

Emergent and Neglected Equine Filariosis in Egypt: Species Diversity and Host Immune Response
 , and
, and
Abstract
:1. Introduction
2. Results
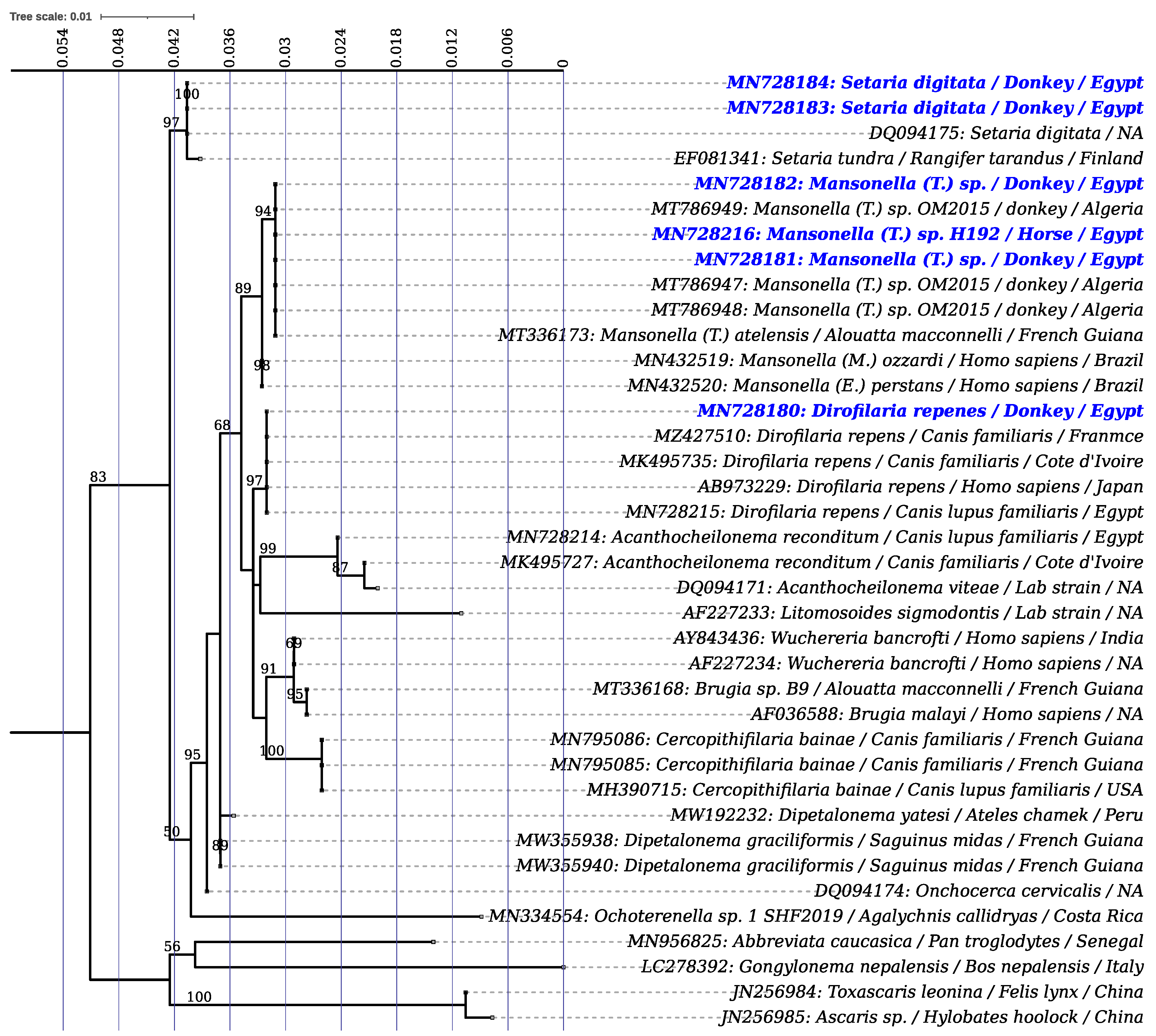
2.1. Prevalence and Phylogeny of Filaroids
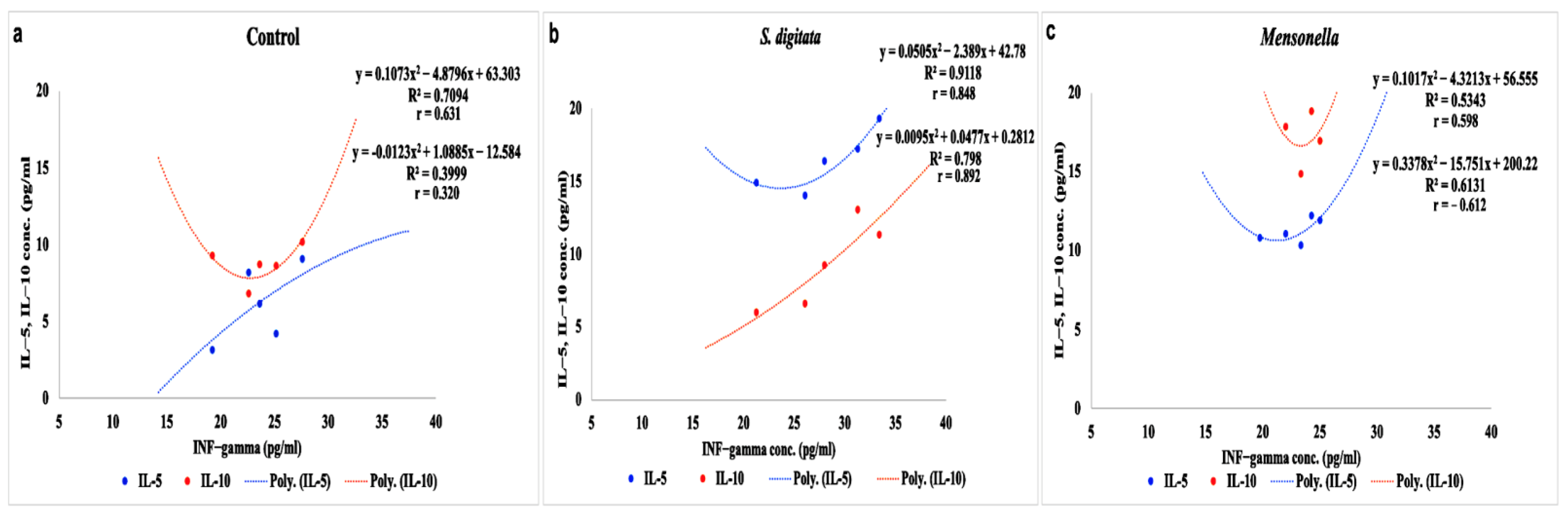
2.2. Immunological Studies
3. Discussion
4. Materials and Methods
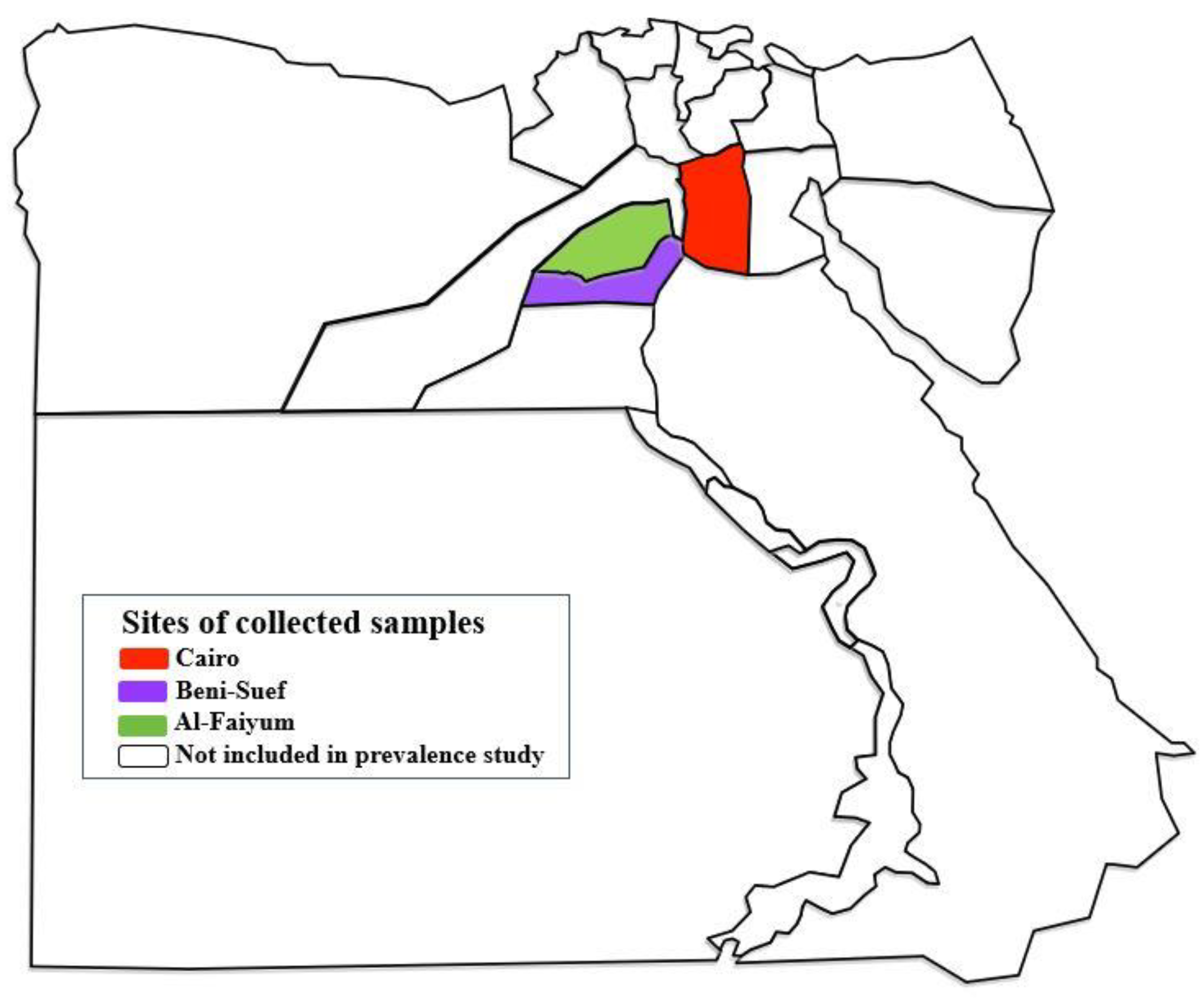
4.1. Study Design and Animal Sampling
4.2. Molecular Studies
4.2.1. Polymerase Chain Reactions and Sequencing
4.2.2. Phylogenetic Analyses
4.3. Immunological Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taylor, L.H.; Latham, S.M.; Woolhouse, M.E.J. Risk factors for human disease emergence. Philos. Trans. R. Soc. Lond. B 2001, 356, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Perumal, A.N.I.; Gunawardene, Y.I.N.S.; Dassanayake, R.S. Setaria digitata in advancing our knowledge of human lymphatic filariasis. J. Helminthol. 2016, 90, 129–138. [Google Scholar] [CrossRef]
- Nabie, R.; Spotin, A.; Rouhani, S. Subconjunctivalsetariasis due to Setaria equina infection; A case report and a literature review. Parasitol. Int. 2016, 66, 930–932. [Google Scholar] [CrossRef] [PubMed]
- Laidoudi, Y.; Medkour, H.; Levasseur, A.; Davoust, B.; Mediannikov, O. New Molecular Data on Filaria and its Wolbachia from Red Howler Monkeys (Alouattamacconnelli) in French Guiana-A Preliminary Study. Pathogens 2020, 9, 626. [Google Scholar] [CrossRef] [PubMed]
- Khamesipour, F.; Taktaz-Hafshejani, T.; Tebit, K.E.; Razavi, S.M.; Hosseini, S.R. Prevalence of endo- and ecto-parasites of equines in Iran: A systematic review. Vet. Med. 2021, 7, 25–34. [Google Scholar] [CrossRef] [PubMed]
- El Namaky, A.H.; Hendawy, S.H.M.; Abo-Aziza, F.A.M.; Ashry, H.M. Cytokines and immunoglobulin G response in donkeys with spontaneous Setaria equina infection. Bulg. J. Vet. Med. 2019, 22, 180–189. [Google Scholar] [CrossRef]
- Radwan, A.M.; Ahmed, N.E.; Elakabawy, L.M.; Ramadan, M.Y.; Elmadawy, R.S. Prevalence and pathogenesis of some filarial nematodes infecting donkeys in Egypt. Vet. World 2016, 9, 888–892. [Google Scholar] [CrossRef]
- Peng, T.L.; Armiladiana, M.M.; Ruhil, H.H.; Maizan, M.; Choong, S.S. First report of equine Setaria digitata (von Linstow 1906) infestation in Malaysia. Vet. Parasitol. Reg. Stud. Rep. 2019, 17, 100310. [Google Scholar] [CrossRef]
- Yu, F.; Liu, B.; Chen, S.; Yi, Z.; Liu, X.; Zhu, Y.; Li, J. First Molecular Confirmation of Equine Ocular Setaria digitata in China. Vet. Sci. 2021, 8, 55. [Google Scholar] [CrossRef]
- Tamilmahan, P.; Zama, M.M.S.; Pathak, R.; Muneeswaran, N.S.; Karthik, K. A retrospective study of ocular occurrence in domestic animals: 799 cases. Vet. World. 2013, 6, 274–276. [Google Scholar] [CrossRef]
- Hillyer, L.; Coles, G.; Randle, T. Setaria equina in the UK. Vet. Rec. 2001, 149, 464. [Google Scholar] [PubMed]
- Abbas, I.; Al-Araby, M.; Al-Kappany, Y. Molecular characterization of Setaria equina infecting donkeys (Equusasinus) from Egypt. Res. J. Parasitol. 2016, 11, 73–78. [Google Scholar] [CrossRef]
- Maharana, B.R.; Potliya, S.; Ganguly, A.; Bisla, R.S.; Mishra, C.; Ganguly, I. First report of the isolation and phylogenetic characterization of equine Setaria digitata from India based on mitochondrial COI, 12S rDNA, and nuclear ITS2 sequence data. Parasitol. Res. 2020, 119, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Laidoudi, Y.; Bedjaoui, S.; Medkour, H.; Latrofa, M.S.; Mekroud, A.; Bitam, I.; Davoust, B.; Otranto, D.; Mediannikov, O. Molecular Approach for the Diagnosis of Blood and Skin Canine Filarioids. Microorganisms 2020, 8, 1671. [Google Scholar] [CrossRef] [PubMed]
- Harnett, W.; Harnett, M.M. Filarial nematode secreted product ES-62 is an anti-inflammatory agent: Therapeutic potential of small molecule derivatives and ES-62 peptide mimetics. Clin. Exp. Pharmacol. Physiol. 2006, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Abo-Aziza, F.A.M.; Hendawy, S.H.M.; El Namaky, A.H.; Ashry, H.M. Th1/Th2 balance and humoral immune response to potential antigens as early diagnostic method of equine Strongylus nematode infection. Vet. World 2017, 10, 679–687. [Google Scholar] [CrossRef]
- Hendawy, S.H.M. Immunity to gastrointestinal nematodes in ruminants: Effector cell mechanisms and cytokines. J. Parasit. Dis. 2018, 42, 471–482. [Google Scholar] [CrossRef]
- Babu, S.; Nutman, T. Immunology of lymphatic filariasis. Parasite Immunol. 2013, 36, 2. [Google Scholar] [CrossRef]
- Specht, S.; Volkmann, L.; Wynn, T.; Hoerauf, A. Interleukin-10 (IL-10) counter regulates IL-4- dependent effector mechanisms in murine filariasis. Infect. Immun. 2004, 72, 6287–6293. [Google Scholar] [CrossRef]
- Metenou, S.; Dembe, B.; Konate, S.; Dolo, H.; Coulibaly, S.Y.; Coulibaly, Y.I.; Diallo, A.A.; Soumaoro, L.; Coulibaly, M.E.; Sanogo, D.; et al. Patent filarial infection modulates malaria-specific type 1 cytokine responses in an IL-10-dependent manner in a filaria/malaria-coinfected population. J. Immunol. 2009, 183, 916–924. [Google Scholar] [CrossRef] [Green Version]
- Schierack, P.; Lucius, R.; Sonnenburg, B.; Schilling, K.; Hartmann, S. Parasite-specific immunomodulatory functions of filarial cystatin. Infect. Immun. 2003, 71, 2422–2429. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.; Eleftherianos, I. Parasitic nematode immunomodulatory strategies: Recent advances and perspectives. Pathogens 2016, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, R.; George, P.J.; Hanna, L.E.; Kumaran, P.; Chandrasekaran, V.; Nutman, T.B.; Babu, S. Expansion of parasite-specific CD4+ and CD8+ T cells expressing IL-10 superfamily cytokine members and their regulation in human lymphatic filariasis. PLoS Negl. Trop. Dis. 2014, 8, e0002762. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.C.; Lindberg, A.C.; Main, D.C.; Whay, H.R. Assessment of the welfare of working horses, mules and donkeys, using health and behaviour parameters. Prev. Vet. Med. 2005, 69, 265–283. [Google Scholar] [CrossRef]
- Valette, D. Invisible Workers. The Economic Contributions of Working Donkeys, Horses and Mules to Livelihoods; The Brooke: Louisville, KY, USA, 2015; pp. 1–23. [Google Scholar]
- Abu El-Magd, A.; Ahmed, Z.G. The occurrence of Setaria equina in donkey’s eyes and their treatment. Assiut Vet. Med. J. 1994, 31, 86–90. [Google Scholar]
- Abd El-Wahab, T.M.; Ashour, A.A. Scanning electron microscopy of the two filariid nematodes Setaria equina and Onchocerca cervicalis from Kafr El-Sheikh area, Egypt. Alex. J. Vet. Sci. 1999, 15, 541–547. [Google Scholar] [CrossRef]
- Mahmoud, A.E. Laboratory Diagnosis of Filariasis in Assiut Governorate. Master’s Thesis, Faculty of Veterinary Medicine, University of Assiut, Assiut, Egypt, 1998. [Google Scholar]
- Mahmoud, A.E. A study of Setaria equina (Abildgaard, 1789) by light and scanning electron microscopy. EL-Minia Med. Bullet. 2006, 17, 157–173. [Google Scholar]
- Marzok, M.A.; Desouky, A.Y. Ocular infection of donkeys (Equusasinus) with Setaria equina. Trop. Anim. Health Prod. 2009, 41, 859–863. [Google Scholar] [CrossRef]
- Ahmed, N.E.; El-Akabawy, L.M.; Ramadan, M.Y.; Radwan, A.M.M. Studies on helminth parasites in necropsied donkeys in Egypt. Benha Vet. Med. J. 2011, 1, 153–162. [Google Scholar]
- Abdel Rahman, M.M.I. Morphological and molecular characterization of Setaria equina in donkeys. Beni-Suef Univ. J. Basic Appl. Sci. 2020, 9, 17. [Google Scholar] [CrossRef]
- World Health Organization. Global programme to eliminate lymphatic filariasis: Annual report on lymphatic filariasis. Wkly. Epidemiol. Record. 2006, 82, 361–380. [Google Scholar]
- Devi, A.; Sudan, V.; Shanker, D. Phylogenetic characterization of Setaria equina and its association with other filarids. Parasitol. Res. 2020, 119, 4267–4270. [Google Scholar] [CrossRef] [PubMed]
- Nagaty, H.F. Parlitomosazakii (Filariinae), a new genus and species and its microfilaria from Leontocebusrosalia. J. Egypt. Med. Assoc. 1935, 18, 483–496. [Google Scholar]
- Eberhard, M.L.; Orihel, T.C. The genus Mansonella (syn. Tetrapetalonema): A new classification. Ann. Parasitol. Hum. Comp. 1984, 59, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Sandground, J.H. Report on the nematode parasites collected by the Kelley-Roosevelts expedition to Indo-China with descriptions of several new species. Part 1. Parasites of birds. Part 2. Parasites of mammals. Z. Parasitenkd. 1933, 5, 542–583. [Google Scholar] [CrossRef]
- Wijesundera, W.S.; Chandrasekharan, N.V.; Karunanayake, E.H. A sensitive polymerase chain reaction-based assay for the detection of Setaria digitata: The causative organism of cerebrospinal nematodiasis in goats, sheep and horses. Vet. Parasitol. 1999, 81, 225–233. [Google Scholar] [CrossRef]
- BinoSundar, S.T.; D’Souza, P.E. Morphological characterization of Setaria worms collected from cattle. J. Parasit. Dis. 2015, 39, 572–576. [Google Scholar] [CrossRef]
- Kaur, D.; Ganai, A.; Parveen, S.; Borkataki, S.; Yadav, A.; Katoch, R.; Godara, R. Occurrence of Setaria digitata in a cow. J. Parasit. Dis. 2015, 39, 477–478. [Google Scholar] [CrossRef]
- Abdullah, H.H.A.M.; Amanzougaghene, N.; Dahmana, H.; Louni, M.; Raoult, D.; Mediannikov, O. Multiple vector-borne pathogens of domestic animals in Egypt. PLoS Negl. Trop. Dis. 2021, 15, e0009767. [Google Scholar] [CrossRef]
- Shin, J.; Ahn, K.S.; Suh, G.H.; Kim, H.J.; Jeong, H.S.; Kim, B.S.; Choi, E.; Shin, S.S. First Blindness Cases of Horses Infected with Setaria Digitata (Nematoda: Filarioidea) in the Republic of Korea. Korean J. Parasitol. 2017, 55, 667–671. [Google Scholar] [CrossRef]
- Lee, H.; Hwang, H.; Ro, Y.; Kim, J.; Lee, K.; Choi, E.; Bae, Y.; So, B.; Lee, I. Setaria digitata was the main cause of equine neurological ataxia in Korea: 50 case s (2015–2016). J. Vet. Med. Sci. 2021, 83, 869–875. [Google Scholar] [CrossRef]
- Genchi, C.; Kramer, L. Subcutaneous dirofilariosis (Dirofilaria repens): An infection spreading throughout the old world. Parasit. Vectors 2017, 10, 517. [Google Scholar] [CrossRef]
- Laidoudi, Y.; Davoust, B.; Varloud, M.; Niang, E.H.A.; Fenollar, F.; Mediannikov, O. Development of a multiplex qPCR-based approach for the diagnosis of Dirofilaria immitis, D. repens and Acanthocheilonema reconditum. Parasit. Vectors 2020, 13, 319. [Google Scholar] [CrossRef] [PubMed]
- Laidoudi, Y.; Otranto, D.; Stolowy, N.; Amrane, S.; SanthakumariManoj, R.R.; Polette, L.; Watier-Grillot, S.; Mediannikov, O.; Davoust, B.; L’Ollivier, C. Human and Animal Dirofilariasis in Southeast of France. Microorganisms 2021, 9, 1544. [Google Scholar] [CrossRef] [PubMed]
- Pupić-Bakrač, A.; Pupić-Bakrač, J.; Beck, A.; Jurković, D.; Polkinghorne, A.; Beck, R. Dirofilaria repens microfilaremia in humans: Case description and literature review. One Health 2021, 13, 100306. [Google Scholar] [CrossRef] [PubMed]
- Al-Qaoud, K.M.; Pearlman, E.; Hartung, T.; Klukowski, J.; Fleischer, B.; Hoerauf, A. A new mechanism for IL-5-dependent helminth control: Neutrophil accumulation and neutrophilmediated worm encapsulation in murine filariasis are abolished in the absence of IL-5. Int. Immunol. 2000, 12, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.E.; Maizels, R.M. Diversity and dialogue in immunity to helminths. Nat. Rev. Immunol. 2011, 11, 375–388. [Google Scholar] [CrossRef]
- Volkmann, L.; Saeftel, M.; Bain, O.; Fischer, K.; Fleischer, B.; Hoerauf, A. Interleukin-4 is essential for the control of microfilariae in murine infection with the filaria Litomosoides sigmodontis. Infect. Immun. 2001, 69, 2950–2956. [Google Scholar] [CrossRef]
- Volkmann, L.; Bain, O.; Saeftel, M.; Specht, S.; Fischer, K.; Brombacher, F.; Matthaei, K.I.; Hoerauf, A. Murine filariasis: Interleukin 4 and interleukin 5 lead to containment of different worm developmental stages. Med. Microbial. Immunol. 2003, 192, 23–31. [Google Scholar] [CrossRef]
- Saeftel, M.; Volkmann, L.; Korten, S.; Brattig, N.; Al-Qaoud, K.; Fleischer, B.; Hoerauf, A. Lack of interferon-gamma confers impaired neutrophil granulocyte function and imparts prolonged 111 survival of adult filarial worms in murine filariasis. Microbes Infect. 2001, 3, 203–213. [Google Scholar] [CrossRef]
- Maizels, R.M.; Balic, A.; Gomez-Escobar, N.; Nair, M.; Taylor, M.D.; Allen, J.E. Helminth parasites-masters of regulation. Immunol. Rev. 2004, 201, 89–116. [Google Scholar] [CrossRef]
- Taylor, M.D.; LeGoff, L.; Harris, A.; Malone, E.; Allen, J.E.; Maizels, R.M. Removal of regulatory T cell activity reverses hyporesponsiveness and leads to filarial parasite clearance in vivo. J. Immunol. 2005, 174, 4924–4933. [Google Scholar] [CrossRef] [PubMed]
- Kwarteng, A.S.T.; Ahuno, S.T.; Akoto, F.O. Killing filarial nematode parasites: Role of treatment options and host immune response. Infect. Dis. Poverty 2016, 5, 86. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.; Sun, J.; Kane, C.; Marshall, F.; Krawczyk, C.; Pearce, E.J. Review series on helminthes, immune modulation and the hygiene hypothesis: Mechanisms underlying helminthes modulation of dendritic cell function. J. Immunol. 2009, 126, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Subash, B.; Nutman, T. Proinflammatory cytokines dominate the early immune response to filarial parasites. J. Immunol. 2003, 171, 6723–6732. [Google Scholar] [CrossRef] [Green Version]
- Seidl, A.; Panzer, M.; Voehringer, D. Protective immunity against the gastrointestinal nematode Nippostrongylus brasiliensis requires a broad T-cell receptor repertoire. J. Immunol. 2011, 134, 214–223. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamada, K.D.; Tomii, K.; Katoh, K. Parallelization of MAFFT for large-scale multiple sequence alignments. Bioinformatics 2018, 34, 2490–2492. [Google Scholar] [CrossRef]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, 293–296. [Google Scholar] [CrossRef]
- Spiegel, M.R. Statistics; McGraw-Hill Natl. Book Co.: Singapore, 1981. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Hosts (no.) | No. of Filaroid-Positive Animals (%) | Species Name (Acc. No., Size bps) | 100% Identical Sequences from GenBank |
|---|---|---|---|
| Horses (320) | 1 (0.3) | Mansonella(T.) sp. (MN728216) | Mansonella sp. (MT786947-49) |
| Total | 1 (0.3) | ||
| Donkeys (109) | 9 (8.3) | Mansonella(T.) sp. (MN728182, MN728181 *) | Mansonella sp. (MT786947-49) |
| 5 (4.6) | Setaria digitata (MN728184, MN728183 **) | S. digitata (DQ094175) | |
| 1 (0.9) | Dirofilaria repens (MN728180, MN728180 *) | D. repens (MN728215) | |
| Total | 15 (13.8) |
| INF-gamma (pg/mL) | IL-5 (pg/mL) | Th1/Th2 (INF-gamma/IL-5) | IL-10 (pg/mL) | |
|---|---|---|---|---|
| Control | 23.634 ± 1.66 | 6.160 ± 0.34 | 3.84 | 8.726 ± 2.11 |
| Mansonella(T) sp. | 21.992 ± 3.18 | 11.054 ± 2.61 a | 1.99 | 17.868 ± 1.02 bB |
| S. digitata | 28.006 ± 1.09 aA | 16.362 ± 1.35 bA | 1.71 | 9.254 ± 2.99 |
| Provinces | Lat./Long. | Hosts | Locations | Total |
|---|---|---|---|---|
| Cairo | 30°03′45.47″ N, 31°14′58.81″ E | Horses | Police Academy (El-Abbasia) | 94 |
| Police Academy (El-Tagamoa) | 70 | |||
| Police Academy (El-Pasateen) | 147 | |||
| Beni-Suef | 29°03′60.00″ N, 31°04′60.00″ E | Horses | households | 9 |
| Donkeys | households | 22 | ||
| Al-Faiyum | 29°18′35.82″ N, 30°50′30.48″ E | Donkeys | households | 87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abo-Aziza, F.A.M.; Hendawy, S.H.M.; Abdullah, H.H.A.M.; El Namaky, A.; Laidoudi, Y.; Mediannikov, O. Emergent and Neglected Equine Filariosis in Egypt: Species Diversity and Host Immune Response. Pathogens 2022, 11, 979. https://doi.org/10.3390/pathogens11090979
Abo-Aziza FAM, Hendawy SHM, Abdullah HHAM, El Namaky A, Laidoudi Y, Mediannikov O. Emergent and Neglected Equine Filariosis in Egypt: Species Diversity and Host Immune Response. Pathogens. 2022; 11(9):979. https://doi.org/10.3390/pathogens11090979
Chicago/Turabian StyleAbo-Aziza, Faten A. M., Seham H. M. Hendawy, Hend H. A. M. Abdullah, Amira El Namaky, Younes Laidoudi, and Oleg Mediannikov. 2022. "Emergent and Neglected Equine Filariosis in Egypt: Species Diversity and Host Immune Response" Pathogens 11, no. 9: 979. https://doi.org/10.3390/pathogens11090979