
Effects of Ceragenins and Antimicrobial Peptides on the A549 Cell Line and an In Vitro Co-Culture Model of A549 Cells and Pseudomonas aeruginosa

Abstract
:1. Introduction
2. Results
2.1. The MICs and MBCs
2.2. MTT Results
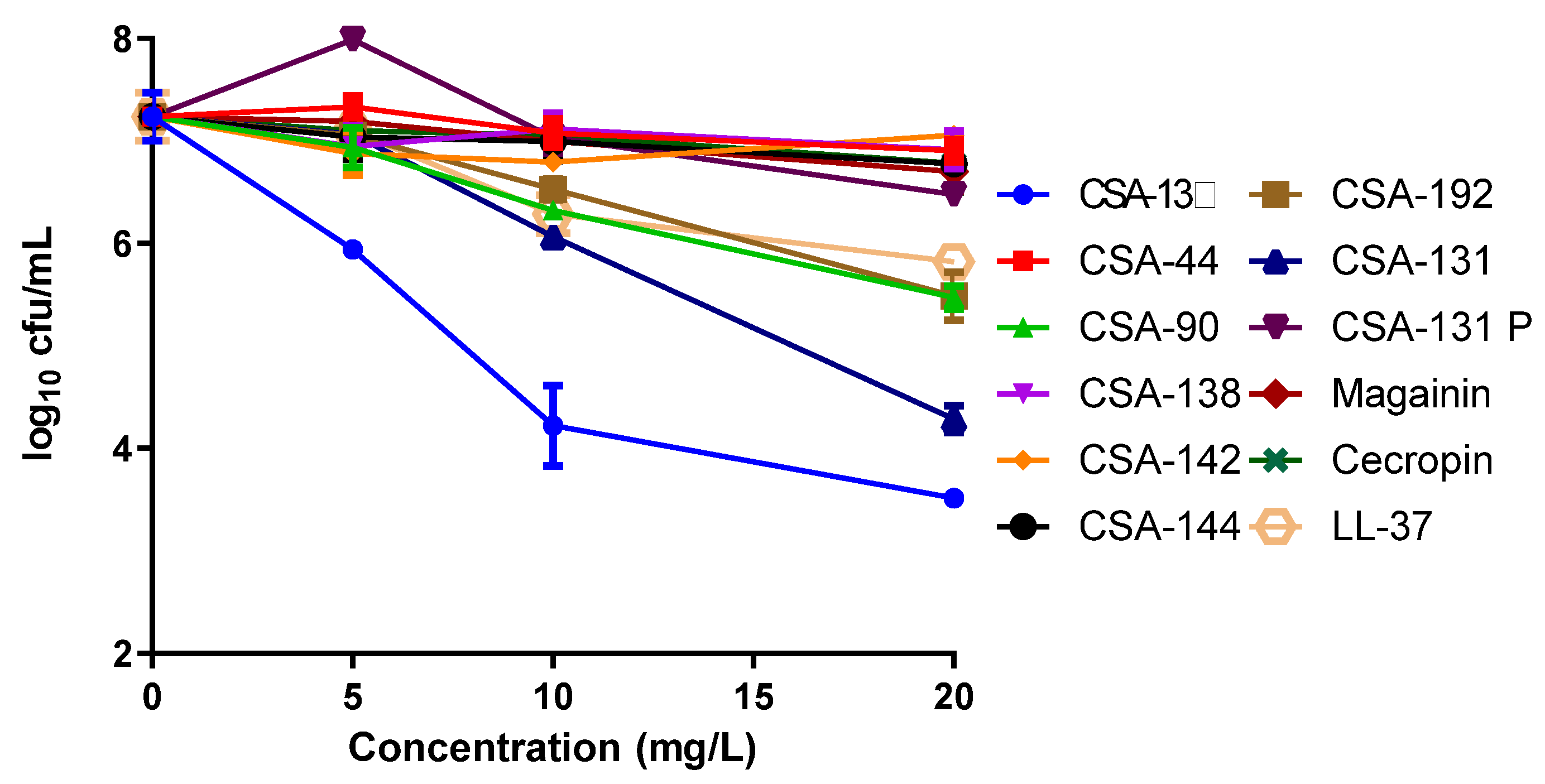
2.3. Intracellular Activity Results
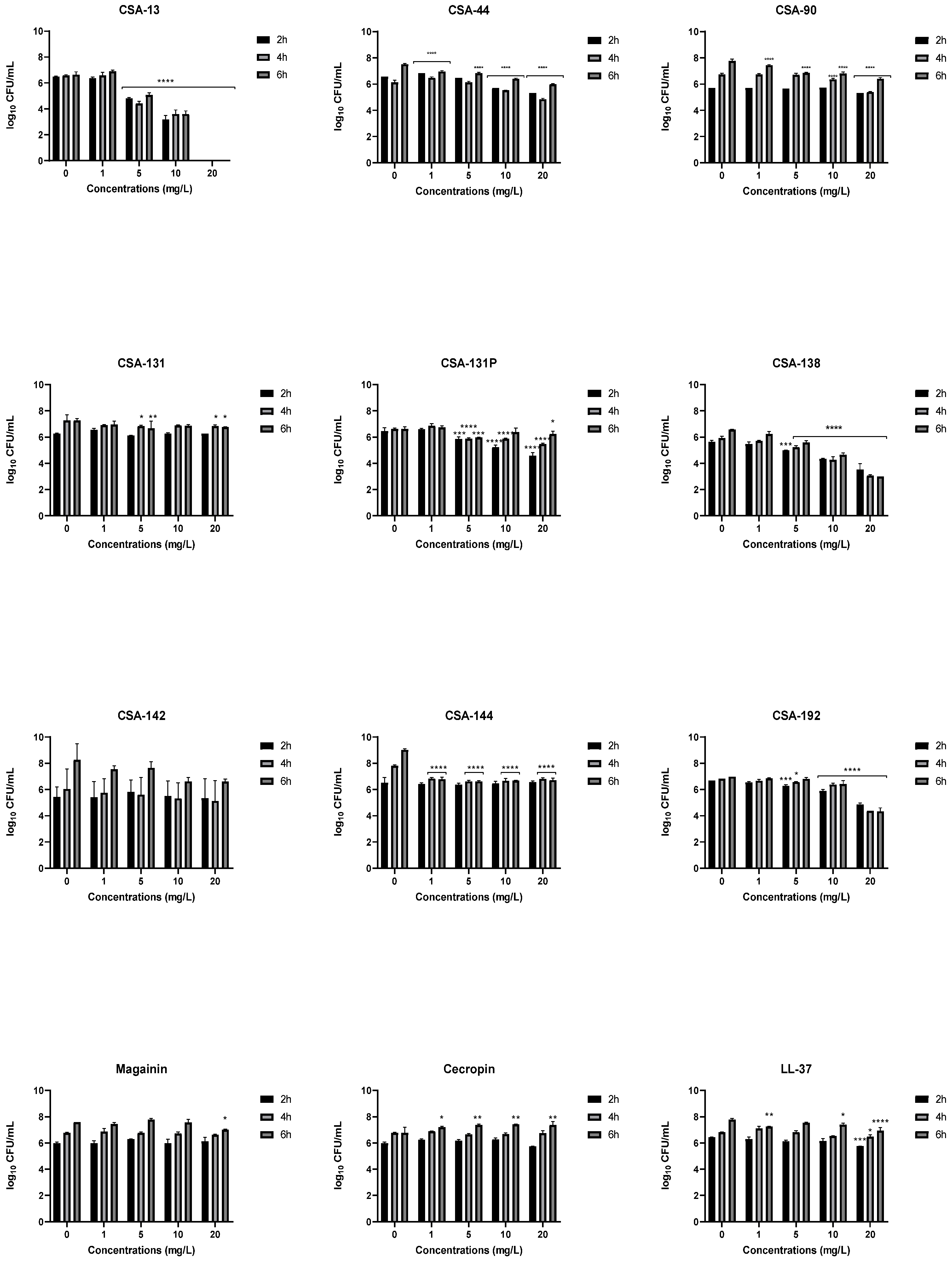
2.4. Inhibition of Bacterial Adhesion
2.5. LDH Cytotoxity Results
3. Discussion
4. Materials and Methods
4.1. Antimicrobial Agents and Bacterial Strain
4.2. Determination of MICs and MBCs
4.3. Cell Culture and Cytotoxixity
4.4. The Effects of Ceragenins and AMPs against Intracellular P. aeruginosa
4.5. The Effects of Ceragenins and AMPs on P. aeruginosa Adhesion on A549 Cells
4.6. Effect of Co-Culture on Cytotoxicity
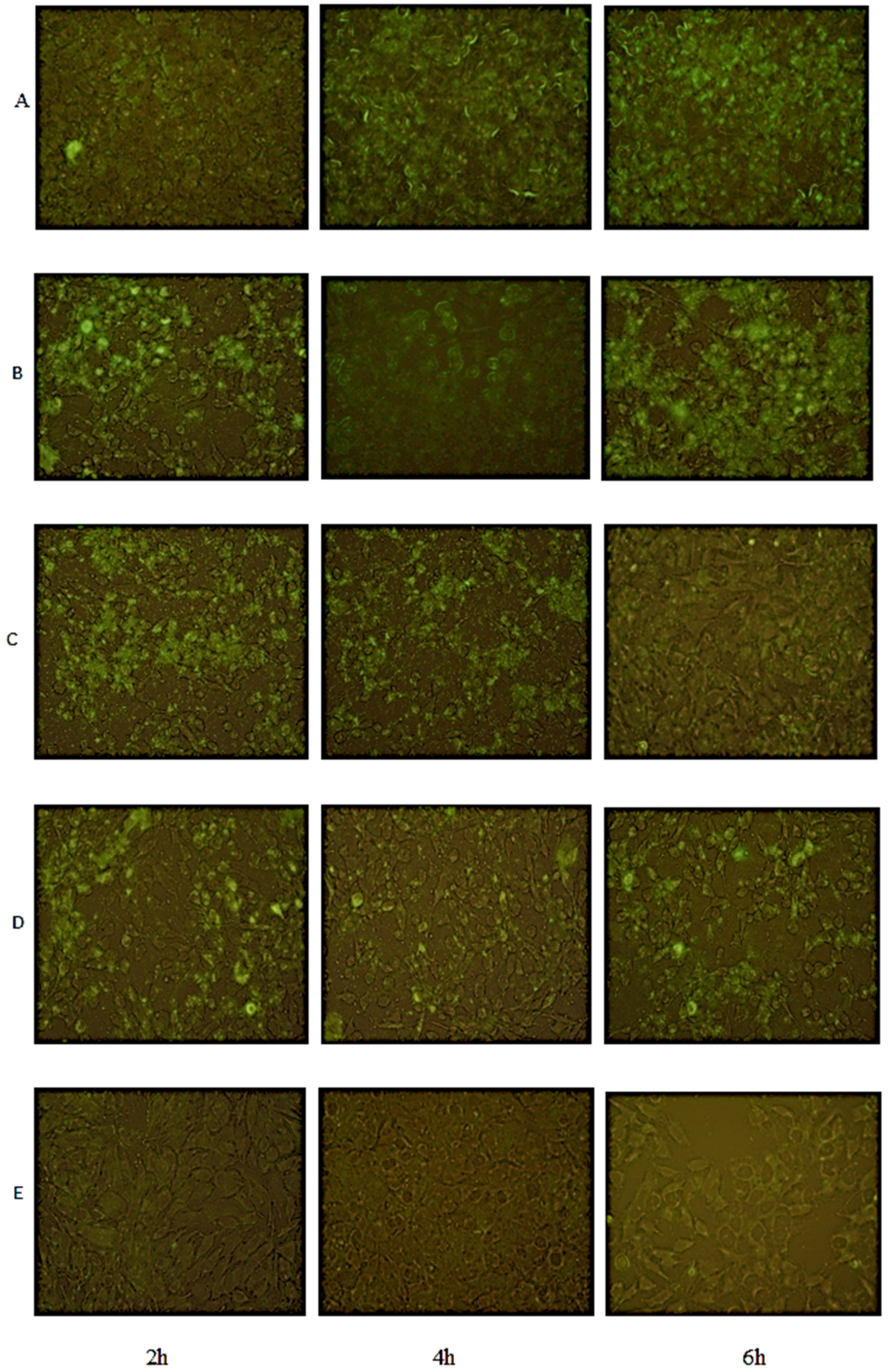
4.7. Fluorescence Microscopy
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Castañeda-Montes, F.J.; Avitia, M.; Sepúlveda-Robles, O.; Cruz-Sánchez, V.; Kameyama, L.; Guarneros, G.; Escalante, A.E. Population structure of Pseudomonas aeruginosa through a MLST approach and antibiotic resistance profiling of a Mexican clinical collection. Infect. Genet. Evol. 2018, 65, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Déraspe, M.; Iqbal, N.; Krajden, S.; Chapman, W.; Dewar, K.; Roy, P.H. Complete Genome of a Panresistant Pseudomonas aeruginosa Strain, Isolated from a Patient with Respiratory Failure in a Canadian Community Hospital. Genome Announc. 2017, 5, e00458-17. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, A.C.; Waters, V.J. Microbiology of Cystic Fibrosis Airway Disease. Semin. Respir. Crit. Care Med. 2019, 40, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Moreau-Marquis, S.; Stanton, B.A.; O’Toole, G.A. Pseudomonas aeruginosa biofilm formation in the cystic fibrosis airway. Pulm. Pharmacol. Ther. 2008, 21, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Chi, E.; Mehl, T.; Nunn, D.; Lory, S. Interaction of Pseudomonas aeruginosa with A549 pneumocyte cells. Infect. Immun. 1991, 59, 822–828. [Google Scholar] [CrossRef]
- Noore, J.; Noore, A.; Li, B. Cationic antimicrobial peptide LL-37 is effective against both extra- and intracellular Staphylococcus aureus. Antimicrob. Agents Chemother. 2013, 57, 1283–1290. [Google Scholar] [CrossRef]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The value of antimicrobial peptides in the age of resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef]
- Mercer, D.K.; Torres, M.D.T.; Duay, S.S.; Lovie, E.; Simpson, L.; von Köckritz-Blickwede, M.; de la Fuente-Nunez, C.; O’Neil, D.A.; Angeles-Boza, A.M. Antimicrobial Susceptibility Testing of Antimicrobial Peptides to Better Predict Efficacy. Front. Cell. Infect. Microbiol. 2020, 10, 326. [Google Scholar] [CrossRef]
- Hashemi, M.M.; Holden, B.S.; Savage, P.B. Ceragenins as non-peptide mimics of endogenous antimicrobial peptides. In Fighting Antimicrobial Resistance; Budimir, A., Ed.; IAPC Publishing: Zagreb, Croatia, 2018; pp. 139–169. [Google Scholar]
- Olekson, M.A.; You, T.; Savage, P.B.; Leung, K.P. Antimicrobial ceragenins inhibit biofilms and affect mammalian cell viability and migration in vitro. FEBS Open Bio. 2017, 7, 953–967. [Google Scholar] [CrossRef]
- Savage, P.B. Hydrophobic Ceragenin Compounds and Devices Incorporating Same. U.S. Patent 8,975,310, 10 March 2015. [Google Scholar]
- Shriky, B.; Kelly, A.; Isreb, M.; Babenko, M.; Mahmoudi, N.; Rogers, S.; Shebanova, O.; Snow, T.; Gough, T. Pluronic F127 thermosensitive injectable smart hydrogels for controlled drug delivery system development. J. Colloid Interface Sci. 2020, 565, 119–130. [Google Scholar] [CrossRef]
- Dao, A.; Mills, R.J.; Kamble, S.; Savage, P.B.; Little, D.G.; Schindeler, A. The application of ceragenins to orthopedic surgery and medicine. J. Orthop. Res. 2020, 38, 1883–1894. [Google Scholar] [CrossRef] [PubMed]
- Dosler, S.; Karaaslan, E. Inhibition and destruction of Pseudomonas aeruginosa biofilms by antibiotics and antimicrobial peptides. Peptides 2014, 62, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Bucki, R.; Niemirowicz, K.; Wnorowska, U.; Byfield, F.J.; Piktel, E.; Wątek, M.; Janmey, P.A.; Savage, P.B. Bactericidal Activity of Ceragenin CSA-13 in Cell Culture and in an Animal Model of Peritoneal Infection. Antimicrob. Agents Chemother. 2015, 59, 6274–6282. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt Guzel, C.; Oyardi, O.; Savage, P.B. Comparative in vitro antimicrobial activities of CSA-142 and CSA-192, second-generation ceragenins, with CSA-13 against various microorganisms. J. Chemother. 2018, 30, 332–337. [Google Scholar] [CrossRef]
- Oyardi, Ö.; Savage, P.B.; Erturan, Z.; Bozkurt-Guzel, C. In vitro assessment of CSA-131 and CSA-131 poloxamer form for the treatment of Stenotrophomonas maltophilia infections in cystic fibrosis. J. Antimicrob. Chemother. 2021, 76, 443–450. [Google Scholar] [CrossRef]
- Wnorowska, U.; Fiedoruk, K.; Piktel, E.; Prasad, S.V.; Sulik, M.; Janion, M.; Daniluk, T.; Savage, P.B.; Bucki, R. Nanoantibiotics containing membrane-active human cathelicidin LL-37 or synthetic ceragenins attached to the surface of magnetic nanoparticles as novel and innovative therapeutic tools: Current status and potential future applications. J. Nanobiotechnol. 2020, 18, 1–18. [Google Scholar] [CrossRef]
- Hashemi, M.M.; Holden, B.S.; Taylor, M.F.; Wilson, J.; Coburn, J.; Hilton, B.; Nance, T.; Gubler, S.; Genberg, C.; Deng, S.; et al. Antibacterial and Antifungal Activities of Poloxamer Micelles Containing Ceragenin CSA-131 on Ciliated Tissues. Molecules 2018, 23, 596. [Google Scholar] [CrossRef]
- Nagant, C.; Savage, P.B.; Dehaye, J.P. Effect of pluronic acid F-127 on the toxicity towards eukaryotic cells of CSA-13, a cationic steroid analogue of antimicrobial peptides. J. Appl. Microbiol. 2012, 112, 1173–1183. [Google Scholar] [CrossRef]
- Garcia-Medina, R.; Dunne, W.M.; Singh, P.K.; Brody, S.L. Pseudomonas aeruginosa acquires biofilm-like properties within airway epithelial cells. Infect. Immun. 2005, 73, 8298–8305. [Google Scholar] [CrossRef]
- Howell, M.D.; Streib, J.E.; Kim, B.E.; Lesley, L.J.; Dunlap, A.P.; Geng, D.; Feng, Y.; Savage, P.B.; Leung, D.Y. Ceragenins: A class of antiviral compounds to treat orthopox infections. J. Investig. Dermatol. 2009, 129, 2668–2675. [Google Scholar] [CrossRef] [Green Version]
- Wnorowska, U.; Piktel, E.; Durnaś, B.; Fiedoruk, K.; Savage, P.B.; Bucki, R. Use of ceragenins as a potential treatment for urinary tract infections. BMC Infect. Dis. 2019, 19, 369. [Google Scholar] [CrossRef] [PubMed]
- Chennupati, S.K.; Chiu, A.G.; Tamashiro, E.; Banks, C.A.; Cohen, M.B.; Bleier, B.S.; Kofonow, J.M.; Tam, E.; Cohen, N.A. Effects of an LL-37-derived antimicrobial peptide in an animal model of biofilm Pseudomonas sinusitis. Am. J. Rhinol. Allergy 2009, 23, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 7th ed.; M07; CLSI: Wayne, PA, USA, 2006. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 31st ed.; CLSI Supplement M100; CLSI: Wayne, PA, USA, 2021. [Google Scholar]
- Anderson, G.G.; Kenney, T.F.; Macleod, D.L.; Henig, N.R.; O’Toole, G.A. Eradication of Pseudomonas aeruginosa biofilms on cultured airway cells by a fosfomycin/tobramycin antibiotic combination. Pathog. Dis. 2013, 67, 39–45. [Google Scholar] [CrossRef]
- Anderson, G.G.; Moreau-Marquis, S.; Stanton, B.A.; O’Toole, G.A. In vitro analysis of tobramycin-treated Pseudomonas aeruginosa biofilms on cystic fibrosis-derived airway epithelial cells. Infect. Immun. 2008, 76, 1423–1433. [Google Scholar] [CrossRef] [PubMed]
- Moreau-Marquis, S.; Redelman, C.V.; Stanton, B.A.; Anderson, G.G. Co-Culture models of Pseudomonas aeruginosa biofilms grown on live human airway cells. J. Vis. Exp. 2010, 44, e2186. [Google Scholar] [CrossRef]
- Price, K.E.; Naimie, A.A.; Griffin, E.F.; Bay, C.; O’Toole, G.A. Tobramycin-Treated Pseudomonas aeruginosa PA14 Enhances Streptococcus constellatus 7155 Biofilm Formation in a Cystic Fibrosis Model System. J. Bacteriol. 2016, 198, 237–247. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. aeruginosa-GFP | ||
|---|---|---|
| Antimicrobial Agents | MIC | MBC |
| CSA-13 | 16 | 32 |
| CSA-44 | 8 | 16 |
| CSA-90 | 32 | 64 |
| CSA-131 | 4 | 4 |
| CSA-131P | 8 | 8 |
| CSA-138 | 16 | 16 |
| CSA-142 | 8 | 8 |
| CSA-144 | 32 | 64 |
| CSA-192 | 32 | 64 |
| Magainin | >128 | >128 |
| Cecropin | >128 | >128 |
| LL-37 | >128 | >128 |
| Ceragenins | IC50 Values (mg/L) |
|---|---|
| CSA-13 | 20.68 ± 4.34 |
| CSA-44 | 26.03 ± 0.95 |
| CSA-90 | 11.62 ± 4.05 |
| CSA-131 | 11.37 ± 3.88 |
| CSA-131P | 40.05 ± 2.65 |
| CSA-138 | 11.78 ± 2.38 |
| CSA-142 | 36.15 ± 1.25 |
| CSA-144 | 26.03 ± 2.08 |
| CSA-192 | 27.65 ± 5.02 |
| Magainin | >200 |
| Cecropin | >200 |
| LL-37 | 40.76 ± 5.35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oyardi, O.; Savage, P.B.; Guzel, C.B. Effects of Ceragenins and Antimicrobial Peptides on the A549 Cell Line and an In Vitro Co-Culture Model of A549 Cells and Pseudomonas aeruginosa. Pathogens 2022, 11, 1044. https://doi.org/10.3390/pathogens11091044
Oyardi O, Savage PB, Guzel CB. Effects of Ceragenins and Antimicrobial Peptides on the A549 Cell Line and an In Vitro Co-Culture Model of A549 Cells and Pseudomonas aeruginosa. Pathogens. 2022; 11(9):1044. https://doi.org/10.3390/pathogens11091044
Chicago/Turabian StyleOyardi, Ozlem, Paul B. Savage, and Cagla Bozkurt Guzel. 2022. "Effects of Ceragenins and Antimicrobial Peptides on the A549 Cell Line and an In Vitro Co-Culture Model of A549 Cells and Pseudomonas aeruginosa" Pathogens 11, no. 9: 1044. https://doi.org/10.3390/pathogens11091044