beyhan, B.Y.; Çiçek, M. A Comprehensive Retrospective Study: Intestinal Parasites in Human in Van Province. Türkiye Parasitol. J. 2019, 43, 70–73. [Google Scholar]
- Khurajog, B.; Masakul, A.; Inpankaew, T.; Kamyingkird, K.; Wongpanit, K.; Jittapalapong, S. Prevalence of Giardia duodenalis and factors associated with its infection in water buffaloes in Northeast Thailand. J. Trop. Med. Parasitol. 2014, 37, 35–41. [Google Scholar]
- Bărburaș, D.A.; Cozma, V.; Ionică, A.M.; Abbas, I.; Bărburaș, R.; Mircean, V.; D’Amico, G.; Dubey, J.P.; Gyorke, A. Intestinal Parasites of Buffalo Calves from Romania: Molecular Characterization of Cryptoporidium spp. and Giardia duodenalis, and the first Report of Eimeria Bareillyi. Folia Parasitol. 2021, 8, 69. [Google Scholar] [CrossRef]
- Aydin, A.F.; Besirbellioglu, B.A.; Avci, I.Y.; Tanyuksel, M.; Araz, E.; Pahsa, A. Classification of Giardia duodenalis parasites in Turkey into groups A and B using restriction fragment length polymorphism. Diagn Microbiol. Infect. Dis. 2004, 50, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Balcioglu, C.; Kurt, O.; Sevil, N.; Dagcı, H.; Tetik, A.; Ergunay, K. Genotyping of Giardia lamblia in a cohort of Turkish patients: A search for a relationship between symptoms and genotypes. Kafkas Univ. Vet. Fak. Derg. 2012, 18 (Suppl. A), A125–A131. [Google Scholar] [CrossRef]
- Değerli, S.; Değerli, N.; Çeliksöz, A.; Özçelik, S. Genotyping of Giardia intestinalis isolated from people living in Sivas, Turkey. Turk J. Med. Sci. 2012, 42, 1268–1272. [Google Scholar] [CrossRef]
- Tamer, G.S.; Kasap, M.; Er, D.K. Genotyping and phylogenetic analysis of Giardia duodenalis isolates from Turkish children. Med. Sci. Monit. 2015, 21, 526–532. [Google Scholar] [PubMed] [Green Version]
- Ertug, S.; Ertabaklar, H.; Ozlem Caliskan, S.; Malatyali, E.; Bozdoğan, B. Genotyping of Giardia intestinalis strains isolated from humans in Aydin, Turkey. Mikrobiyol. Bul. 2016, 50, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Koloren, Z.; Seferoğlu, O.; Karanis, P. Occurrency of Giardia duodenalis assemblages in river water sources of Black Sea, Turkey. Acta Trop. 2016, 164, 337–344. [Google Scholar] [CrossRef]
- Sursal, N.; Simsek, E.; Yildiz, K. Feline giardiasis in Turkey: Prevalence and genetic and haplotype diversity of Giardia duoedenalis based on the β-Giardin gene sequence in symptomatic cats. J. Parasitol. 2020, 106, 699–706. [Google Scholar] [CrossRef]
- Homan, W.L.; Mank, T.G. Human giardiasis: Genotype linked differences in clinical symptomatology. Int. J. Parasitol. 2001, 31, 822–826. [Google Scholar] [CrossRef]
- Mahdy, A.K.M.; Surin, J.; Wan, K.L.; Mohd-Adnan, A.; Al-Mekhlafi, M.S.H.; Lim, Y.A.L. Giardia intestinalis genotypes: Risk factors and correlation with clinical symptoms. Acta Trop. 2009, 12, 67–70. [Google Scholar] [CrossRef]
- Heyworth, M.F. Giardia duodenalis genetic assemblages and hosts. Parasite 2016, 23, 13. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sequence Analyzed Samples and Accession Numbers | Age | Access Codes of the Most Similar Example and Similarity Rate | |
|---|---|---|---|
| Assemblage B (MG736344) | Assemblage E (KT922248) | ||
| Sample 1 (OP985406) | 4 months | 93.95% | 99.34% |
| Sample 2 (OP985407) | 4 months | 94.19% | 99.57% |
| Sample 3 (OP985408) | 9 months | 98.85% | 93.35% |
| Sample 4 (OP985409) | 3 months | 99.58% | 94.80% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kılınç, Ö.O.; Ayan, A.; Çelik, B.A.; Çelik, Ö.Y.; Yüksek, N.; Akyıldız, G.; Oğuz, F.E. The Investigation of Giardiasis (Foodborne and Waterborne Diseases) in Buffaloes in Van Region, Türkiye: First Molecular Report of Giardia duodenalis Assemblage B from Buffaloes. Pathogens 2023, 12, 106. https://doi.org/10.3390/pathogens12010106
Kılınç ÖO, Ayan A, Çelik BA, Çelik ÖY, Yüksek N, Akyıldız G, Oğuz FE. The Investigation of Giardiasis (Foodborne and Waterborne Diseases) in Buffaloes in Van Region, Türkiye: First Molecular Report of Giardia duodenalis Assemblage B from Buffaloes. Pathogens. 2023; 12(1):106. https://doi.org/10.3390/pathogens12010106
Chicago/Turabian StyleKılınç, Özlem Orunç, Adnan Ayan, Burçak Aslan Çelik, Özgür Yaşar Çelik, Nazmi Yüksek, Gürkan Akyıldız, and Fatma Ertaş Oğuz. 2023. "The Investigation of Giardiasis (Foodborne and Waterborne Diseases) in Buffaloes in Van Region, Türkiye: First Molecular Report of Giardia duodenalis Assemblage B from Buffaloes" Pathogens 12, no. 1: 106. https://doi.org/10.3390/pathogens12010106